 ,1,2,3,*
,1,2,3,*Screening of germplasm tolerant to low phosphorus of seedling stage and response of root protective enzymes to low phosphorus in foxtail millet
YUAN Yi-Chuan1,*, CHEN Xiao-Yu1,*, LI Ming-Ming1, LI Ping1, JIA Ya-Tao1, HAN Yuan-Huai1,2,3, XING Guo-Fang,1,2,3,*通讯作者:
收稿日期:2018-07-25接受日期:2018-12-24网络出版日期:2019-01-04
| 基金资助: |
Received:2018-07-25Accepted:2018-12-24Online:2019-01-04
| Fund supported: |
作者简介 About authors

摘要
关键词:
Abstract
Keywords:
PDF (1042KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
苑乂川, 陈小雨, 李明明, 李萍, 贾亚涛, 韩渊怀, 邢国芳. 谷子苗期耐低磷种质筛选及其根系保护酶系统对低磷胁迫的响应[J]. 作物学报, 2019, 45(4): 601-612. doi:10.3724/SP.J.1006.2019.82029
YUAN Yi-Chuan, CHEN Xiao-Yu, LI Ming-Ming, LI Ping, JIA Ya-Tao, HAN Yuan-Huai, XING Guo-Fang.
磷(P)是作物必需的营养元素之一[1], 对植物生长发育起着至关重要的作用。但土壤中能被作物吸收利用的有效磷仅占土壤全磷的1%[2], 土壤有效磷不足已成为制约作物产量和品质提高的主要因素之一。另外, 磷是不可再生资源, 50~400年内将会被耗尽[3,4]。为了应对未来的磷缺陷对农业生产带来的危机, 了解作物磷胁迫响应机制, 培育耐低磷品种显得尤为重要[5]。
了解作物磷胁迫响应机制, 培育耐低磷品种, 筛选耐低磷种植资源, 鉴定耐低磷材料是关键。目前关于耐低磷种质资源的筛选与评价, 在水稻、玉米等作物中已经开展了大量的试验。郭再华等[6]对来自不同国家的多个水稻品种开展苗期土培筛选, 设置多个磷浓度处理, 确定了水稻苗期耐低磷指标筛选的最适营养液配比和鉴定指标。湛静等[7]在缺磷条件下对玉米苗期进行了性状和生理机制的研究, 获得了一套比较完善的玉米耐低磷指标筛选体系。刘渊等[8]以酸性磷酸酶的耐低磷系数作为评判基础, 分析了不同磷水平下大豆株高、干重、磷含量等指标, 发现不同基因型品种耐低磷差异极大, 并筛选出耐低磷种质。深入研究同一作物不同基因型品种间所表现的磷吸收和利用的生理特点差异, 它的意义不仅在于筛选磷高效品种, 还致力于形成完善的耐低磷筛选体系。
谷子(Setaria italica L.)是原产我国北方的小杂粮, 耐旱, 耐贫瘠, 具有养分高效利用的特质和潜力[9,10], 是中国北方地区最主要的种植作物之一[11]。谷子对磷缺乏有着相应的适应机制, 不同基因型谷子在磷吸收方面存在很大差异[12,13,14]。缺磷会对谷子的生长发育造成一定影响, 杨艳君等[15]发现磷缺乏导致收获期谷穗重量降低, Ceasar等[16]在处理后前2周, 发现300 μmol L-1与10 μmol L-1供磷量对谷子生物量的影响无差异, 5周后, 10 μmol L-1供磷水平下的谷子出现叶片变紫等缺磷症状。据报道, 谷子中存在12个磷转运载体PHR家族成员, 并且不同成员之间存在组织特异性表达[16]。然而, 目前关于谷子耐低磷基因型品种筛选的相关研究并未深入。
植物体内的保护性酶如超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT), 是抵抗营养胁迫引起氧化应激的关键[17]。逆境胁迫不仅会影响植物的正常生理功能, 还会导致植物的新陈代谢失去平衡[18]。植物的抗逆能力与植物保护性酶系统的活性变化之间存在密切的关联性。本研究从实验室1500多份材料中挑选了160份核心种质资源, 综合鉴定与评价了它们苗期的株高、根长、地上部鲜重、地下部鲜重、叶长、叶宽、茎粗、地上部磷含量、地下部磷含量、地上部干重和地下部干重, 筛选出耐低磷特性的指标和苗期具有耐低磷特性的谷子种质, 并进一步研究苗期根系保护酶系统在低磷胁迫下的响应, 为在遗传育种研究中利用这些优异种质资源提供理论依据。
1 材料与方法
1.1 供试材料
160份核心谷子种质资源, 由中国农业科学院作物科学研究所刁现民研究员提供。1.2 试验设计
在山西农业大学农学院温室内进行随机排列的盆栽试验, 以石英沙培养和营养液浇灌, 筛选耐低磷品种。挑选整齐饱满的种子, 播于育苗盆(长32.5 cm, 宽25.0 cm, 深7.0 cm), 盆内放置总体积4/5的石英沙, 播种行距4.5 cm, 株距3.5 cm, 双粒点播。待幼苗生长至一叶一心期时, 向盆内加Hoagland营养液, 待幼苗至三叶期时间苗, 培育至30 d。另以水培试验用于谷子根系酶活性鉴定, 先在石英沙中盆栽培育至一心一叶期, 然后移栽到Hoagland营养液中, 每3天更换1次营养液, 培育至30 d。试验条件均为温度25℃, 光照175 μmol m-2 s-1, 光周期16 h光照/8 h黑暗。设低磷水平(LP, 0.005 mmol L-1)和正常磷水平(NP, 0.25 mmol L-1) 2个处理。
1.3 指标测定
测量生长30 d谷子幼苗的株高、叶长、叶宽、茎粗和根长; 用 H2SO4-H2O2消解和钒钼黄比色法测定地上和地下磷含量[19], 氮蓝四唑(NBT)光化还原法测定根系SOD活性, 愈创木酚法测定根系POD活性, 紫外吸收法测定根系CAT活性[20]。1.4 数据处理
采用Origin 8和Microsoft Excel 2003作图表, 利用SPSS 19.0进行相关性分析、主成分分析、方差分析, 采用LSD进行显著性检验, 利用DPS 7.05和R语言进行聚类分析, 采用隶属函数法对160份核心谷子种质进行耐低磷综合评价, 参考相关文献[21]计算指标。式中, μ (Xj)表示第j个综合指标的隶属函数值, Xj表示第 j 个综合指标值; Xmax表示第j个综合指标的最大值, Xmin表示第j个综合指标的最小值。
式中, Wj表示第j个综合指标在所有综合指标中的重要程度及权重; Pj为各品种第j个综合指标的贡献率。
式中, D表示在低磷胁迫条件下各品种耐低磷能力的综合评价值。
2 结果与分析
2.1 不同品种单项指标耐低磷系数的分析
160份不同基因型的谷子在NP与LP环境下各指标均存在不同程度的变化。为避免作物的生物学固有差异, 采用了能准确反映品种耐低磷能力的耐低磷系数(相对值)作为判断谷子耐低磷特性的依据[22]。株高、地上部鲜重、叶长、地上部磷含量、地下部磷含量和地上部干重耐低磷系数均小于1, 叶宽有9个品种耐低磷系数大于1, 茎粗仅有1个品种耐低磷系数大于1, 综合说明低磷胁迫抑制地上部各性状生长, 低磷下谷子磷含量低于正常环境下磷含量。大部分品种根长、地下部鲜重和地下部干重耐低磷系数大于1, 说明低磷影响植物的根系发育。11个指标的相对值变异系数均在10%以上, 可以看出, 不同基因型谷子的耐低磷特性存在较大差异, 160份种质在不同磷水平下存在广泛的遗传变异。鉴于上述各指标在不同磷环境中变异情况均较大, 说明这些指标都可能是鉴定谷子耐低磷能力较好的筛选指标(表1)。Table 1
表1
表1谷子部分性状耐低磷系数(LP/NP)的变化范围及变异系数
Table 1
| 性状 Trait | 变化范围 Variation range (%) | 平均值 Average (%) | 变异系数 Coefficient of variance (%) |
|---|---|---|---|
| 株高Plant height | 56.15-93.05 | 71.16 | 11.00 |
| 根长Root length | 51.92-140.88 | 105.48 | 15.61 |
| 地下鲜重Underground fresh weight | 92.47-151.40 | 149.02 | 18.42 |
| 地上鲜重Aboveground fresh weight | 35.04-90.63 | 67.13 | 14.19 |
| 叶长Leaf length | 37.90-59.86 | 46.69 | 26.69 |
| 叶宽Leaf width | 50.00-122.73 | 81.84 | 12.93 |
| 茎粗Diameter of main stem | 44.96-106.20 | 74.26 | 11.39 |
| 地上磷含量Aboveground phosphorus content | 36.00-95.00 | 72.09 | 15.80 |
| 地下磷含量Subsurface phosphorus content | 24.00-89.00 | 58.85 | 20.69 |
| 地上干重Aboveground dry weight | 36.00-91.00 | 66.89 | 14.98 |
| 地下干重Underground dry weight | 41.00-124.00 | 75.74 | 17.34 |
新窗口打开|下载CSV
2.2 指标间耐低磷指数的相关性分析
地上部磷含量的耐低磷系数与株高、地上部鲜重、叶长、叶宽、地下部磷含量、地上部干重、地下部干重等指标的耐低磷系数呈极显著正相关, 与地下部鲜重耐低磷系数呈极显著负相关; 株高的耐低磷系数与地上部鲜重、叶长、叶宽、茎粗、地上部磷含量、地下部干重等指标的耐低磷系数呈极显著正相关, 与地上部干重耐低磷系数呈显著正相关, 与地下部鲜重耐低磷系数呈极显著负相关; 根长的耐低磷系数与地下部鲜重耐低磷系数极显著正相关, 与叶长耐低磷系数呈极显著负相关; 株高、地上部鲜重、叶长和茎粗4个指标的耐低磷系数均存在两两之间极显著正相关; 地上部磷含量、地上部干重和地下部干重3个指标之间的耐低磷系数均为极显著正相关(表2)。可见, 部分指标间存在一定相关关系, 不同指标间可能存在信息重叠, 单靠单项指标难以对表现复杂的耐低磷性综合评价, 综合各项指标能更有效地筛选低磷耐性种质资源。Table 2
表2
表2谷子各性状耐低磷系数(LP/NP)的相关性分析
Table 2
| 性状 Trait | 株高 PH | 根长 RL | 地上部 鲜重 AFW | 地下部 鲜重 UFW | 叶长 LL | 叶宽 LW | 茎粗 DMS | 地上部 磷含量 APC | 地下部 磷含量 SPC | 地上部 干重 ADW | 地下部 干重 UDW |
|---|---|---|---|---|---|---|---|---|---|---|---|
| PH | 1 | ||||||||||
| RL | 0.036 | 1 | |||||||||
| AFW | 0.442** | 0.172* | 1 | ||||||||
| UFW | -0.531** | 0.821** | -0.103 | 1 | |||||||
| LL | 0.648** | -0.542** | 0.603** | -0.808** | 1 | ||||||
| LW | 0.343** | -0.200* | 0.066 | -0.361** | 0.334** | 1 | |||||
| DMS | 0.249** | -0.044 | 0.357** | -0.179* | 0.300** | 0.109 | 1 | ||||
| APC | 0.268** | -0.121 | 0.190* | -0.233** | 0.303** | 0.225** | 0.011 | 1 | |||
| SPC | 0.019 | 0.104 | 0.269** | 0.075 | 0.066 | -0.153 | -0.024 | 0.573** | 1 | ||
| ADW | 0.187* | 0.054 | 0.494** | -0.069 | 0.290** | 0.0177 | 0.128 | 0.477** | 0.557** | 1 | |
| UDW | 0.222** | -0.073 | 0.100 | -0.179* | 0.196* | 0.318** | -0.084 | 0.365** | 0.115 | 0.352** | 1 |
新窗口打开|下载CSV
2.3 主成分分析
为达到简化指标及浓缩数据的目的, 将上述单项指标转化为有效的少量指标[21], 以160份核心谷子种质各单项指标的耐低磷系数为基础进行主成分分析。从11个主要耐低磷特征指标在各个主成分上的权重系数即特征向量看, 可以发现, 第1主成分贡献率最大 , 主要特征向量包括株高、地上部鲜重、叶长、叶宽、茎粗、地上部磷含量和地上部干重7个单项指标, 特征值为4.657, 反映了地上因子, 可说明原始指标信息量的42.334%; 第2主成分载荷较高的是根长和地下部鲜重, 反映了根系因子, 可说明原始指标信息量的20.793%; 第3主成分载荷较高的是地下部磷含量, 可反映总信息量的11.206%; 第4主成分载荷较高的是地下部干重, 可反映总信息量的8.719%。前4个主成分累积贡献率达83.049%, 足以代表原指标的极大部分信息, 其余贡献率可忽略不计(表3和表4)。因此, 利用4个相互独立的综合指标可以对不同谷子的耐低磷特性进行客观的分析。Table 3
表3
表3各综合指标特征值及贡献率
Table 3
| 主成分 Principal component | 特征值 Eigenvalue | 贡献率 Contributive ratio (%) | 累积贡献率 Cumulative contributive ratio (%) |
|---|---|---|---|
| 1 | 4.657 | 42.334 | 42.334 |
| 2 | 2.287 | 20.793 | 63.127 |
| 3 | 1.233 | 11.206 | 74.333 |
| 4 | 0.959 | 8.716 | 83.049 |
新窗口打开|下载CSV
Table 4
表4
表4各因子载荷矩阵
Table 4
| 耐低磷指标 Low-P tolerant index | 主成分 Principal component | |||
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |
| 株高PH | 0.814 | -0.180 | 0.319 | 0.007 |
| 根长RL | 0.421 | 0.827 | 0.260 | -0.019 |
| 地上部鲜重AFW | 0.810 | 0.005 | 0.156 | -0.412 |
| 地下部鲜重UFW | 0.164 | 0.957 | 0.114 | 0.031 |
| 叶长LL | 0.580 | -0.744 | 0.074 | -0.233 |
| 叶宽LW | 0.647 | -0.172 | 0.336 | 0.479 |
| 茎粗DMS | 0.673 | 0.027 | 0.408 | -0.234 |
| 地上部磷含量APC | 0.748 | -0.043 | -0.432 | 0.193 |
| 地下部磷含量SPC | 0.584 | 0.236 | 0.649 | -0.202 |
| 地上部干重ADW | 0.793 | 0.106 | -0.344 | -0.113 |
| 地下部干重UDW | 0.529 | -0.044 | -0.119 | 0.599 |
新窗口打开|下载CSV
2.4 品种耐低磷能力的综合分析
2.4.1 隶属函数鉴定品种耐低磷能力 根据主成分分析结果, 利用公式(2)计算每个综合指标的隶属函数值μ(Xj)。从表5可以看出, 低磷胁迫下, 综合指标Z1中69号品种μ(X1)最小, 为0, 表明其在Z1中耐低磷能力最弱; V147号品种μ(X1)最大, 为1.0000, 表明其在Z1中耐低磷能力最强; 综合指标Z2、Z3和Z4中μ(X)最小的品种分别为V151、V19、V81号, μ(X)最大的品种分别V72、V35、V69。可见4个综合指标鉴定的结果各不相同, 因此, 为对谷子耐低磷性进行全面充实的分析, 通过公式(3), 结合各综合指标的贡献率, 得出4个综合指标的权重Wj分别为50.97%、25.04%、13.49%和10.05%, 进一步根据公式(4)计算4个综合指标的综合评价值D, 并根据D值大小对其耐低磷能力的强弱进行排序。结果见表5, 排名较前的品种D值较小, 表明耐低磷能力弱, 称之为磷敏感品种, 排名较后的品种D值较大, 说明耐低磷力强, 称之为磷耐性品种。Table 5
表5
表5各品种的综合性状指标值、权重、μ(X)及综合评价值(D)
Table 5
| 品种 Variety | Z1 | Z2 | Z3 | Z4 | μ(X1) | μ(X2) | μ(X3) | μ(X4) | 综合评价值 Comprehensive assessment value (D) | 排名 Order |
|---|---|---|---|---|---|---|---|---|---|---|
| V1 | -1.6323 | 0.3830 | 1.7162 | -0.3955 | 0.2264 | 0.6479 | 0.7765 | 0.3123 | 0.4152 | 27 |
| V2 | -1.4500 | -2.1682 | 1.3999 | -0.5085 | 0.2539 | 0.3513 | 0.7244 | 0.2908 | 0.3456 | 7 |
| V3 | -1.2102 | -0.6284 | -0.2063 | 1.0063 | 0.2901 | 0.5303 | 0.4597 | 0.5793 | 0.4035 | 20 |
| V4 | 1.6335 | 0.7376 | -2.4004 | 1.1895 | 0.7197 | 0.6891 | 0.0981 | 0.6142 | 0.6171 | 134 |
| V5 | 2.1822 | -0.2511 | -2.4419 | 0.4781 | 0.8026 | 0.5742 | 0.0913 | 0.4787 | 0.6154 | 132 |
| V6 | 0.0319 | 0.6162 | 0.2194 | -0.2165 | 0.4778 | 0.6750 | 0.5298 | 0.3464 | 0.5204 | 79 |
| V7 | -0.0242 | -1.3712 | -0.1959 | 0.1746 | 0.4693 | 0.4440 | 0.4614 | 0.4209 | 0.4568 | 49 |
| V8 | 0.1818 | 0.6668 | -0.6091 | -1.6641 | 0.5004 | 0.6809 | 0.3933 | 0.0707 | 0.4860 | 67 |
| V9 | 3.0438 | -2.0966 | 1.9004 | 0.6023 | 0.9327 | 0.3596 | 0.8069 | 0.5024 | 0.7270 | 158 |
| V10 | 0.3314 | 1.3360 | -1.2708 | 0.1252 | 0.5230 | 0.7587 | 0.2842 | 0.4115 | 0.5381 | 95 |
| V11 | -1.3979 | -0.0456 | 0.3979 | -0.1125 | 0.2618 | 0.5981 | 0.5593 | 0.3662 | 0.3971 | 17 |
| V12 | -1.1021 | 1.0375 | 0.4391 | 0.0901 | 0.3065 | 0.7240 | 0.5660 | 0.4048 | 0.4564 | 47 |
| V13 | 0.7983 | 1.0699 | -1.3880 | -0.9506 | 0.5935 | 0.7278 | 0.2649 | 0.2066 | 0.5422 | 97 |
| V14 | -1.6737 | 2.0691 | -0.2322 | 1.2051 | 0.2201 | 0.8439 | 0.4554 | 0.6172 | 0.4498 | 42 |
| V15 | 1.0917 | -0.5273 | -1.6633 | 0.7557 | 0.6378 | 0.5421 | 0.2196 | 0.5316 | 0.5463 | 99 |
| V16 | -0.5638 | -0.1478 | 0.0901 | -1.2853 | 0.3878 | 0.5862 | 0.5085 | 0.1428 | 0.4280 | 30 |
| V17 | -1.8131 | -0.7841 | -0.2550 | 1.8789 | 0.1991 | 0.5122 | 0.4517 | 0.7455 | 0.3689 | 12 |
| V18 | -2.9829 | -0.8147 | 0.6199 | -0.8614 | 0.0224 | 0.5087 | 0.5958 | 0.2236 | 0.2426 | 1 |
| V19 | 0.0209 | 2.1536 | -2.9956 | 1.1885 | 0.4761 | 0.8537 | 0.0000 | 0.6140 | 0.5209 | 81 |
| V20 | -1.7178 | 0.1697 | -1.5586 | 0.5914 | 0.2135 | 0.6231 | 0.2368 | 0.5003 | 0.3493 | 9 |
| V21 | 0.2587 | 0.1713 | 0.1004 | 1.0974 | 0.5120 | 0.6233 | 0.5102 | 0.5967 | 0.5485 | 102 |
| V22 | -0.3407 | -1.1073 | 1.5481 | -0.6580 | 0.4215 | 0.4747 | 0.7488 | 0.2623 | 0.4622 | 52 |
| V23 | -2.7587 | 3.3932 | -0.1598 | 1.4613 | 0.0563 | 0.9979 | 0.4673 | 0.6660 | 0.4115 | 24 |
| V24 | 2.3725 | 0.2468 | -1.4864 | 0.5768 | 0.8313 | 0.6321 | 0.2487 | 0.4975 | 0.6678 | 149 |
| V25 | 2.8794 | 0.2426 | -1.2286 | 1.4558 | 0.9079 | 0.6316 | 0.2912 | 0.6649 | 0.7300 | 159 |
| V26 | 1.9496 | 0.0191 | -2.3027 | 0.9441 | 0.7674 | 0.6056 | 0.1142 | 0.5675 | 0.6178 | 135 |
| 品种 Variety | Z1 | Z2 | Z3 | Z4 | μ(X1) | μ(X2) | μ(X3) | μ(X4) | 综合评价值 Comprehensive assessment value (D) | 排名 Order |
| V27 | -0.0735 | 1.0066 | -0.2675 | 0.0285 | 0.4618 | 0.7204 | 0.4496 | 0.3931 | 0.5177 | 76 |
| V28 | -1.4796 | -2.7691 | -1.5485 | 0.4326 | 0.2495 | 0.2815 | 0.2385 | 0.4700 | 0.2791 | 2 |
| V29 | -0.3805 | -1.4994 | 0.0644 | -0.8302 | 0.4155 | 0.4291 | 0.5043 | 0.2295 | 0.4113 | 23 |
| V30 | -2.3010 | 0.5568 | 0.8604 | -0.4532 | 0.1254 | 0.6681 | 0.6355 | 0.3013 | 0.3486 | 8 |
| V31 | -0.3831 | -0.6384 | 0.3210 | -0.2612 | 0.4151 | 0.5292 | 0.5466 | 0.3379 | 0.4533 | 45 |
| V32 | -0.3526 | -0.8341 | 0.2491 | 0.3958 | 0.4197 | 0.5064 | 0.5347 | 0.4630 | 0.4615 | 51 |
| V33 | -1.2572 | 2.0635 | 0.1175 | -0.6559 | 0.2830 | 0.8433 | 0.5130 | 0.2627 | 0.4522 | 44 |
| V34 | -2.0066 | -0.0741 | 1.2416 | -0.0945 | 0.1699 | 0.5948 | 0.6983 | 0.3696 | 0.3685 | 11 |
| V35 | -2.9688 | 1.9081 | 3.0723 | 0.2469 | 0.0245 | 0.8252 | 1.0000 | 0.4347 | 0.3997 | 18 |
| V36 | -1.1128 | 2.6623 | 1.4580 | 0.2940 | 0.3049 | 0.9129 | 0.7340 | 0.4436 | 0.5296 | 89 |
| V37 | -1.0320 | 0.6961 | -0.0191 | 0.7611 | 0.3171 | 0.6843 | 0.4905 | 0.5326 | 0.4551 | 46 |
| V38 | 2.3586 | -0.0702 | -0.7951 | -0.5512 | 0.8292 | 0.5952 | 0.3626 | 0.2827 | 0.6503 | 146 |
| V39 | 1.3125 | -0.1426 | 0.7184 | 0.8312 | 0.6712 | 0.5868 | 0.6121 | 0.5459 | 0.6289 | 142 |
| V40 | 1.7558 | -1.0203 | -1.9245 | 1.5502 | 0.7382 | 0.4848 | 0.1765 | 0.6829 | 0.5931 | 123 |
| V41 | -0.4456 | -0.3004 | 0.9464 | -0.0120 | 0.4056 | 0.5685 | 0.6496 | 0.3853 | 0.4772 | 63 |
| V42 | -1.8962 | -2.1770 | 0.8829 | 0.1763 | 0.1865 | 0.3503 | 0.6392 | 0.4212 | 0.3132 | 4 |
| V43 | -0.8274 | 0.2928 | -1.8849 | -0.7725 | 0.3480 | 0.6374 | 0.1830 | 0.2405 | 0.3869 | 14 |
| V44 | -1.3986 | 0.4234 | -0.7386 | -1.0700 | 0.2617 | 0.6526 | 0.3720 | 0.1838 | 0.3663 | 10 |
| V45 | 0.0631 | -0.2475 | 0.0927 | -0.7990 | 0.4825 | 0.5746 | 0.5090 | 0.2355 | 0.4832 | 64 |
| V46 | -0.0955 | 1.4289 | -0.2399 | 0.3159 | 0.4585 | 0.7695 | 0.4541 | 0.4478 | 0.5347 | 91 |
| V47 | 1.7838 | 1.5395 | -2.2310 | 1.2522 | 0.7424 | 0.7824 | 0.1260 | 0.6261 | 0.6570 | 147 |
| V48 | 0.3831 | 1.5463 | -1.2741 | -0.8337 | 0.5308 | 0.7831 | 0.2837 | 0.2288 | 0.5290 | 88 |
| V49 | -0.5436 | 0.5557 | 1.0839 | 1.1437 | 0.3908 | 0.6680 | 0.6723 | 0.6055 | 0.5207 | 80 |
| V50 | -1.3887 | 0.8153 | 1.0506 | 0.8640 | 0.2632 | 0.6982 | 0.6668 | 0.5522 | 0.4569 | 50 |
| V51 | 2.1270 | -1.1928 | -0.1009 | -0.0165 | 0.7942 | 0.4647 | 0.4771 | 0.3845 | 0.6259 | 140 |
| V52 | 0.3242 | 0.7315 | -0.8191 | -0.0001 | 0.5219 | 0.6884 | 0.3587 | 0.3876 | 0.5275 | 87 |
| V53 | 0.2900 | -0.6707 | -1.9185 | -1.2474 | 0.5168 | 0.5254 | 0.1775 | 0.1500 | 0.4347 | 32 |
| V54 | -0.6756 | 0.2877 | -0.4629 | -0.1039 | 0.3709 | 0.6368 | 0.4174 | 0.3678 | 0.4434 | 37 |
| V55 | 3.0862 | -0.8419 | 0.2682 | 0.2587 | 0.9391 | 0.5055 | 0.5379 | 0.4369 | 0.7237 | 156 |
| V56 | -0.1720 | 1.2193 | -1.3032 | -0.0738 | 0.4470 | 0.7451 | 0.2789 | 0.3736 | 0.4913 | 68 |
| V57 | -0.6719 | 0.9067 | -1.0655 | -0.6782 | 0.3715 | 0.7088 | 0.3181 | 0.2585 | 0.4369 | 33 |
| V58 | 2.8844 | 0.6624 | -0.2868 | 1.0581 | 0.9086 | 0.6804 | 0.4464 | 0.5892 | 0.7556 | 160 |
| V59 | 1.3463 | 2.2963 | 0.1229 | 0.2876 | 0.6763 | 0.8703 | 0.5139 | 0.4424 | 0.6784 | 151 |
| V60 | -1.5799 | 2.0050 | 0.9341 | -0.0246 | 0.2343 | 0.8365 | 0.6476 | 0.3829 | 0.4565 | 48 |
| V61 | -1.2737 | 1.9840 | -0.6119 | 1.3180 | 0.2806 | 0.8340 | 0.3928 | 0.6387 | 0.4719 | 60 |
| V62 | -1.9948 | 2.2161 | 0.1507 | 0.5801 | 0.1716 | 0.8610 | 0.5185 | 0.4981 | 0.4253 | 29 |
| V63 | 0.7281 | 1.9543 | 0.0045 | 0.4504 | 0.5829 | 0.8306 | 0.4944 | 0.4734 | 0.6215 | 137 |
| V64 | -1.9503 | -0.9239 | -0.0778 | -0.5369 | 0.1784 | 0.4960 | 0.4809 | 0.2854 | 0.3099 | 3 |
| V65 | -0.4570 | 1.5584 | 1.3589 | 0.4630 | 0.4039 | 0.7846 | 0.7176 | 0.4758 | 0.5491 | 103 |
| V66 | -0.1959 | -0.8819 | -0.1243 | 0.5303 | 0.4434 | 0.5009 | 0.4732 | 0.4886 | 0.4665 | 55 |
| V67 | -1.4532 | -1.5355 | 1.5459 | 1.4533 | 0.2534 | 0.4249 | 0.7484 | 0.6644 | 0.4063 | 22 |
| V68 | -0.0935 | 1.1747 | -1.0937 | 0.9499 | 0.4588 | 0.7399 | 0.3134 | 0.5686 | 0.5211 | 82 |
| V69 | -3.1311 | 0.7949 | -1.4602 | 3.2151 | 0.0000 | 0.6958 | 0.2530 | 1.0000 | 0.3134 | 5 |
| V70 | -0.8250 | -0.4004 | -0.0584 | -0.2687 | 0.3483 | 0.5568 | 0.4841 | 0.3365 | 0.4176 | 28 |
| 品种 Variety | Z1 | Z2 | Z3 | Z4 | μ(X1) | μ(X2) | μ(X3) | μ(X4) | 综合评价值 Comprehensive assessment value (D) | 排名 Order |
| V71 | -1.4984 | 2.0480 | 1.6374 | -0.5784 | 0.2466 | 0.8415 | 0.7635 | 0.2775 | 0.4685 | 57 |
| V72 | 0.3822 | 3.4116 | 0.4863 | -0.6490 | 0.5307 | 1.0000 | 0.5738 | 0.2640 | 0.6260 | 141 |
| V73 | -1.3044 | 1.9319 | 1.5631 | -0.2074 | 0.2759 | 0.8280 | 0.7513 | 0.3481 | 0.4859 | 66 |
| V74 | 0.3536 | 0.6448 | -0.2815 | -1.0117 | 0.5264 | 0.6783 | 0.4473 | 0.1949 | 0.5189 | 77 |
| V75 | -0.4450 | 1.4046 | -0.1209 | -0.5443 | 0.4057 | 0.7667 | 0.4738 | 0.2840 | 0.4925 | 69 |
| V76 | -1.4930 | 1.5681 | 0.4010 | -0.4236 | 0.2474 | 0.7857 | 0.5598 | 0.3070 | 0.4306 | 31 |
| V77 | 0.2232 | -1.9715 | -0.2916 | 0.9274 | 0.5067 | 0.3742 | 0.4456 | 0.5643 | 0.4713 | 59 |
| V78 | -0.9094 | 1.0281 | 0.1685 | 0.0851 | 0.3356 | 0.7229 | 0.5214 | 0.4038 | 0.4648 | 53 |
| V79 | -0.0005 | 0.7317 | 1.1602 | -0.9947 | 0.4729 | 0.6884 | 0.6849 | 0.1982 | 0.5266 | 86 |
| V80 | 0.8224 | 2.6255 | -0.6271 | -0.7117 | 0.5972 | 0.9086 | 0.3903 | 0.2521 | 0.6110 | 130 |
| V81 | 1.2214 | -2.9115 | -0.8400 | -2.0352 | 0.6574 | 0.2649 | 0.3552 | 0.0000 | 0.4493 | 40 |
| V82 | 0.8209 | 0.5664 | -0.5639 | -0.7469 | 0.5969 | 0.6692 | 0.4007 | 0.2454 | 0.5517 | 107 |
| V83 | -0.4357 | -1.2831 | 0.8076 | 1.5209 | 0.4071 | 0.4542 | 0.6268 | 0.6773 | 0.4769 | 62 |
| V84 | -0.5671 | -0.8329 | 0.4943 | -0.1860 | 0.3873 | 0.5066 | 0.5751 | 0.3522 | 0.4388 | 34 |
| V85 | 1.2493 | 1.3814 | 0.0058 | -0.8098 | 0.6617 | 0.7640 | 0.4946 | 0.2334 | 0.6198 | 136 |
| V86 | 0.3429 | 1.1338 | -0.7069 | -1.0188 | 0.5247 | 0.7352 | 0.3772 | 0.1936 | 0.5228 | 83 |
| V87 | 0.1775 | 0.9740 | -0.4391 | 0.4596 | 0.4998 | 0.7166 | 0.4213 | 0.4752 | 0.5409 | 96 |
| V88 | 0.8198 | -0.3046 | -0.1972 | 0.4407 | 0.5968 | 0.5680 | 0.4612 | 0.4716 | 0.5581 | 109 |
| V89 | -1.2621 | 0.1656 | 0.4366 | -1.3000 | 0.2823 | 0.6226 | 0.5656 | 0.1400 | 0.3908 | 16 |
| V90 | 0.3801 | -0.0729 | 1.1570 | -0.2461 | 0.5304 | 0.5949 | 0.6844 | 0.3408 | 0.5474 | 100 |
| V91 | 0.7566 | 1.3846 | 0.2228 | -0.2748 | 0.5872 | 0.7643 | 0.5304 | 0.3353 | 0.5975 | 125 |
| V92 | 0.9868 | 0.3051 | 0.5359 | -0.3856 | 0.6220 | 0.6389 | 0.5820 | 0.3142 | 0.5885 | 119 |
| V93 | -0.6307 | 0.0425 | 1.6762 | -1.1183 | 0.3777 | 0.6083 | 0.7699 | 0.1746 | 0.4670 | 56 |
| V94 | 1.7287 | -0.0029 | -0.6032 | -0.9731 | 0.7341 | 0.6030 | 0.3943 | 0.2023 | 0.5996 | 128 |
| V95 | 0.7446 | 1.2538 | -0.3449 | -0.7549 | 0.5854 | 0.7491 | 0.4368 | 0.2439 | 0.5705 | 115 |
| V96 | 0.8910 | -0.6957 | -1.4774 | -0.7926 | 0.6075 | 0.5225 | 0.2502 | 0.2367 | 0.4991 | 71 |
| V97 | 1.0161 | 1.5727 | -0.8139 | -0.4504 | 0.6264 | 0.7862 | 0.3595 | 0.3018 | 0.5964 | 124 |
| V98 | -2.4021 | 1.1652 | 1.6025 | 0.2695 | 0.1101 | 0.7388 | 0.7578 | 0.4390 | 0.3894 | 15 |
| V99 | 0.8537 | 0.3506 | -0.3481 | 0.1239 | 0.6019 | 0.6441 | 0.4363 | 0.4112 | 0.5701 | 114 |
| V100 | 0.2219 | 1.2902 | -1.8862 | 1.0882 | 0.5065 | 0.7534 | 0.1828 | 0.5949 | 0.5339 | 90 |
| V101 | 2.1516 | -0.8218 | 0.6916 | 1.4801 | 0.7979 | 0.5078 | 0.6077 | 0.6695 | 0.6861 | 153 |
| V102 | 0.2359 | 0.3941 | 1.0508 | 1.9153 | 0.5086 | 0.6492 | 0.6669 | 0.7524 | 0.5907 | 121 |
| V103 | 1.5819 | -1.5365 | -0.2466 | 1.0536 | 0.7119 | 0.4248 | 0.4530 | 0.5883 | 0.5921 | 122 |
| V104 | -0.9350 | -0.7984 | 1.6214 | 1.7402 | 0.3317 | 0.5106 | 0.7609 | 0.7191 | 0.4751 | 61 |
| V105 | -0.9468 | -0.8273 | 0.4000 | -0.4906 | 0.3299 | 0.5072 | 0.5596 | 0.2942 | 0.4016 | 19 |
| V106 | 1.2936 | -0.4993 | 1.2425 | 0.8576 | 0.6683 | 0.5453 | 0.6984 | 0.5510 | 0.6293 | 143 |
| V107 | 0.8802 | -3.0949 | 0.1413 | 1.0083 | 0.6059 | 0.2436 | 0.5170 | 0.5797 | 0.5004 | 72 |
| V108 | 1.7261 | 2.4421 | -0.6870 | -1.1093 | 0.7337 | 0.8873 | 0.3805 | 0.1764 | 0.6660 | 148 |
| V109 | 0.0716 | -0.8917 | 1.1465 | -1.0074 | 0.4838 | 0.4997 | 0.6826 | 0.1958 | 0.4843 | 65 |
| V110 | 1.1676 | -0.6450 | 0.4010 | -1.4996 | 0.6493 | 0.5284 | 0.5598 | 0.1020 | 0.5495 | 105 |
| V111 | 2.2048 | -2.3144 | 0.4298 | 0.5973 | 0.8060 | 0.3343 | 0.5645 | 0.5014 | 0.6233 | 138 |
| V112 | 1.6566 | 0.4933 | -0.7047 | -1.3796 | 0.7232 | 0.6607 | 0.3775 | 0.1249 | 0.5981 | 126 |
| V113 | 1.0447 | -1.7958 | -1.3696 | -1.3406 | 0.6307 | 0.3946 | 0.2680 | 0.1323 | 0.4703 | 58 |
| V114 | 2.5698 | -1.6432 | -0.9985 | -1.3906 | 0.8611 | 0.4123 | 0.3291 | 0.1228 | 0.5995 | 127 |
| 品种 Variety | Z1 | Z2 | Z3 | Z4 | μ(X1) | μ(X2) | μ(X3) | μ(X4) | 综合评价值 Comprehensive assessment value (D) | 排名 Order |
| V115 | 0.7084 | -0.2833 | -0.6263 | 1.3865 | 0.5799 | 0.5704 | 0.3905 | 0.6517 | 0.5595 | 111 |
| V116 | 0.2016 | -0.9832 | -0.6574 | -1.4154 | 0.5034 | 0.4891 | 0.3853 | 0.1181 | 0.4434 | 38 |
| V117 | 0.3319 | 0.5234 | 1.0628 | -0.7934 | 0.5231 | 0.6642 | 0.6688 | 0.2365 | 0.5480 | 101 |
| V118 | -0.0904 | 1.9288 | -0.2509 | -1.7092 | 0.4593 | 0.8276 | 0.4523 | 0.0621 | 0.5089 | 74 |
| V119 | -0.6536 | -0.4628 | 1.4120 | -1.3866 | 0.3742 | 0.5496 | 0.7264 | 0.1235 | 0.4393 | 35 |
| V120 | 0.2346 | 1.2992 | -0.3514 | -1.0919 | 0.5084 | 0.7544 | 0.4358 | 0.1797 | 0.5257 | 85 |
| V121 | 0.1400 | 0.3997 | -0.1264 | -0.8832 | 0.4941 | 0.6498 | 0.4728 | 0.2194 | 0.5014 | 73 |
| V122 | -1.0625 | 1.0788 | 0.6917 | -0.7088 | 0.3125 | 0.7288 | 0.6077 | 0.2526 | 0.4503 | 43 |
| V123 | 0.2827 | 0.6421 | -0.1297 | -0.5724 | 0.5156 | 0.6780 | 0.4723 | 0.2786 | 0.5256 | 84 |
| V124 | 1.3192 | 0.3550 | 2.7395 | 1.6487 | 0.6722 | 0.6447 | 0.9452 | 0.7017 | 0.7052 | 155 |
| V125 | 1.0251 | -1.4867 | 2.1165 | -0.5607 | 0.6278 | 0.4305 | 0.8425 | 0.2808 | 0.5709 | 116 |
| V126 | -0.7859 | -0.5590 | -0.5303 | 2.7169 | 0.3542 | 0.5384 | 0.4063 | 0.9051 | 0.4652 | 54 |
| V127 | 0.6901 | -0.7110 | -0.2695 | 0.5643 | 0.5772 | 0.5207 | 0.4493 | 0.4951 | 0.5372 | 93 |
| V128 | -0.1924 | -2.1455 | 0.8792 | 0.0768 | 0.4439 | 0.3540 | 0.6386 | 0.4023 | 0.4433 | 36 |
| V129 | 2.0218 | -3.0902 | 0.8532 | 0.0116 | 0.7783 | 0.2441 | 0.6343 | 0.3898 | 0.5843 | 118 |
| V130 | 2.3030 | 0.6975 | 0.1622 | -0.7651 | 0.8208 | 0.6845 | 0.5204 | 0.2419 | 0.6854 | 152 |
| V131 | 1.1158 | 1.6194 | 0.8286 | -0.8007 | 0.6415 | 0.7916 | 0.6302 | 0.2351 | 0.6349 | 144 |
| V132 | -1.3467 | 0.5684 | -0.7876 | 0.9307 | 0.2695 | 0.6695 | 0.3639 | 0.5649 | 0.4134 | 25 |
| V133 | -0.8406 | -0.8182 | -0.4682 | 0.1958 | 0.3460 | 0.5083 | 0.4165 | 0.4249 | 0.4044 | 21 |
| V134 | 2.0695 | -4.6858 | 0.4021 | 0.8945 | 0.7855 | 0.0586 | 0.5599 | 0.5580 | 0.5492 | 104 |
| V135 | 0.6691 | 0.4128 | 0.1090 | -0.2092 | 0.5740 | 0.6514 | 0.5116 | 0.3478 | 0.5612 | 112 |
| V136 | -0.3019 | 1.5029 | 1.1814 | -0.1033 | 0.4273 | 0.7781 | 0.6884 | 0.3680 | 0.5442 | 98 |
| V137 | 1.0540 | 1.4276 | 1.2276 | 1.1156 | 0.6322 | 0.7693 | 0.6960 | 0.6001 | 0.6718 | 150 |
| V138 | 1.7374 | 0.5377 | -0.0778 | -1.1412 | 0.7354 | 0.6659 | 0.4809 | 0.1703 | 0.6243 | 139 |
| V139 | -0.0621 | -2.0903 | 0.5436 | -1.5520 | 0.4636 | 0.3604 | 0.5833 | 0.0920 | 0.4149 | 26 |
| V140 | 1.6572 | -1.9967 | -0.2135 | -0.3041 | 0.7233 | 0.3713 | 0.4585 | 0.3297 | 0.5581 | 110 |
| V141 | -1.2772 | 0.9526 | 1.0633 | -0.1421 | 0.2800 | 0.7141 | 0.6689 | 0.3606 | 0.4496 | 41 |
| V142 | 0.8626 | -0.1704 | -0.9908 | 0.6097 | 0.6032 | 0.5836 | 0.3304 | 0.5038 | 0.5511 | 106 |
| V143 | -0.4207 | 1.6479 | -0.2193 | -0.6341 | 0.4094 | 0.7950 | 0.4575 | 0.2669 | 0.4975 | 70 |
| V144 | -0.0266 | -0.0241 | 0.5838 | 1.4158 | 0.4689 | 0.6006 | 0.5899 | 0.6573 | 0.5380 | 94 |
| V145 | 0.0194 | 1.1904 | -0.9115 | 0.1998 | 0.4759 | 0.7418 | 0.3435 | 0.4257 | 0.5193 | 78 |
| V146 | 1.4047 | 0.5100 | -0.6107 | -0.9683 | 0.6851 | 0.6627 | 0.3930 | 0.2032 | 0.5895 | 120 |
| V147 | 3.4893 | -3.9982 | -0.5485 | -0.2163 | 1.0000 | 0.1386 | 0.4033 | 0.3464 | 0.6352 | 145 |
| V148 | 1.2168 | -0.6707 | 2.6366 | -0.8229 | 0.6567 | 0.5254 | 0.9282 | 0.2309 | 0.6158 | 133 |
| V149 | 3.4272 | -1.4023 | -0.4987 | 0.6199 | 0.9906 | 0.4404 | 0.4115 | 0.5057 | 0.7238 | 157 |
| V150 | 1.2416 | -0.0553 | -0.4547 | -1.4753 | 0.6605 | 0.5970 | 0.4187 | 0.1066 | 0.5538 | 108 |
| V151 | 1.2981 | -5.1901 | -0.5088 | 0.5695 | 0.6690 | 0.0000 | 0.4098 | 0.4961 | 0.4484 | 39 |
| V152 | 0.6812 | -0.4861 | 0.0443 | -1.3083 | 0.5758 | 0.5469 | 0.5010 | 0.1384 | 0.5126 | 75 |
| V153 | 1.7589 | -0.4240 | -1.2320 | -1.3231 | 0.7386 | 0.5541 | 0.2906 | 0.1356 | 0.5687 | 113 |
| V154 | 1.8504 | -0.3416 | 0.5364 | -1.5895 | 0.7524 | 0.5637 | 0.5821 | 0.0849 | 0.6121 | 131 |
| V155 | -3.0664 | 0.5577 | 2.7888 | 1.8880 | 0.0098 | 0.6682 | 0.9533 | 0.7472 | 0.3794 | 13 |
| V156 | -2.4860 | -0.5374 | -0.1214 | 2.2582 | 0.0974 | 0.5409 | 0.4737 | 0.8177 | 0.3349 | 6 |
| V157 | 2.9685 | -1.3375 | -0.4448 | 0.8433 | 0.9213 | 0.4479 | 0.4204 | 0.5483 | 0.6960 | 154 |
| V158 | 2.0892 | -2.2147 | 0.5843 | 0.0875 | 0.7885 | 0.3459 | 0.5900 | 0.4043 | 0.6106 | 129 |
| V159 | 1.5189 | -2.8861 | 1.9919 | -0.2344 | 0.7024 | 0.2679 | 0.8219 | 0.3430 | 0.5720 | 117 |
| V160 | 0.5692 | -0.6643 | 0.8973 | -0.3538 | 0.5589 | 0.5262 | 0.6416 | 0.3202 | 0.5368 | 92 |
| Weights | 0.5097 | 0.2504 | 0.1349 | 0.1050 |
新窗口打开|下载CSV
2.4.2 各品种耐低磷能力的聚类分析 为使本试验结果更加客观, 使用最大距离法对4个耐低磷综合指标的综合评价值D进行聚类(图1)。结果将实验中谷子种质分为4类。第I类为低磷敏感型品种, 耐低磷D值范围为0.2426~0.3493, 耐低磷能力最弱, 共9份; 第II类为低磷较敏感型品种, 耐低磷能力中等偏弱, 耐低磷D值范围为0.3663~0.5126, 共66份; 第III类为较耐低磷品种, 耐低磷能力中等偏强, 耐低磷D值范围为0.5177~ 0.6352, 共70份; 第IV类为耐低磷品种, 耐低磷能力最强, 耐低磷D值范围为0.6503~0.7556, 共15份。
图1
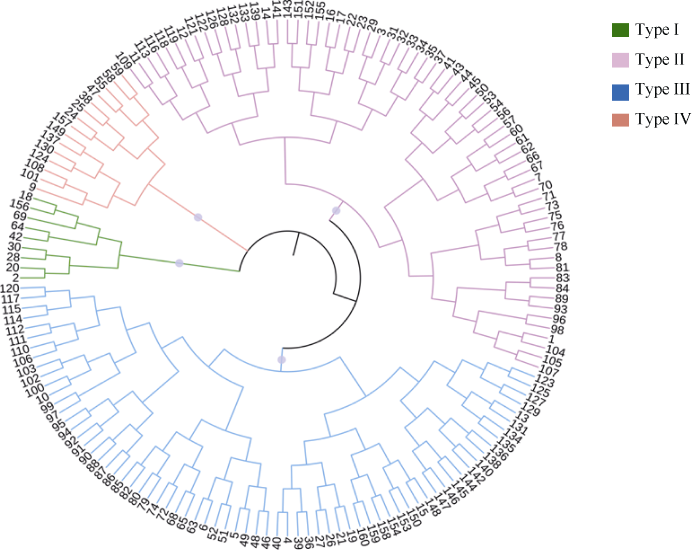 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1160个谷子品种的耐低磷性聚类图
Fig. 1Cluster of low-phosphorus tolerance in 160 foxtail millet varieties
2.5 不同品种在磷胁迫下的根系保护酶活性差异
逆境下, 植物会产生破坏细胞膜结构的超氧自由基。SOD是植物体内天然存在的自由基清除因子, 与POD和CAT结合把毒害的H2O2转换分解为完全无害的水, 保持植物正常的生长发育。这3种酶被认为是植物胁迫评估的重要生理指标。由于在LP环境下, 植物会优先将光合产物分配到地下部分, 地上部分生长受到抑制的同时地下部分促进根系发育[23]。因此, 我们根据筛选结果选择具有代表性的耐低磷品种(V47、V124、V130)与低磷敏感品种(V18、V69、V156)各3份进行水培, 测定其苗期根系SOD、POD、CAT活性。在NP环境下, SOD、POD和CAT根系酶活力平均值分别为282.83 U g-1 FW、914.80 U g-1 min-1 FW和3.51 U g-1 min-1 FW; LP环境下, 3种酶活力平均值分别为297.83 U g-1, FW、998.46 U g-1 min-1 FW和3.81 U g-1 min-1 FW。低磷胁迫促使6个谷子根系酶活力平均增加了5.30%、9.14%和8.54%。可见, 不同谷子根系在LP环境下相对于NP环境的SOD、POD、CAT活力均增大。与NP环境下相比, 低磷胁迫使V47、V124、V136、V18、V69和V156的SOD活力分别升高8.67%、6.20%、5.07%、4.42%、1.96%和5.74%。其中耐低磷品种V47增长幅度最大, 低磷敏感品种V69增长幅度最小, 6个谷子根系酶活力NP与LP处理间均达到显著差异。V47、V124、V136、V18、V69和V156在LP条件下的POD活力与NP条件下相比, 分别升高7.23%、14.61%、19.74%、5.02%、3.96%和3.29%。其中耐低磷品种V124和V136根系酶活力显著高于其余4个谷子品种。与NP环境下相比, 低磷胁迫使V47、V124、V136、V18、V69和V156的CAT活力分别升高5.41%、12.82%、7.91%、6.00%、1.05%和5.00%, 其中耐低磷品种V124增长幅度最大, 显著高于其余5个谷子根系酶活力, 低磷敏感品种V156增长幅度最小(图2)。图2
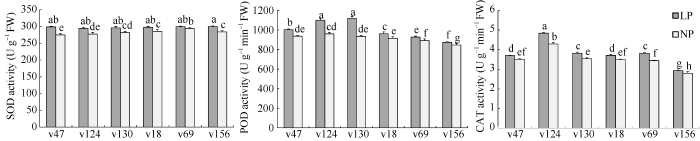 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同供磷强度下谷子SOD、POD、CAT活性的比较
图柱上不同的字母表示5%水平下差异的显著性。
Fig. 2Comparison of SOD, POD, CAT activities of foxtail millet under different phosphate conditions
Bars superscripted by different letters are significantly different at the 5% probability level.
3 讨论
3.1 筛选耐低磷种质资源的方法
苗期是谷子需磷量的瓶颈期和筛选低磷耐性指标的关键时期[24]。苗期筛选不仅能快速、准确地鉴定和筛选品种, 还可以显著减少田间筛选的工作量与成本[25]。而盆栽和水培是实施苗期试验简便有效的方法, 且精确度较高。因此本试验采取盆栽方法对160份谷子苗期耐低磷性进行种质筛选, 进一步采用水培方法对耐低磷和低磷敏感品种进行根系酶活性分析。3.2 苗期耐低磷综合指标的确定
谷子的耐低磷表现是一个复杂的过程, 缺磷会抑制植株某些生理特性, LP环境下地上部指标株高、地上部鲜重、叶长、地上部磷含量和地上部干重均小于NP环境下的各对应指标, 同时缺磷也会促进根系生长, 引起地下部鲜重增大。本试验通过主成分分析, 将所有单项指标划分为具有代表性的综合指标, 对160份核心谷子种质进行耐低磷能力的区分。结果表明, 缺磷对低磷敏感品种的影响要远远高于耐低磷品种, 由于部分耐低磷品种本身含有少量可用于谷物吸收的磷元素, 因而出现部分品种在LP环境下的茎粗、叶长等性状没有受到抑制反而促进生长的趋势, 表明磷元素的缺乏对不同基因型谷子苗期的单项指标有不同的推动作用, 为避免利用单个指标描述耐低磷的局限性, 最终采用4个综合指标分析不同品种的耐低磷特性。采用综合指标对作物的抗逆能力进行评价已在苜蓿[22]、花生[26]、水稻[27]等中取得了一定的成果。3.3 根系保护酶系统对低磷胁迫的响应
许多研究发现SOD、POD、CAT等酶类是植物抗逆的重要酶保护系统, 这3种酶在低磷胁迫下可以反映植物的逆境胁迫效应[28]。万美亮等[29]研究认为, 低磷胁迫诱导甘蔗体内保护酶系统活性升高, 是植物耐低磷的生理机制之一。本研究发现, 低磷环境诱导SOD、POD和CAT 3种酶活性增加以清除谷子幼苗根系内的H2O2, 但不同谷子根系酶活性的变化幅度存在一定差异, 与NP环境相比, LP环境中3种酶活性增幅最大的均为耐低磷品种, 增幅最小的均为低磷敏感性品种。表明低磷胁迫对不同基因型谷子根系酶活性具有不同的调控作用, 耐低磷品种和低磷敏感性品种在逆境下的胁迫响应存在差异, 因而清除植物体内产生的自由基的能力也不同。本研究结果与李俊等[17,30-32]对油菜、大豆、玉米等研究结果类似。4 结论
本研究对160份核心谷子种质的苗期耐低磷特性进行研究, 根据耐低磷能力强弱可分为低磷敏感型、低磷较敏感型、较耐低磷型和耐低磷型4种不同类型。低磷胁迫下, 本研究所测得的11个指标均可作为鉴定谷子苗期耐低磷能力和品种选育的参考指标。其中低磷耐性品种和低磷敏感品种根系保护酶系统对低磷胁迫响应能力的不同, 是谷子磷胁迫可能的重要逆境胁迫效应。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.gloenvcha.2008.10.009URL [本文引用: 1]

Food production requires application of fertilizers containing phosphorus, nitrogen and potassium on agricultural fields in order to sustain crop yields. However modern agriculture is dependent on phosphorus derived from phosphate rock, which is a non-renewable resource and current global reserves may be depleted in 50 100 years. While phosphorus demand is projected to increase, the expected global peak in phosphorus production is predicted to occur around 2030. The exact timing of peak phosphorus production might be disputed, however it is widely acknowledged within the fertilizer industry that the quality of remaining phosphate rock is decreasing and production costs are increasing. Yet future access to phosphorus receives little or no international attention. This paper puts forward the case for including long-term phosphorus scarcity on the priority agenda for global food security. Opportunities for recovering phosphorus and reducing demand are also addressed together with institutional challenges.
DOI:10.1016/j.foodpol.2010.11.012URL [本文引用: 1]
http://linkinghub.elsevier.com/retrieve/pii/S0306919210001260
DOI:10.1016/S0264-410X(02)00552-2URL [本文引用: 1]
The Green Revolution boosted crop yields in developing nations by introducing dwarf genotypes of wheat and rice capable of responding to fertilisation without lodging. We now need a second Green Revolution, to improve the yield of crops grown in infertile soils by farmers with little access to fertiliser, who represent the majority of thirdworld farmers. Just as the Green Revolution was based on crops responsive to high soil fertility, the second Green Revolution will be based on crops tolerant of low soil fertility. Substantial genetic variation in the productivity of crops in infertile soil has been known for over a century. In recent years we have developed a better understanding of the traits responsible for this variation. Root architecture is critically important by determining soil exploration and therefore nutrient acquisition. Architectural traits under genetic control include basal-root gravitropism, adventitious-root formation and lateral branching. Architectural traits that enhance topsoil foraging are important for acquisition of phosphorus from infertile soils. Genetic variation in the length and density of root hairs is important for the acquisition of immobile nutrients such as phosphorus and potassium. Genetic variation in root cortical aerenchyma formation and secondary development ('root etiolation') are important in reducing the metabolic costs of root growth and soil exploration. Genetic variation in rhizosphere modification through the efflux of protons, organic acids and enzymes is important for the mobilisation of nutrients such as phosphorus and transition metals, and the avoidance of aluminum toxicity. Manipulation of ion transporters may be useful for improving the acquisition of nitrate and for enhancing salt tolerance. With the noteworthy exceptions of rhizosphere modi. cation and ion transporters, most of these traits are under complex genetic control. Genetic variation in these traits is associated with substantial yield gains in low-f
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1081/PLN-100106973URL [本文引用: 1]
Intra-specific variations of phosphorus-utilization efficiency (PUE) hold great promise in rationalizing the application of cost-intensive phosphatic fertilizers in resource-poor situations, and to extend the useful life of the phosphate reserves in the world. In developed countries, efficient P-using crops may find significance in protecting the environment by reducing pollution of surface and ground water resources. This paper briefly reviews some of the work done so far on genotypic variations of Phosphorus (P)-nutrition of crops. Several physiological mechanisms responsible for such variations have been recognized. Various screening techniques to select P-efficient genotypes are also discussed. In addition, a brief historical account is also given on plant genetic approach to tackle soil problems.
DOI:10.1046/j.1469-8137.2003.00695.xURL [本文引用: 1]
Phosphorus (P) is limiting for crop yield on > 30% of the world's arable land and, by some estimates, world resources of inexpensive P may be depleted by 2050. Improvement of P acquisition and use by plants is critical for economic, humanitarian and environmental reasons. Plants have evolved a diverse array of strategies to obtain adequate P under limiting conditions, including modifications to root architecture, carbon metabolism and membrane structure, exudation of low molecular weight organic acids, protons and enzymes, and enhanced expression of the numerous genes involved in low-P adaptation. These adaptations may be less pronounced in mycorrhizal-associated plants. The formation of cluster roots under P-stress by the nonmycorrhizal species white lupin (Lupinus albus), and the accompanying biochemical changes exemplify many of the plant adaptations that enhance P acquisition and use. Physiological, biochemical, and molecular studies of white lupin and other species response to P-deficiency have identified targets that may be useful for plant improvement. Genomic approaches involving identification of expressed sequence tags (ESTs) found under low-P stress may also yield target sites for plant improvement. Interdisciplinary studies uniting plant breeding, biochemistry, soil science, and genetics under the large umbrella of genomics are prerequisite for rapid progress in improving nutrient acquisition and use in plants.
[本文引用: 1]
[本文引用: 1]
DOI:10.1371/journal.pone.0108459URLPMID:4177549 [本文引用: 2]
Abstract Phosphorus (P) is an essential element which plays several key roles in all living organisms. Setaria italica (foxtail millet) is a model species for panacoid grasses including several millet species widely grown in arid regions of Asia and Africa, and for the bioenergy crop switchgrass. The growth responses of S. italica to different levels of inorganic phosphate (Pi) and to colonisation with the arbuscular mycorrhizal fungus Funneliformis mosseae (syn. Glomus mosseae) were studied. Phosphate is taken up from the environment by the PHT1 family of plant phosphate transporters, which have been well characterized in several plant species. Bioinformatic analysis identified 12 members of the PHT1 gene family (SiPHT1;1-1;12) in S. italica, and RT and qPCR analysis showed that most of these transporters displayed specific expression patterns with respect to tissue, phosphate status and arbuscular mycorrhizal colonisation. SiPHT1;2 was found to be expressed in all tissues and in all growth conditions tested. In contrast, expression of SiPHT1;4 was induced in roots after 15 days growth in hydroponic medium of low Pi concentration. Expression of SiPHT1;8 and SiPHT1;9 in roots was selectively induced by colonisation with F. mosseae. SiPHT1;3 and SiPHT1;4 were found to be predominantly expressed in leaf and root tissues respectively. Several other transporters were expressed in shoots and leaves during growth in low Pi concentrations. This study will form the basis for the further characterization of these transporters, with the long term goal of improving the phosphate use efficiency of foxtail millet.
[本文引用: 2]
[本文引用: 2]
DOI:10.1016/S0378-4290(02)00061-8URL [本文引用: 1]
A solution culture experiment was conducted on five wheat cultivars and nine nutrient levels of cadmium (Cd) to determine the genotypic differences on biomass production, yield and yield components of wheat. Addition of 1 mg l 1 of Cd significantly reduced root and stem dry weight as well as tillers weight. The reduction rates were different among wheat genotypes. Number of grains per ear and single grain weight, was not reduced consistently in five genotypes because of the compensation among yield components. The accumulation of Cd in roots was 20 times higher than shoots and about 200 times higher than grains. A significant difference among genotypes existed in response of Cd concentration in various plant parts to Cd addition. The effect of Cd addition on nutrient concentrations in plants varied among elements, plant parts and genotypes. Cd treatment caused more P, K and Mn accumulation in roots, probably due to the inhibition of their translocation to plant shoots.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.1071/fp03060URL [本文引用: 1]
A field trial, a lysimeter system study and a nutrient solution experiment were conducted to determine the genotypic differences in (N) among (L.) genotypes differing in vigour of early . Plant and N of Vigour 18, a breeding line with early vigour, and the commercial cultivars Westonia, Tincurrin, Camm and Janz were compared. Shoot biomass of Vigour 18 was higher than that of the other genotypes, except for Westonia at booting when 50 kg N ha1 was applied 3 d after emergence. Vigour 18 had significantly higher efficiency of fertiliser-N than the other four cultivars at tillering when 50 kg N ha1 was applied. Fertiliser-N efficiency at booting was similar in Vigour 18 and Westonia, but significantly higher than in three other commercial cultivars. Vigour 18 had higher root dry matter, root-length density and root surface area than Janz when examined in columns of soil. The greater root of Vigour 18 occurred across all soil layers to a depth of 0.6 m. Differences in total N between Vigour 18 and Janz were apparent from tillering (Z14,22) to booting (Z19,24,49). Vigour 18 also had significantly higher shoot biomass and N than Janz when grown in nutrient culture. activity (NRA) expressed on a whole-plant basis was higher for Vigour 18 than for Janz, and was related to total N . However, NRA expressed on a per-unit-fresh-weight basis was not significantly different across the cultivars tested. It is concluded that vigorous early root and shoot in Vigour 18 was the main driving force for higher N .
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}