关键词:经验模型; 作物模型; 生长方程; 气候变化; 分蘖 Analysis and Simulation of Impact of Light and Temperature on Rice Tillering WANG Meng-Meng1,2, YANG Shen-Bin1,2,*, JIANG Xiao-Dong1,2, WANG Ying-Ping3, CHEN De2, HUANG Wei2, YU Geng-Kang4, SHI Chun-Lin5 1 Collaborative Innovation Center on Forecast and Evaluation of Meteorological Disasters / Jiangsu Key Laboratory of Agricultural Meteorology, Nanjing University of Information Science & Technology, Nanjing 210044, China
2 College of Applied Meteorology, Nanjing University of Information Science and Technology, Nanjing 210044, China
3 CSIRO Marine and Atmospheric Research, PMB # 1, Aspendale, Victoria 3195, Australia
4 Jiangsu Provincial Meteorological Bureau, Nanjing 210008, China
5 Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China
Fund:This study was supported by Special Fund for Meteorological-scientific Research in the Public Interest (GYHY201306035, GYHY2013 06036, GYHY201206020) and Key Projects in the National Science & Technology Pillar Program during the Twelfth Five-year Plan Period (2011BAD32B01) and the Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions AbstractIn order to investigate the effects of light and temperature on rice tillering dynamics in tillering stage and validate the light and temperature effect equation in current dynamic tillering models. We performed two-year field-seeding experiments (2012-2013) using two indica two-line hybrid rice cultivars, Lingliangyou 268 and Liangyoupeijiu. And we collected the observation data in tillering. To fit Richards equation for retrieving characteristic parameters related to the maximum tiller density, average growth rate of tillers, and duration of the growing period. Moreover, we analyzed the relationships between all these characteristic parameters and average data of light, temperature and climatic variables during the growing period for each rice cultivar. Based on the relationships we established co-effect equations for the growth rate of tillers and maximum tiller density as a function of co-effect of light and temperature. The new co-effect functions for each cultivar were substituted for the effect functions of light and temperature in a widely used dynamic tillering model to simulate the dynamic tillering in tillering stage. Finally, validation and comparison were carried out for the models applied observation data in tillers period. The result showed that the light and temperature affected crop growth in tillering period. The growth rate of tillers and the actual maximum tiller density positively correlated with the two meteorological factors significantly. Compared with current dynamic tillering model, the model with the established co-effects of light and temperature functions decreased the error significantly in simulating the growth dynamics of tillers. The simulated rice tillering was well consistent with the observed in both growth rate and maximum tiller density for both rice cultivars. However, the discrepancy could also be found in some seeding periods and validation samples, which may be caused by rice adaptability to different light and temperature environments. In conclusion, the validation and improvement of the light and temperature co-effect functions put forward in this study can be used further for understanding the effects of light and temperature factors on rice tillering and improving dynamic tillering models in the future.
Keyword:Empirical model; Crop model; Growth equation; Climate change; Tillering Show Figures Show Figures
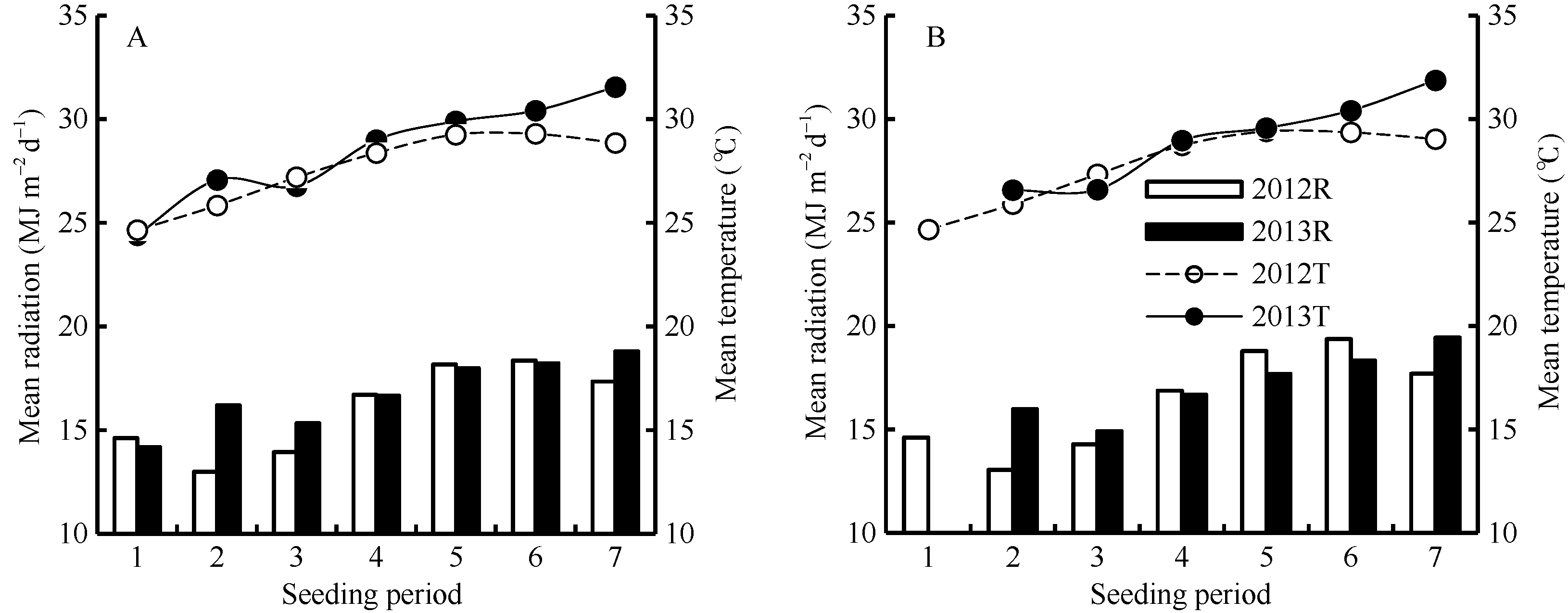
图1 2012-2013年陵两优268(A)和两优培九(B)各播期群体茎蘖增长期内平均光照与气温Fig. 1 Average daily total radiation and mean temperature during increasing period of tillering for Lingliangyou 268 (A) and Liangyoupeijiu (B) with different seeding periods in 2012 and 2013, respectively
表1 陵两优268各播期群体茎蘖生长参数和增长期内光温要素的方差分析 Table 1 Variance analysis of tillering characteristic parameters and radiation and temperature variables between seeding periods for Lingliangyou 268
年份 Year
播期 Seeding period
水稻群体茎蘖生长参数TCP
增长期内平均光温要素 ADTR
A (tiller m-2)
D (d)
Gm (tiller m-2 d-1)
T (℃)
R (MJ m-2d-1)
2012
S1
261.44 a
45.00 c
5.32 a
24.65 a
14.61 b
S2
238.81 a
24.67 a
9.15 c
25.83 b
13.00 a
S3
266.58 a
40.00 bc
6.13 ab
27.19 c
13.93 b
S4
324.42 b
42.67 c
6.97 b
28.37 d
16.70 c
S5
413.83 c
37.33 b
10.30 d
29.26 e
18.16 d
S6
403.20 c
35.67 b
10.51 d
29.29 f
18.36 d
S7
332.55 b
36.67 b
8.39 c
28.86 e
17.34 c
平均值 Mean
320.12
37.43
8.11
27.64
16.02
2013
S1
203.54 a
31.00 a
5.85 ab
24.41 a
14.18 a
S2
236.00 ab
49.33 b
4.22 a
27.07 b
16.19 c
S3
258.62 bc
32.33 a
6.98 bc
26.77 b
15.33 b
S4
244.10 abc
37.67 a
5.93 ab
29.00 c
16.66 c
S5
326.14 d
34.00 a
8.89 cd
29.90 d
17.99 d
S6
295.25 d
29.33 a
9.41 d
30.41 e
18.23 de
S7
463.63 e
33.00 a
13.12 e
31.55 f
18.80 e
平均值 Mean
289.61
35.24
7.77
28.44
16.77
S1-S7: seeding periods from 1 to 7; A: the maximum tiller density; Gm: the average growth rate of tillers; D: growth period; T: daily mean temperature; R: daily total radiation. ADTR: average daily total radiation and mean temperature during the tillering phase; TCP: tillering characteristic parameters. Values followed by different letters are significantly different at the 0.05 probability level. S1~S7: 第1至第7播期; A: 群体茎蘖密度最大值; Gm: 平均群体茎蘖增长速率; D: 群体茎蘖增长期; T: 茎蘖增长期温度平均值; R: 茎蘖增长期光照平均值。表中标以不同小写字母的各处理数值在0.05水平上的差异显著。
表1 陵两优268各播期群体茎蘖生长参数和增长期内光温要素的方差分析 Table 1 Variance analysis of tillering characteristic parameters and radiation and temperature variables between seeding periods for Lingliangyou 268
表2 Table 2 表2(Table 2)
表2 两优培九各播期群体茎蘖生长系数和茎蘖增长期光温要素的方差分析 Table 2 Variance analysis of tillering characteristic parameters and radiation and temperature variables between seeding periods for Liangyoupeijiu
年份 Year
播期 Seeding period
水稻群体茎蘖生长参数TCP
增长期内平均光温要素 ADTR
A (tiller m-2)
D (d)
Gm (tiller m-2 d-1)
T (℃)
R (MJ m-2 d-1)
2012
S1
378.79 a
42.67 d
8.08 a
24.66 a
14.62 b
S2
340.52 a
29.00 a
11.12 ab
25.89 b
13.05 a
S3
373.13 a
38.67 cd
8.87 a
27.33 c
14.29 b
S4
471.10 bc
38.33 cd
11.31 ab
28.17 d
16.87 c
S5
450.03 b
32.33 ab
13.01 b
29.40 f
18.79 d
S6
518.11 c
30.33 a
16.44 c
29.35 f
19.36 d
S7
521.50 c
36.00 bc
13.25 b
29.03 e
17.69 c
平均值 Mean
436.17
35.33
11.73
27.77
16.38
2013
S2
363.47 a
44.33 b
7.82 a
26.56 a
15.98 b
S3
383.71 a
31.00 a
11.27 b
26.59 a
14.92 a
S4
307.87 a
37.67 ab
7.38 a
28.96 b
16.69 c
S5
370.57 a
28.67 a
11.80 b
29.57 b
17.68 d
S6
371.27 a
29.00 a
12.10 b
30.42 c
18.33 e
S7
524.80 b
28.00 a
17.98 c
31.87 d
19.43 f
平均值 Mean
386.95
33.11
11.39
28.99
17.17
缩写同表1。Abbreviations are the same as those given in Table 1.
表2 两优培九各播期群体茎蘖生长系数和茎蘖增长期光温要素的方差分析 Table 2 Variance analysis of tillering characteristic parameters and radiation and temperature variables between seeding periods for Liangyoupeijiu
表3 Table 3 表3(Table 3)
表3 陵两优268群体茎蘖生长参数与光温要素的相关系数 Table 3 Correlation coefficients between tillering characteristic parameters and radiation and temperature factors for Lingliangyou 268
D
Rsum
Tsum
Ncold
Nheat
Tcold
Theat
R
T
Gm
A
-0.05
0.47
0.34
-0.68* *
0.64*
-0.67* *
0.65*
0.78* *
0.76* *
0.84* *
Gm
-0.56*
-0.01
-0.10
-0.82* *
0.68*
-0.75* *
0.70*
0.61*
0.71* *
T
-0.13
0.40
0.31
-0.90* *
0.89* *
-0.91* *
0.86* *
0.89* *
R
0.05
0.55*
0.34
-0.77* *
0.79* *
-0.71* *
0.75* *
Theat
-0.17
0.45
0.38
-0.79* *
0.99* *
-0.68*
Tcold
0.38
-0.11
-0.06
0.94* *
-0.72* *
Nheat
-0.16
0.46
0.38
-0.81* *
Ncold
0.52
-0.02
0.09
Tsum
0.83* *
0.95* *
Rsum
0.80* *
A: the maximum tiller density; Gm: the average growth rate of tillers; D: growth period; T: daily mean temperature; R: daily total solar radiation; Theat: hot accumulated temperature; Tcold: cold accumulated temperature; Nheat: number of days that daily maximum temperature ≥ 35℃; Ncold: number of days that daily mean temperature ≤ 24℃; Tsum: accumulative daily temperature; Rsum: accumulative daily solar radiation. * * Correlation is significant at the 0.01 probability level; * Correlation is significant at the 0.05 probability level. Sample size n=14. A: 群体茎蘖密度最大值; Gm: 平均群体茎蘖增长速率; D: 群体茎蘖增长期; T: 茎蘖增长期温度平均值; R: 茎蘖增长期光照平均值; Theat: 热积温; Tcold: 冷积温; Nheat: 日最高气温≥ 35℃出现的日数; Ncold: 日平均气温≤ 24℃出现的日数; Tsum: 日平均气温总和; Rsum: 累积光照。* 表示在0.05水平上显著相关, * * 表示在0.01水平上显著相关。统计样本量n=14。
表3 陵两优268群体茎蘖生长参数与光温要素的相关系数 Table 3 Correlation coefficients between tillering characteristic parameters and radiation and temperature factors for Lingliangyou 268
表4 Table 4 表4(Table 4)
表4 两优培九群体茎蘖生长参数与光温要素的相关系数 Table 4 Correlation coefficients between tillering characteristic parameters and radiation and temperature factors for Liangyoupeijiu
D
Rsum
Tsum
Ncold
Nheat
Tcold
Theat
R
T
Gm
A
-0.23
-0.22
-0.04
-0.53
0.44
-0.47
0.44
0.65*
0.52
0.82* *
Gm
-0.72* *
-0.24
-0.53
-0.74* *
0.59*
-0.66*
0.59*
0.70* *
0.69* *
T
-0.56*
0.06
-0.17
-0.95* *
0.93* *
-0.91* *
0.92* *
0.88* *
R
-0.39
0.33
-0.04
-0.83*
0.80* *
-0.73*
0.80* *
Theat
-0.41
0.15
-0.05
-0.83*
1.00* *
-0.72* *
Tcold
0.65*
0.12
0.30
0.96* *
-0.73* *
Nheat
-0.41
0.16
-0.04
-0.83*
Ncold
0.69* *
0.08
0.33
Tsum
0.91* *
0.90* *
Rsum
0.74* *
缩写同表3。样本统计量n=13。Abbreviations are the same as those given in Table 3. Sample size n=13.
表4 两优培九群体茎蘖生长参数与光温要素的相关系数 Table 4 Correlation coefficients between tillering characteristic parameters and radiation and temperature factors for Liangyoupeijiu
表5 Table 5 表5(Table 5)
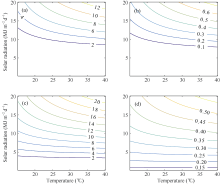
表5 光温组合影响方程 Table 5 Equations of combined effect from radiation and temperature for average growth rate of tillers Gm and final tillering rateLe
品种 Cultivar
Gm与光温组合因子的回归方程 Regression function relates Gm to K
Le与光温组合因子的回归方程 Regression function relates Le to K
陵两优268 Lingliangyou 268
FG(K)=1.817× K-10.688 (R2=0.49, n=19, P< 0.01)
FL(K)=0.095× K-0.523 (R2=0.58, n=21, P< 0.01)
两优培九 Liangyoupeijiu
FG(K)=1.916× K-3.921 (R2=0.98, n=18, P< 0.01)
FL(K)=0.034× K+0.124 (R2=0.698, n=18, P< 0.01)
表5 光温组合影响方程 Table 5 Equations of combined effect from radiation and temperature for average growth rate of tillers Gm and final tillering rateLe
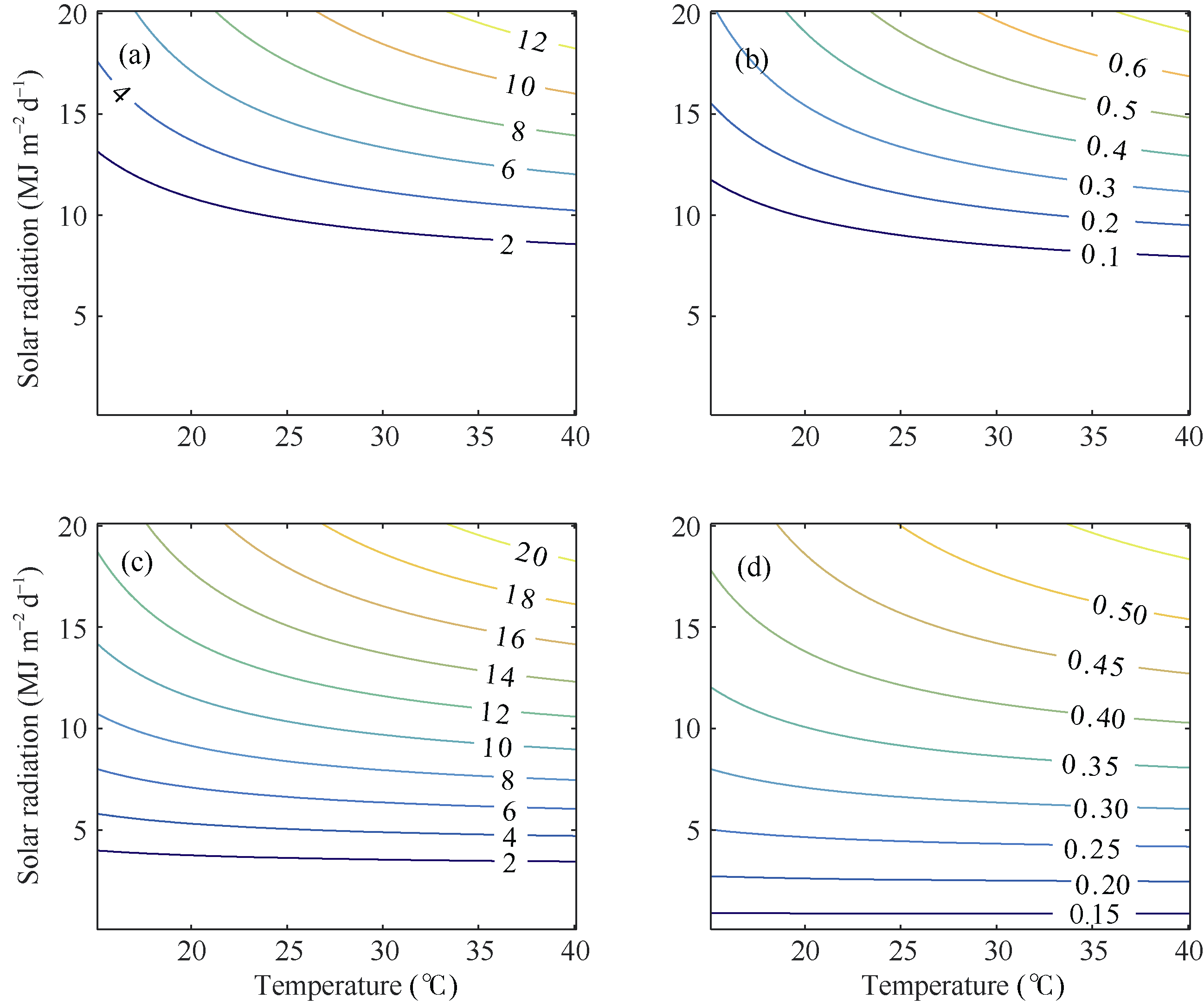
图2 两品种FG和FL的光温响应关系 (a)和(b)分别对应陵两优268的FG和FL; (c)和(d)分别对应两优培九的FG和FL。Fig. 2 Changes of FG and FL in response to radiation and temperature for both rice varieties (a) and (b) for FG and FL of Lingliangyou 268, respectively; (c) and (d) for FG and FL of Liangyoupeijiu, respectively.
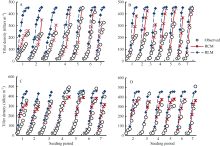
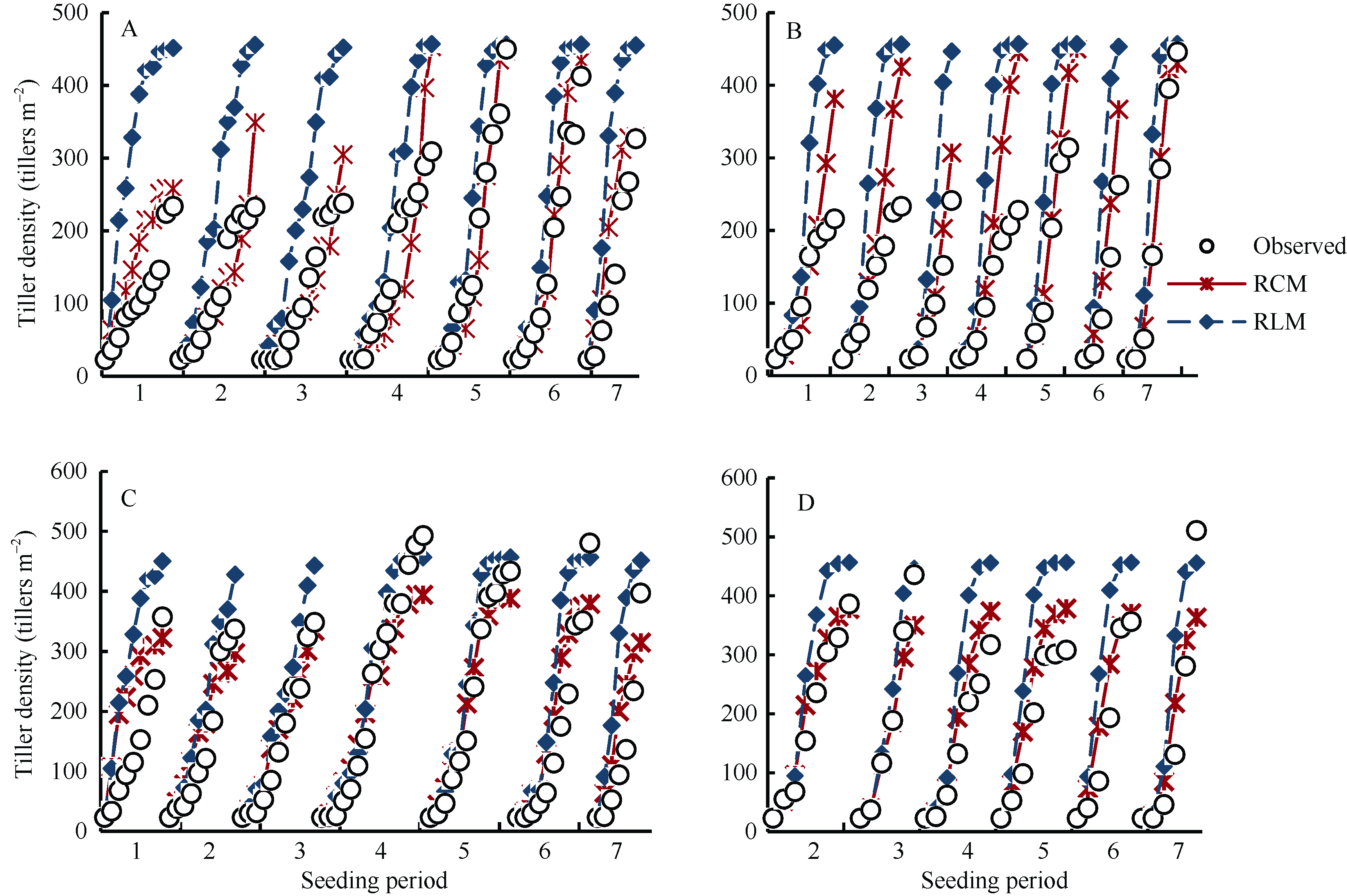
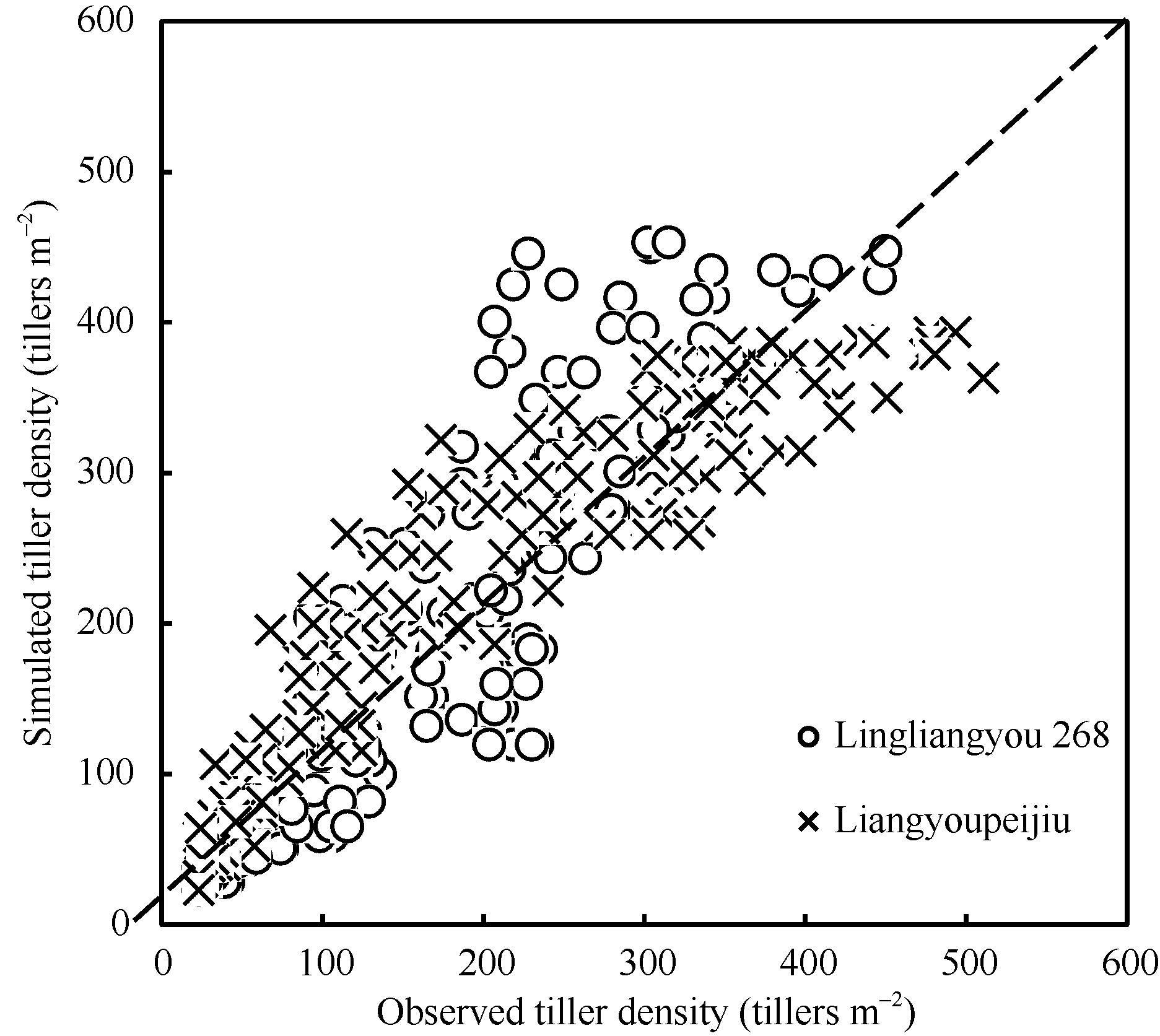
图3 陵两优268 2012年(A)和2013年(B)及两优培九2012年(C)和2013年(D)群体茎蘖动态模拟值与实测值的比较Fig. 3 Comparison between simulated tiller density and the observed for Lingliangyou 268 in 2012 (A) and 2013 (B), and for Liangyoupeijiu in 2012 (C) and 2013 (D), respectively
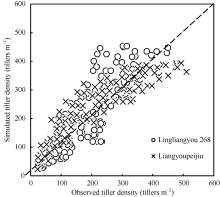
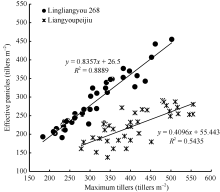
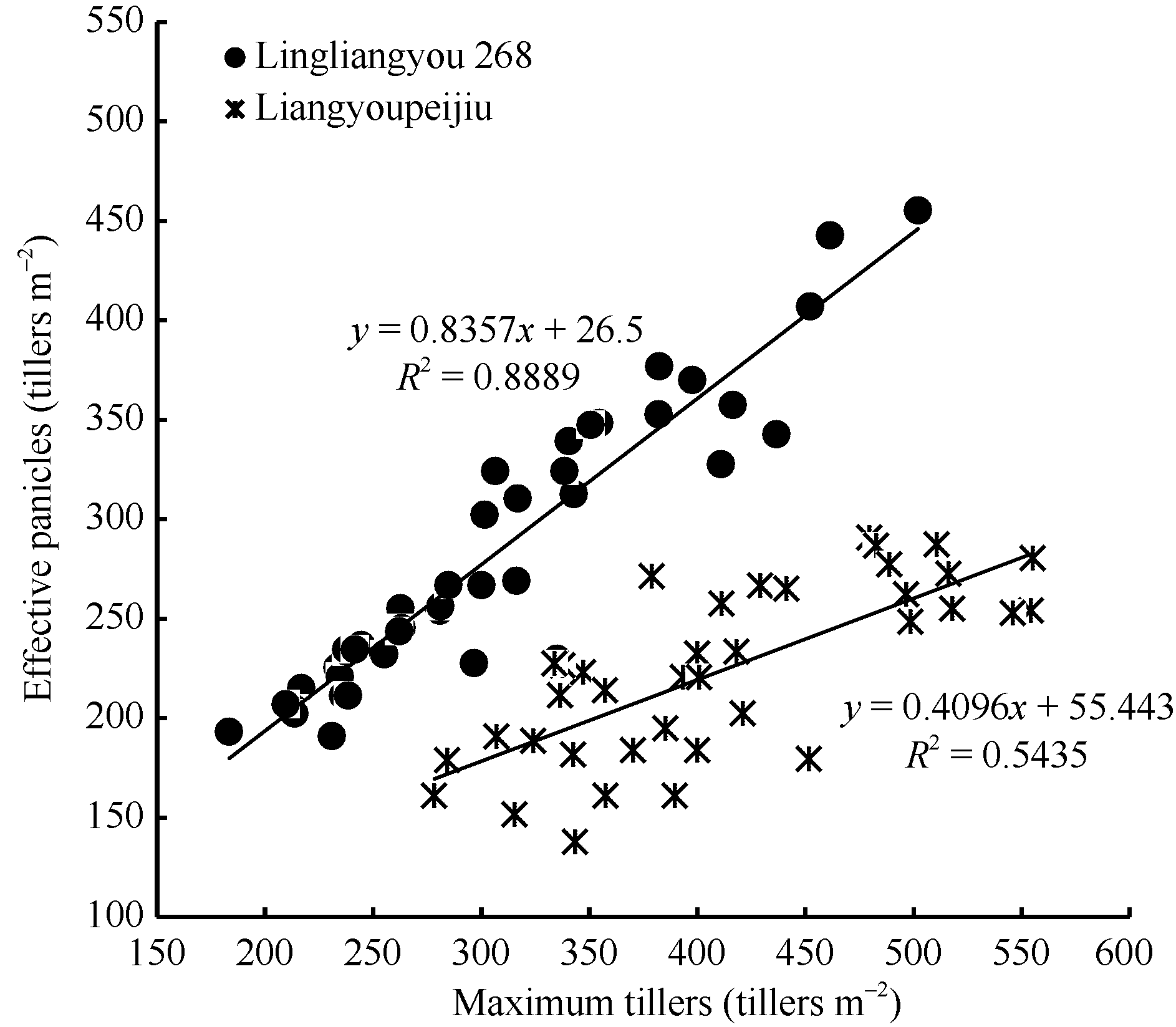
图5 陵两优268和两优培九群体最大茎蘖数与群体最大有效茎蘖数的关系Fig. 5 Comparison between observed number of effective panicles and maximum tiller density for Lingliangyou 268 and Liangyoupeijiu, respectively
HuangM, Zou YB, JiangP, XiaB, MdIbrahim, Ao HJ. Relationship between grain yield and yield components in super hybrid rice. Agric Sci China, 2011, 10(10): 1537-1544[本文引用:1]
[2]
蒋彭炎. 水稻分蘖的发生、控制与茎蘖成穗率的提高. 中国稻米, 1999, (4): 7-9Jiang PY. Tillering and its control and the improvement of panicles. China Rice, 1999, (4): 7-9 (in Chinese)[本文引用:1]
[3]
Deng NY, Ling XX, SunY, Zhang CD, FahadS, Peng SB, Cui KH, Nie LX, Huang JL. Influence of temperature and solar radiation on grain yield and quality in irrigated rice system. Eur J Agron, 2015, 64: 37-46[本文引用:1]
[4]
敖和军, 王淑红, 邹应斌, 彭少兵, 唐启源, 方远祥, 肖安民, 陈玉梅, 熊昌明. 超级杂交稻干物质生产特点与产量稳定性研究. 中国农业科学, 2008, 41: 1927-1936Ao HJ, Wang SH, Zou YB, Peng SB, Tang QY, Fang YX, Xiao AM, Chen YM, Xiong CM. Study on yield stability and dry matter characteristics of super hybrid rice. Agric Sci China, 2008, 41: 1927-1936 (in Chinese with English abstract)[本文引用:1]
[5]
张恒栋, 张发丽, 石明, 钱晓刚. 不同移栽群体和水分调控对水稻分蘖成穗的影响. 中国农学通报, 2013, 29(6): 20-23Zhang HD, Zhang FL, ShiM, Qian XG. The effects of different transplanting groups and water control on formation of spike from the tillers in rice. Chin Agric Sci Bull, 2013, 29(6): 20-23 (in Chinese with English abstract)[本文引用:2]
[6]
PasuquinE, LafargeT, TubanaB. Transplanting young seedlings in irrigated rice fields: early and high tiller production enhanced grain yield. Field Crops Res, 2008, 105: 141-155[本文引用:1]
[7]
彭国照, 郝克俊. 四川盆地杂交水稻茎蘖动态及气象模拟模型. 西南农业大学学报(自然科学版), 2003, 25: 243-247Peng GZ, Hao KJ. Stem density dynamics of hybrid rice production in Sichuan Basin and simulated meteorological models for it. J Southwest Agric Univ (Nat Sci), 2003, 25: 243-247 (in Chinese with English abstract)[本文引用:1]
[8]
Ao HJ, Peng SB, Zou YB, Tang YX, Visperas RM. Reduction of unproductive tillers did not increase the grain yield of irrigated rice. Field Crops Res, 2010, 116: 108-115[本文引用:1]
[9]
YuQ, HengsdijkH, Liu JD. Application of a progressive- difference method to identify climatic factors causing variation in the rice yield in the Yangtze Delta, China. Int J Biometeorol, 2001, 45: 53-58[本文引用:1]
[10]
邵玺文, 阮长春, 赵兰坡, 胡辉耀, 孙长占. 分蘖期水分胁迫对水稻生长发育及产量的影响. 吉林农业大学学报, 2005, 27(1): 6-10Shao XW, Ruan CC, Zhao LP, Hu HY, Sun CZ. Effects of water stress on growth and yield of rice in tillering stage. J Jilin Agric Univ, 2005, 27(1): 6-10 (in Chinese with English abstract)[本文引用:1]
[11]
黄耀, 高亮之, 金之庆, 陈华, 葛道阔. 水稻群体茎蘖动态的计算机模拟模型. 生态学杂志, 1994, 13(4): 27-32HuangY, Gao LZ, Jin ZQ, ChenH, Ge DK. Simulation model of tillering dynamics of rice community. Chin J Ecol, 1994, 13(4): 27-32 (in Chinese with English abstract)[本文引用:9]
[12]
苏祖芳, 张娟. 水稻群体茎蘖动态与成穗率和产量形成关系的研究. 江苏农学院学报, 1997, 18(1): 36-40Su ZF, ZhangJ. Study on relationship of tiller development of rice population with the effective ear percentage and rice formulation. J Jiangsu Agric Coll, 1997, 18(1): 36-40 (in Chinese with English abstract)[本文引用:3]
[13]
YoshidaS. Fundamentals of Rice Crop Science. Los Banos, Philippines: International Rice Research Institute, 1981. pp 70-76[本文引用:2]
[14]
KumagaiT, HidemaJ, Kang HS, SatoT. Effects of supplemental UV-B radiation on the growth and yield of two cultivars of Japanese lowland rice (Oryza sativa L. ) under the field in a cool rice-growing region of Japan. Agric Ecosyst Environ, 2001, 83: 201-208[本文引用:2]
[15]
王立志, 王春艳, 李忠杰, 李锐, 李禹尧, 孟英, 王连敏. 黑龙江水稻冷害Ⅳ分蘖期低温对水稻分蘖的影响. 黑龙江农业科学, 2009, (4): 18-20Wang LZ, Wang CY, Li ZJ, LiR, Li YY, MengY, Wang LM. Rice cooling injury in Heilongjiang Province: IV. Effect of low temperature on rice tillering. Heilongjiang Agric Sci, 2009, (4): 18-20 (in Chinese with English abstract)[本文引用:1]
[16]
姚克敏, 田红. 我国籼型杂交稻分蘖的生态类型及其利用. 南京气象学院学报, 1998, 11: 443-453Yao KM, TianH. Tillering ecologic characteristics of China hybrid indica rice and their application. J Nanjing Inst Meteorol, 1998, 11: 443-453 (in Chinese with English abstract)[本文引用:2]
[17]
刘瑞华, 崔贞玉, 冯权. 水稻分蘖期的温度条件与适宜移栽密度的研究. 吉林农业科学, 1993, (3): 40-43Liu RH, Cui ZY, FengQ. The study of temperature during the tillering phase and suitable transplanting density. J Jilin Agric Sci, 1993, (3): 40-43 (in Chinese with English abstract)[本文引用:1]
[18]
孙成明, 庄恒扬, 杨连新, 杨洪建, 黄建晔, 董桂春, 朱建国, 王余龙. 开放式空气CO2浓度增高对水稻茎蘖动态影响的模拟研究. 农业环境科学学报, 2006, 25: 1122-1126Sun CM, Zhuang HY, Yang LX, Yang HJ, Huang JY, Dong GC, Zhu JG, Wang YL. Simulation study on effects of free-air CO2 enrichment (FACE) on tiller dynamic of rice. J Agro-Environ Sci, 2006, 25: 1122-1126 (in Chinese with English abstract)[本文引用:2]
[19]
孙成明, 庄恒扬, 杨连新, 杨洪建, 黄建晔, 董桂春, 朱建国, 王余龙. FACE水稻茎蘖动态模型. 应用生态学报, 2006, 17: 1448-1452Sun CM, Zhuang HY, Yang LX, Yang HJ, Huang JY, Dong GC, Zhu JG, Wang YL. Dynamic model of rice tiller in FACE. Chin J Appl Ecol, 2006, 17: 1448-1452 (in Chinese with English abstract)[本文引用:2]
[20]
孟亚利, 曹卫星, 柳新伟, 周治国, 荆奇. 水稻茎蘖动态的模拟研究. 南京农业大学学报, 2003, 26(2): 1-6Meng YL, Cao WX, Liu XW, Zhou ZG, JingQ. Simulation on the tiller dynamic in rice. J Nanjing Agric Univ, 2003, 26(2): 1-6 (in Chinese with English abstract)[本文引用:1]
[21]
王夫玉, 黄丕生. 水稻群体茎蘖消长模型及群体分类研究. 中国农业科学, 1997, 30(1): 57-64Wang FY, Huang PS. Study on basic dynamic model for stem and tiller growth and population classification in rice. Agric Sci China, 1997, 30(1): 57-64 (in Chinese with English abstract)[本文引用:1]
[22]
严定春, 朱艳, 曹卫星, 王绍华. 水稻群体生长指标动态的知识模型研究. 中国农业科学, 2005, 38: 38-44Yan DC, ZhuY, Cao WX, Wang SH. A knowledge model for design of suitable dynamics of growth index in rice. Sci Agric Sin, 2005, 38: 38-44 (in Chinese with English abstract)[本文引用:1]
[23]
蒋德隆. 水稻分蘖与光、温条件关系的统计模式. 植物学报, 1982, 24: 247-251Jiang DL. The model of relations of rice tillering to light and temperature conditions. Acta Bot Sin, 1982, 24: 247-251 (in Chinese with English abstract)[本文引用:1]
[24]
Penning de Vries F W T. Simulation of Ecophysiological Processes of Growth in Several Annual Crops. Los Banos, Philippines: International Rice Research Institute, 1989[本文引用:2]
[25]
钟旭华, 彭少兵, Sheehy JE, 刘鸿先. 水稻群体成穗率与干物质积累动态关系的模拟研究. 中国水稻科学, 2001, 15: 107-112Zhong XH, Peng SB, Sheehy JE, Liu HX. Relationship between productive tiller percentage and biomass accumulation in rice (Oryza sativa L. ): a simulation approach. Chin J Rice Sci, 2001, 15: 107-112 (in Chinese with English abstract)[本文引用:2]
[26]
周劲松, 梁国华. 水稻分蘖性状的分子遗传研究进展. 江西农业学报, 2006, 18(1): 80-84Zhou JS, Liang GH. Advances in molecular genetics of tillering characters in rice. Acta Agric Jiangxi, 2006, 18(1): 80-84 (in Chinese with English abstract)[本文引用:1]
[27]
丁正生, 丁志芳, 朱从海, 刘广文. 不同播栽行距对水稻产量、产量构成及茎蘖动态的影响. 农技服务, 2012, 29(2): 130-130Ding ZS, Ding ZF, Zhu CH, Liu GW. Influence of different transplanting spaced on rice yield and yield components and tillering dynamic. Agric Technol Serv, 2012, 29(2): 130-130 (in Chinese)[本文引用:1]
[28]
Yan JQ, ZhuJ, He CX, BenmoussaM, WuP. Quantitative trait loci analysis for the developmental behavior of tiller number in rice (Oryza sativa L. ). Theor Appl Genet, 1998, 97: 267-274[本文引用:1]
[29]
凌启鸿, 苏祖芳, 张海泉. 水稻成穗率与群体质量的关系及其影响因素的研究. 作物学报, 1995, 21: 463-469Ling QH, Su ZF, Zhang HQ. Relationship between ear bearing tiller percentage and population quality and influence factors in rice. Acta Agron Sin, 1995, 21: 463-469 (in Chinese with English abstract)[本文引用:1]
[30]
Xu YB, Shen ZT. Diallel analysis of tiller number at different growth stages in rice (Oryza sativa L. ). Theor Appl Genet, 1991, 83(2): 243-249[本文引用:1]
[31]
梁康迳, 林文雄, 王雪仁, 陈志雄, 郭玉春, 梁义元, 陈芳育, 李亚娟. 籼型三系杂交水稻茎蘖数的发育遗传研究. 中国农业科学, 2002, 35: 1033-1039Liang KJ, Lin WX, Wang XR, Chen ZX, Guo YC, Liang YY, Chen FY, Li YJ. Studies on developmental genetics of the tiller numbers in three-line indica hybrid rice. Agric Sci China, 2002, 35: 1033-1039 (in Chinese with English abstract)[本文引用:1]
[32]
徐俊增, 彭世彰, 魏征. 分蘖期水分调控对覆膜旱作水稻茎蘖动态影响分析. 河海大学学报(自然科学版), 2010, 38: 511-515Xu JZ, Peng SZ, WeiZ. Effect of soil moisture regulation during tillering period on shoot dynamics of rice cultivated in plastic film mulched dryland and its simulation. J Hohai Univ (Nat Sci), 2010, 38: 511-515 (in Chinese with English abstract)[本文引用:1]
[33]
原晓明, 俞双恩, 谢俊英, 王艳艳. 农田水位调控对水稻茎蘖动态和株高的影响. 灌溉排水学报, 2011, 30(3): 56-59Yuan XM, Yu SE, Xie JY, Wang YY. Effects of controlled water level on the development of the tiller number and plant height. J Irrig & Drain, 2011, 30(3): 56-59 (in Chinese with English abstract)[本文引用:1]
[34]
刘杨, 王强盛, 丁艳锋, 王绍华. 水稻分蘖发生机理的研究进展. 中国农学通报, 2011, 27(3): 1-5LiuY, Wang QS, Ding YF, Wang SH. Advances in mechanisms of tiller occurs in rice. Chin Agric Sci Bull, 2011, 27(3): 1-5 (in Chinese with English abstract)[本文引用:1]
 , 江晓东
, 江晓东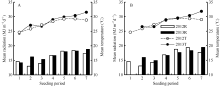
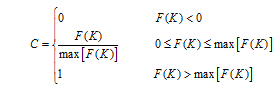
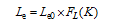

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}