关键词:萌发主要事件; 植物激素; 蛋白合成与翻译后修饰; 蛋白质组; 萌发的调节; 种子萌发 Research Progress in Seed Germination and Its Control XU Heng-Heng1, LI Ni2, LIU Shu-Jun1, WANG Wei-Qing1, WANG Wei-Ping2, ZHANG Hong1, CHENG Hong-Yan1, SONG Song-Quan1,* 1Institute of Botany, Chinese Academy of Sciences, Beijing 100093
2Hunan Hybrid Rice Research Center / State Key Laboratory of Hybrid Rice, Changsha 410125
Fund: AbstractMost flowering plants reproduce through sexual breeding and seed production. The success of seed germination and the establishment of a normal seedling determinate the propagation and survival of plant species. Seed germination is the most critical and flimsy phase in plant life cycle because of its high vulnerability to injury, disease and environmental stress, which are very important especially for annual and biannual plant species. Seed germination is a complex and multistep process during which the quiescent dry seeds quickly recover the metabolic activity, complete essential cellular events for the embryo to emerge from surrounding structures, and prepare for subsequent seedling growth. In the present paper, the recent research progress in seed germination and its control is reviewed, mainly including the important physiological events during germination, the protein synthesis, posttranslational modification and proteome associated with seed germination, and regulation of phytohormone on seed germination. Furthermore, we have also proposed an energy stimulating hypothesis for seed germination, which provides a new idea for decreasing the pre-harvest sprouting occurred in seed and foodstuff production of cereal crops and answering two fundamental questions in botany, agronomy, and horticulture: how does the embryo emerge from its surrounding structures to complete germination and how is embryo emergence blocked so that seeds can be maintained in the dormant state?
Keyword:Main events of germination; Phytohormone; protein synthesis and posttranslational modification; Proteome; Regula-tion of germination; Seed germination Show Figures Show Figures
RajjouL, DuvalM, GallardoK, CatusseJ, BallyJ, JobC, JobD. Seed germination and vigor. Annu Rev Plant Biol, 2012, 63: 507-533[本文引用:6]
[2]
WeitbrechtK, MüllerK, Leubner-MetzgerG. First off the mark: early seed germination. J Exp Bot, 2011, 62: 3289-3309[本文引用:4][JCR: 5.242]
[3]
BewleyJ D, BradfordK J, HilhorstH W M, NonogakiH. Physiology of Development, Germination and Dormancy, 3rd edn. New York: Springer, 2013[本文引用:11]
[4]
GublerF, MillarA A, JacobsenJ V. Dormancy release, ABA and pre-harvest sprouting. Curr Opin Plant Biol, 2005, 8: 183-187[本文引用:2][JCR: 8.455]
[5]
宋松泉. 种子休眠. “10000个科学难题”农业科学编委会. 10000个科学难题. 北京: 科学出版社, 2011. pp31-35SongS Q. Seed dormancy. In: The Editorial Board of Agricultural Science for 10000 Selected Problems in Sciences, ed. 10000 Selected Problems in Sciences. Beijing: Science Press, 2011. pp31-35(in Chinese)[本文引用:1]
[6]
NonogakiH, BasselG W, BewleyJ D. Germination—still a mystery. Plant Sci, 2010, 179: 574-581[本文引用:6][JCR: 2.922]
[7]
ObrouchevaN V, AntipovaO V. Physiology of the initiation of seed germination. Russian J Plant Physiol, 1997, 44: 250-264[本文引用:1]
[8]
KrishnanP, JoshiD K, NagarajanS, MoharirA V. Characterization of germinating and non-viable soybean seeds by nuclear magnetic resonance (NMR) spectroscopy. Seed Sci Res, 2004, 14: 355-362[本文引用:1][JCR: 1.931]
[9]
NonogakiH. Seed Germination and Reserve Mobilization. Encyclopedia of Life Sciences, Chichester: John Wiley & Sons Ltd, 2008[本文引用:1]
[10]
ManzB, MüllerK, KuceraB, VolkeF, Leubner-MetzgerG. Water uptake and distribution in germinating tobacco seeds investigated in vivo by nuclear magnetic resonance imaging. Plant Physiol, 2005, 138: 1538-1551[本文引用:1][JCR: 6.555]
[11]
RathjenJ R, StrouninaE V, MaresD J. Water movement into dormant and nondormant wheat (Triticum aestivum L. ) grains. J Exp Bot, 2009, 60: 1619-1631[本文引用:1][JCR: 5.242]
[12]
TerskikhV V, FeurtadoJ A, RenC, AbramsS A, KermodeA R. Water uptake and oil distribution during imbibition of seeds of western white pine (Pinus monticola Dougl. ex D. Don) monitored in vivo using magnetic resonance imaging. Planta, 2005, 221: 17-27[本文引用:1][JCR: 3.347]
[13]
RobertC, NoriegaA, TocinoA, CervantesE. Morphological analysis of seed shape in Arabidopsis thaliana reveals altered polarity in mutants of the ethylene signaling pathway. J Plant Physiol, 2008, 165: 911-919[本文引用:1][JCR: 2.699]
[14]
JoosenR V L, LigterinkW, DekkersB J W, HilhorstH W M. Visualization of molecular processes associated with seed dormancy and germination using MapMan. Seed Sci Res, 2010, 21: 143-152[本文引用:1][JCR: 1.931]
[15]
GreletJ, BenamarA, TeyssierE, Avelange-MacherelM H, GrunwaldD, MacherelD. Identification in pea seed mitochondria of a late-embryogenesis abundant protein able to protect enzymes from drying. Plant Physiol, 2005, 137: 157-167[本文引用:1][JCR: 6.555]
[16]
TolleterD, JaquinodM, MangavelC, PassiraniC, SaulnierP, ManonS, TeyssierE, PayetN, Avelange-MacherelM H, MacherelD. Structure and function of a mitochondrial late embryogenesis abundant protein are revealed by desiccation. Plant Cell, 2007, 19: 1580-1589[本文引用:1][JCR: 9.251]
[17]
BoubriakI, PouschukV, GrodzinskyA, OsborneD J. Telomeres and seed banks. Tsitologiia i Genetika, 2007, 41: 23-29[本文引用:1]
[18]
BrayC M, WestC E. DNA repair mechanisms in plants: crucial sensors and effectors for the maintenance of genome integrity. New Phytol, 2005, 168: 511-528[本文引用:1][JCR: 6.736]
[19]
WaterworthW M, MasnaviG, BhardwajR M, JiangQ, BrayC M, WestC. A plant DNA ligase is an important determinant of seed longevity. Plant J, 2010, 63: 848-860[本文引用:2][JCR: 6.582]
[20]
MacoveiA, BalestrazziA, ConfalonieriM, FaéM, CarboneraD. New insights on the barrel medic MtOGG1 and MtFPG functions in relation to oxidative stress response in planta and during seed imbibition. Plant Physiol Biochem, 2011, 49: 1040-1050[本文引用:1]
[21]
HuntL, HoldsworthM J, GrayJ E. Nicotinamidase activity is important for germination. Plant J, 2007, 51: 341-351[本文引用:2][JCR: 6.582]
[22]
DinkinsR D, MajeeS M, NayakN R, MartinD, XuQ, BelcastroM P, HoutzR L, BeachC M, DownieA B. Changing transcriptional initiation sites and alternative 5'- and 3'-splice site selection of the first intron deploys Arabidopsis PROTEIN ISOASPARTYL METHYLTRANSFERASE2 variants to different subcellular compartments. Plant J, 2008, 55: 1-13[本文引用:2][JCR: 6.582]
[23]
OgéL, BourdaisG, BoveJ, ColletB, GodinB, GranierF, BoutinJ P, JobD, JullienM, GrappinP. Protein repair L-isoaspartyl methyltransferase 1 is involved in both seed longevity and germination vigor in Arabidopsis. Plant Cell, 2008, 20: 3022-3037[本文引用:2][JCR: 9.251]
[24]
NakabayashiK, OkamotoM, KoshibaT, KamiyaY, NambaraE. Genome-wide profiling of stored mRNA in Arabidopsis thaliana seed germination: epigenetic and genetic regulation of transcription in seed. Plant J, 2005, 41: 697-709[本文引用:1][JCR: 6.582]
[25]
SreenivasuluN, UsadelB, WinterA, RadchukV, ScholzU, SteinN, WeschkeW, StrickertM, CloseT J, StittM, GranerA, WobusU. Barley grain maturation and germination: metabolic pathway and regulatory network commonalities and differences highlighted by new MapMan/PageMan profiling tools. Plant Physiol, 2008, 146: 1738-1758[本文引用:1][JCR: 6.555]
[26]
HowellK A, NarsaiR, CarrollA, IvanovaA, LohseM, UsadelB, MillarA H, WhelanJ. Mapping metabolic and transcript temporal switches during germination in rice highlights specific transcription factors and the role of RNA instability in the germination process. Plant Physiol, 2009, 149: 961-980[本文引用:2][JCR: 6.555]
[27]
RajjouL, GallardoK, DebeaujonI, Vand ekerckhoveJ, JobC, JobD. The effect of α-amanitin on the Arabidopsis seed proteome highlights the distinct roles of stored and neosynthesized mRNAs during germination. Plant Physiol, 2004, 134: 1598-1613[本文引用:1][JCR: 6.555]
[28]
SanoN, PermanaH, KumadaR, ShinozakiY, TanabataT, YamadaT, HirasawaT, KanekatsuM. Proteomic analysis of embryonic proteins synthesized from long-lived mRNAs during germination of rice seeds. Plant Cell Physiol, 2012, 53: 687-698[本文引用:6][JCR: 4.134]
GallardoK, JobC, GrootS P C, PuypeM, DemolH, Vand ekerckhoveJ, JobD. Proteomic analysis of Arabidopsis seed germination and priming. Plant Physiol, 2001, 126: 835-848[本文引用:6][JCR: 6.555]
[31]
GallardoK, JobC, GrootS P C, PuypeM, DemolH, Vand ekerckhoveJ, JobD. Proteomics of Arabidopsis seed germination. A comparative study of wild-type and gibberellin-deficient seeds. Plant Physiol, 2002, 129: 823-837[本文引用:9][JCR: 6.555]
[32]
JobC, RajjouL, LovignyY, BelghaziM, JobD. Patterns of protein oxidation in Arabidopsis seeds and during germination. Plant Physiol, 2005, 138: 790-802[本文引用:4][JCR: 6.555]
[33]
YangP F, LiX J, WangX Q, ChenH, ChenF, ShenS H. Proteomic analysis of rice (Oryza sativa) seeds during germination. Proteomics, 2007, 7: 3358-3368[本文引用:4][JCR: 4.132]
[34]
HeD L, HanC, YaoJ L, ShenS H, YangP F. Constructing the metabolic and regulatory pathways in germinating rice seeds through proteomic approach. Proteomics, 2011, 11: 2693-2713[本文引用:1][JCR: 4.132]
[35]
HuangH, MøllerI M, SongS Q. Proteomics of desiccation tolerance during development and germination of maize embryos. J Proteomics, 2012, 75: 1247-1262[本文引用:2][JCR: 4.088]
[36]
WangW Q, MøllerI M, SongS Q. Proteomic analysis of embryonic axis of Pisum sativum seeds during germination and identification of proteins associated with loss of desiccation tolerance. J Proteomics, 2012, 77: 68-86[本文引用:9][JCR: 4.088]
[37]
SheoranI S, OlsonD J H, RossA R S, SawhneyV K. Proteome analysis of embryo and endosperm from germinating tomato seeds. Proteomics, 2005, 5: 3752-3764[本文引用:1][JCR: 4.132]
[38]
MaltmanD J, GaddS M, SimonW J, SlabasA R. Differential proteomic analysis of the endoplasmic reticulum from developing and germinating seeds of castor (Ricinus communis) identifies seed protein precursors as significant components of the endoplasmic reticulum. Proteomics, 2007, 7: 1513-1528[本文引用:2][JCR: 4.132]
[39]
YangM F, LiuX J, LiuY, ChenH, ChenF, ShenS H. Proteomic analysis of oil mobilization in seed germination and postgermination development of Jatropha curcas. J Proteome Res, 2009, 8: 1441-1451[本文引用:5][JCR: 5.056]
[40]
Sghaier-HammamiB, ValledorL, DriraN, Jorrin-NovoJ V. Proteomic analysis of the development and germination of date palm (Phoenix dactylifera L. ) zygotic embryos. Proteomics, 2009, 9: 2543-2554[本文引用:2][JCR: 4.132]
[41]
KimS T, KangS Y, WangY, KimS G, HwangD H, KangK Y. Analysis of embryonic proteome modulation by GA and ABA from germination rice seeds. Proteomics, 2008, 8: 3577-3587[本文引用:7][JCR: 4.132]
[42]
KimS T, WangY, KangS Y, KimS G, RakwalR, KimY C, KangK Y. Developing rice embryo proteomics reveals essential role for embryonic proteins in regulation of seed germination. J Proteome Res, 2009, 8: 3598-3605[本文引用:11][JCR: 5.056]
[43]
RavanelS, GakièreG, JobD, DouceR. The specific features of methionine biosynthesis and metabolism in plants. Proc Natl Acad Sci USA, 2008, 95: 7805-7812[本文引用:1][JCR: 9.737]
[44]
TakahashiH, KoprivaS, GiordanoM, SaitoK, HellR. Sulfur assimilation in photosynthetic organisms: molecular functions and regulations of transporters and assimilatory enzymes. Annu Rev Plant Biol, 2011, 62: 157-184[本文引用:1][JCR: 25.962]
[45]
LuT C, MengL B, YangC P, LiuG F, LiuG J, MaW, WangB C. A shotgun phosphoproteomics analysis of embryos in germinated maize seeds. Planta, 2008, 228: 1029-1041[本文引用:4][JCR: 3.347]
[46]
RajjouL, BelghaziM, HuguetR, RobinC, MoreauA, JobC, JobD. Proteomic investigation of the effect of salicylic acid on Arabidopsis seed germination and establishment of early defense mechanisms. Plant Physiol, 2006, 141: 910-923[本文引用:3][JCR: 6.555]
[47]
KuceraB, CohnM A, Leubner-MetzgerG L. Plant hormone interactions during seed dormancy release and germination. Seed Sci Res, 2005, 15: 282-307[本文引用:9][JCR: 1.931]
[48]
ArcE, Galland M, CueffG, GodinB, LounifiI, JobD, RajjouL. Reboot the system thanks to protein post-translational modifications and proteome diversity: how quiescent seeds restart their metabolism to prepare seedling establishment. Proteomics, 2011, 11: 1606-1618[本文引用:4][JCR: 4.132]
[49]
BuchananB B, BalmerY. Redox regulation: a broadening horizon. Annu Rev Plant Biol, 2005, 56: 187-220[本文引用:3][JCR: 25.962]
[50]
AlkhalfiouiF, RenardM, VenselW H, WongJ, TanakaC K, HurkmanW, BuchananB B, MontrichardF. Thioredoxin-linked proteins are reduced during germination of Medicago truncatula seeds. Plant Physiol, 2007, 144: 1559-1579[本文引用:1][JCR: 6.555]
[51]
KrannerI, RoachT, BeckettR P, WhitakerC, MinibayevaF V. Extracellular production of reactive oxygen species during seed germination and early seedling growth in Pisum sativum. J Plant Physiol, 2010, 167: 805-811[本文引用:1][JCR: 2.699]
[52]
BrockA K, WillmannR, KolbD, GrefenL, LajunenH M, BethkeG, LeeJ, NürnbergerT, GustA A. The Arabidopsis mitogen-activated protein kinase phosphatase PP2C5 affects seed germination, stomatal aperture, and abscisic acid-inducible gene expression. Plant Physiol, 2010, 153: 1098-1111[本文引用:2][JCR: 6.555]
[53]
Montoya-GarciaL, Munoz-OcoteroV, AguilarR, Sanchez de JimenezE. Regulation of acidic ribosomal protein expression and phosphorylation in maize. Biochemistry, 2002, 41: 10166-10172[本文引用:1][JCR: 3.377]
[54]
MoreauM, LindermayrC, DurnerJ, KlessigD F. NO synthesis and signaling in plants—where do we stand ?Physiol Plant, 2010, 138: 372-383[本文引用:1][JCR: 6.555]
[55]
LiuY, YeN, LiuR, ChenM, ZhangJ. H2O2 mediates the regulation of ABA catabolism and GA biosynthesis in Arabidopsis seed dormancy and germination. J Exp Bot, 2010, 61: 2979-2990[本文引用:1][JCR: 5.242]
[56]
JasidS, SimontacchiM, PuntaruloS. Exposure to nitric oxide protects against oxidative damage but increases the labile iron pool in sorghum embryonic axes. J Exp Bot, 2008, 59: 3953-5962[本文引用:1][JCR: 5.242]
CorpasF J, del RíoL A, BarrosoJ B. Need of biomarkers of nitrosative stress in plants. Trends Plant Sci, 2007, 12: 436-438[本文引用:1][JCR: 11.808]
[59]
Lozano-JusteJ, Colom-MorenoR, LeónJ. In vivo protein tyrosine nitration in Arabidopsis thaliana. J Exp Bot, 2011, 62: 3501-3517[本文引用:1][JCR: 5.242]
[60]
NambaraE, OkamotoM, TatematsuK, YanoR, SeoM, KamiyaY. Abscisic acid and the control of seed dormancy and germination. Seed Sci Res, 2010, 20: 55-67[本文引用:3][JCR: 1.931]
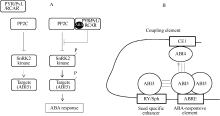
[61]
GraeberK, LinkiesA, MüllerK, WunchovaA, RottA, Leubner-MetzgerG. Cross-species approaches to seed dormancy and germination: conservation and biodiversity of ABA-regulated mechanisms and the Brassicaceae DOG1 genes. Plant Mol Biol, 2010, 73: 67-87[本文引用:3][JCR: 3.518]
[62]
OkamotoM, KuwaharaA, SeoM, KushiroT, AsamiT, HiraiN, KamiyaY, KoshibaT, NambaraE. CYP707A1 and CYP707A2, which encode ABA 8'-hydroxylases, are indispensable for a proper control of seed dormancy and germination in Arabidopsis. Plant Physiol, 2006, 141: 97-107[本文引用:2][JCR: 6.555]
[63]
OkamotoM, TatematsuK, MatsuiA, MorosawaT, IshidaJ, TanakaM, EndoT, MochizukiY, ToyodaT, KamiyaY, ShinozakiK, NambaraE, SekiM. Genome-wide analysis of endogenous abscisic acid-mediated transcription in dry and imbibed seeds of Arabidopsis using tiling arrays. Plant J, 2010, 62: 39-51[本文引用:1][JCR: 6.582]
[64]
Ali-RachediS, BouinotD, WagnerM H, BonnetM, SottaB, GrappinP, JullienM. Changes in endogenous abscisic acid levels during dormancy release and maintenance of mature seeds: studies with the Cape Verde Island s ecotype, the dormant model of Arabidopsis thaliana. Planta, 2004, 219: 479-488[本文引用:3][JCR: 3.347]
[65]
NambaraE, Marion-PollA. Abscisic acid biosynthesis and catabolism. Annu Rev Plant Biol, 2005, 56: 165-185[本文引用:1][JCR: 25.962]
[66]
MillarA A, JacobsenJ V, RossJ J, HelliwellC A, PooleA T, ScofieldG, ReidJ B, GublerF. Seed dormancy and ABA metabolism in Arabidopsis and barley: the role of ABA 8’-hydroxylase. Plant J, 2006, 45: 942-954[本文引用:1][JCR: 6.582]
[67]
LiuY, ShiL, YeN, LiuR, JiaW, ZhangJ. Nitric oxide-induced rapid decrease of abscisic acid concentration is required in breaking seed dormancy in Arabidopsis. New Phytol, 2009, 183: 1030-1042[本文引用:3][JCR: 6.736]
[68]
PrestonJ, TatematsuK, KannoY, HoboT, KimuraM, JikumaruY, YanoR, KamiyaY, NambaraE. Temporal expression patterns of hormone metabolism genes during imbibition of Arabidopsis thaliana seeds: a comparative study on dormant and non-dormant accessions. Plant Cell Physiol, 2009, 50: 1786-1800[本文引用:2][JCR: 4.134]
[69]
MatakiadisT, AlboresiA, JikumaruY, TatematsuK, PichonO, RenouJ P, SottaB, KamiyaY, NambaraE, TroungH N. The Arabidopsis abscisic acid catabolism gene CYP707A2 plays a key role in nitrate control of seed dormancy. Plant Physiol, 2009, 149: 949-960[本文引用:1][JCR: 6.555]
[70]
TohS, ImamuraA, WatanabeA, NakabayashiK, OkamotoM, JikumaruY, HanadaA, AsoY, IshiyamaK, TamuraN, IuchiS, KobayashiM, YamaguchiS, KamiyaY, NambaraE, KawakamiN. High temperature-induced abscisic acid biosynthesis and its role in the inhibition of gibberellin action in Arabidopsis seeds. Plant Physiol, 2008, 146: 1368-1385[本文引用:1][JCR: 6.555]
[71]
GostiF, BeaudoinN, SerizetC, WebbA A R, VartanianN, GiraudatJ. ABI1 protein phosphatase 2C is a negative regulator of abscisic acid signaling. Plant Cell, 1999, 11: 1897-1909[本文引用:1][JCR: 9.251]
[72]
YoshidaT, NishimuraN, KitahataN, KuromoriT, ItoT, AsamiT, ShinozakiK, HirayamaT. ABA-hypersensitive germination3 encodes a protein phosphatase 2C (AtPP2CA) that strongly regulates abscisic acid signaling during germination among Arabidopsis protein phosphatase 2Cs. Plant Physiol, 2006, 140: 115-126[本文引用:1][JCR: 6.555]
[73]
FujiiH, ZhuJ K. Arabidopsis mutant deficient in 3 abscisic acid-activated protein kinases reveals critical roles in growth, reproduction, and stress. Proc Natl Acad Sci USA, 2009, 106: 8380-8385[本文引用:1][JCR: 9.737]
[74]
NakashimaK, FujitaY, KanamoriN, KatagiriT, UmezawaT, KidokoraS, MaruyamaK, YoshidaT, IshiyamaK, KobayashiM, ShinozakiK, Yamaguchi-ShinozakiK. Three Arabidopsis SnRK2 protein kinases, SRK2D/SnRK2. 2, SRK2E/SnRK2. 6/OST1 and SRK2I/SnRK2. 3, involved in ABA signaling are essential for the control of seed development and dormancy. Plant Cell Physiol, 2009, 50: 1345-1363[本文引用:2][JCR: 4.134]
[75]
FujiiH, ChinnusamyV, RodriguesA, RubioS, AntoniR, ParkS Y, CutlerS R, SheenJ, RodriguezP L, ZhuJ K. In vitro reconstitution of an abscisic acid signalling pathway. Nature, 2009, 462: 660-664[本文引用:1][JCR: 38.597]
[76]
UmezawaT, SugiyamaN, MizoguchiM, HayashiS, MyougaF, Yamaguchi-ShinozakiK, IshihamaY, HirayamaT, ShinozakiK. Type 2C protein phosphatases directly regulate abscisic acid-activated protein kinases in Arabidopsis. Proc Natl Acad Sci USA, 2009, 106: 17588-17593[本文引用:1][JCR: 9.737]
[77]
McCourtP, CreelmanR. The ABA receptors—we report you decide. Curr Opin Plant Biol, 2008, 11: 474-478[本文引用:1][JCR: 8.455]
[78]
MaY, SzostkiewiczI, KorteA, MoesD, YangY, ChristmannA, GrillE. Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science, 2009, 324: 1064-1068[本文引用:2]
[79]
Pand eyS, NelsonD C, AssmannS M. Two novel GPCR-type G proteins are abscisic acid receptors in Arabidopsis. Cell, 2009, 136: 136-148[本文引用:1][JCR: 31.957]
[80]
ParkS Y, FungP, NishimuraN, JensenD R, FujiiH, ZhaoY, LumbaS, SantiagoJ, RodriguesA, ChowT F, AlfredS E, BonettaD, FinkelsteinR, ProvartN J, DesveauxD, RodriguezP L, McCourtP, ZhuJ K, SchroederJ I, VolkmanB F, CutlerS R. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science, 2009, 324: 1068-1071[本文引用:3]
[81]
HoldsworthM J, BentsinkL, SoppeW J J. Molecular networks regulating Arabidopsis seed maturation, after-ripening, dormancy and germination. New Phytol, 2008, 179: 33-54[本文引用:3][JCR: 6.736]
[82]
Lopez-MolinaL, Mongrand S, ChuaN H. A postgermination developmental arrest checkpoint is mediated by abscisic acid and requires the ABI5 transcription factor in Arabidopsis. Proc Natl Acad Sci USA, 2001, 98: 4782-4787[本文引用:1][JCR: 9.737]
[83]
OgawaM, HanadaA, YamauchiY, KuwaharaA, KamiyaY, YamaguchiS. Gibberellin biosynthesis and response during Arabidopsis seed germination. Plant Cell, 2003, 15: 1591-1604[本文引用:2][JCR: 9.251]
[84]
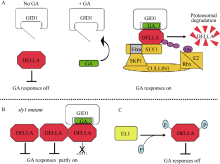
HauvermaleA L, AriizumiT, SteberC M. Gibberellin signalling: a thene and variations on DELLA repression. Plant Physiol, 2012, 160: 83-92[本文引用:6][JCR: 6.555]
[85]
SakamotoT, MiuraK, ItohH, TatsumiT, Ueguchi-TanakaM, IshiyamaK, KobayashiM, AgrawalG K, TakedaS, AbeK, MiyaoA, HirochikaH, KitanoH, AshikariM, MatsuokaM. An overview of gibberellin metabolism enzyme genes and their related mutants in rice. Plant Physiol, 2004, 134: 1642-1653[本文引用:1][JCR: 6.555]
[86]
ApplefordN E J, WilkinsonM D, MaQ, EvansD J, StoneM C, PearceS P, PowersS J, ThomasS G, JonesH D, PhillipsA L, HeddenP, LentonJ. Decreased shoot stature and grain α-amylase activity following ectopic expression of a gibberellin 2-oxidase gene in transgenic wheat. J Exp Bot, 2007, 58: 3213-3226[本文引用:1][JCR: 5.242]
[87]
Ueguchi-TanakaM, AshikariM, NakajimaM, ItohH, KatohE, KobayashiM, ChowT Y, HsingY I C, KitanoH, YamaguchiI, MatsuokaM. GIBBERELLIN INSENSITIVE DWARF1 encodes a soluble receptor for gibberellin. Nature, 2005, 437: 693-698[本文引用:1][JCR: 38.597]
[88]
Chand lerP M, HardingC A, AshtonA R, MulcairM D, DixonN E, Mand erL N. Characterization of gibberellin receptor mutants of barley (Hordeum vulgare L. ). Mol Plant, 2008, 1: 285-294[本文引用:1][JCR: 6.126][CJCR: 0.9487]
WilligeB C, GhoshS, NillC, ZourelidouM, DohmannE M N, MaierA, SchwechheimerC. The DELLA domain of GA INSENSITIVE mediates the interaction with the GA INSENSITIVE DWARF1A gibberellin receptor of Arabidopsis. Plant Cell, 2007, 19: 1209-1220[本文引用:4][JCR: 9.251]
[91]
ItohH, Ueguchi-TanakaM, SatoY, AshikariM, MatsuokaM. The gibberellin signaling pathway is regulated by the appearance and disappearance of SLENDER RICE1 in nuclei. Plant Cell, 2002, 14: 57-70[本文引用:1][JCR: 9.251]
[92]
AriizumiT, LawrenceP K, SteberC M. The role of two F-box proteins, SLEEPY1 and SNEEZY, in Arabidopsis gibberellin signaling. Plant Physiol, 2011, 155: 765-775[本文引用:3][JCR: 6.555]
[93]
WangF, DengX W. Plant ubiquitin-proteasome pathway and its role in gibberellin signaling. Cell Res, 2011, 21: 1286-1294[本文引用:1][JCR: 10.526][CJCR: 1.1032]
[94]
SmalleJ, VierstraR D. The ubiquitin 26S proteasome proteolytic pathway. Annu Rev Plant Biol, 2004, 55: 555-590[本文引用:2][JCR: 25.962]
[95]
AriizumiT, SteberC M. Seed germination of GA-insensitive sleepy1 mutants does not require RGL2 protein disappearance in Arabidopsis. Plant Cell, 2007, 19: 791-804[本文引用:2][JCR: 9.251]
[96]
ZhangZ L, OgawaM, FleetC M, ZentellaR, HuJ, HeoJ O, LimJ, KamiyaY, YamaguchiS, SunT P. SCARECROW-LIKE 3 promotes gibberellin signaling by antagonizing master growth repressor DELLA in Arabidopsis. Proc Natl Acad Sci USA, 2011, 108: 2160-2165[本文引用:1][JCR: 9.737]
[97]
HussainA, CaoD, ChengH, WenZ, PengJ. Identification of the conserved serine/threonine residues important for gibberellin sensitivity of Arabidopsis RGL2 protein. Plant J, 2005, 44: 88-99[本文引用:2][JCR: 6.582]
[98]
HiranoK, AsanoK, TsujiH, KawamuraM, MoriH, KitanoH, Ueguchi-TanakaM, MatsuokaM. Characterization of the molecular mechanism underlying gibberellin perception complex formation in rice. Plant Cell, 2010, 22: 2680-2696[本文引用:1][JCR: 9.251]
[99]
CuiH, BenfeyP N. Interplay between SCARECROW, GA and LIKE HETEROCHROMATIN PROTEIN 1 in ground tissue patterning in the Arabidopsis root. Plant J, 2009, 58: 1016-1027[本文引用:1][JCR: 6.582]
[100]
AriizumiT, SteberC M. Mutations in the F-box gene SNEEZY result in decreased Arabidopsis GA signaling. Plant Signal Behav, 2011, 6: 831-833[本文引用:1]
[101]
Ueguchi-TanakaM, HiranoK, HasegawaY, KitanoH, MatsuokaM. Release of the repressive activity of rice DELLA protein SLR1 by gibberellin does not require SLR1 degradation in the gid2 mutant. Plant Cell, 2008, 20: 2437-2446[本文引用:2][JCR: 9.251]
[102]
AriizumiT, MuraseK, SunT P, SteberC M. Proteolysis independent down-regulation of DELLA repression in Arabidopsis by the gibberellin receptor GIBBERELLIN INSENSITIVE DWARF1. Plant Cell, 2008, 20: 2447-2459[本文引用:2][JCR: 9.251]
[103]
YamamotoY, HiraiT, YamamotoE, KawamuraM, SatoT, KitanoH, MatsuokaM, Ueguchi-TanakaM. A rice gid1 suppressor mutant reveals that gibberellin is not always required for interaction between its receptor, GID1, and DELLA proteins. Plant Cell, 2010, 22: 3589-3602[本文引用:1][JCR: 9.251]
[104]
GriffithsJ, MuraseK, RieuI, ZentellaR, ZhangZ L, PowersS J, GongF, PhillipsA L, HeddenP, SunT P, ThomasS G. Genetic characterization and functional analysis of the GID1 gibberellin receptors in Arabidopsis. Plant Cell, 2006, 18: 3399-3414[本文引用:1][JCR: 9.251]
[105]
FuX, RichardsD E, FleckB, XieD, BurtonN, HarberdN P. The Arabidopsis mutant sleepy1gar2-1 protein promotes plant growth by increasing the affinity of the SCFSLY1 E3 ubiquitin ligase for DELLA protein substrates. Plant Cell, 2004, 16: 1406-1418[本文引用:1][JCR: 9.251]
[106]
DaiC, XueH W. Rice early flowering1, a CKI, phosphorylates DELLA protein SLR1 to negatively regulate gibberellin signalling. EMBO J, 2010, 29: 1916-1927[本文引用:1][JCR: 9.822]
[107]
SwainS M, TsengT S, OlszewskiN E. Altered expression of SPINDLY affects gibberellin response and plant development. Plant Physiol, 2001, 126: 1174-1185[本文引用:1][JCR: 6.555]
[108]
ShimadaA, Ueguchi-TanakaM, SakamotoT, FujiokaS, TakatsutoS, YoshidaS, SazukaT, AshikariM, MatsuokaM. The rice SPINDLY gene functions as a negative regulator of gibberellin signaling by controlling the suppressive function of the DELLA protein, SLR1, and modulating brassinosteroid synthesis. Plant J, 2006, 48: 390-402[本文引用:2][JCR: 6.582]
[109]
FilardoF, RobertsonM, SinghD P, ParishR W, SwainS M. Functional analysis of HvSPY, a negative regulator of GA response, in barley aleurone cells and Arabidopsis. Planta, 2009, 229: 523-537[本文引用:1][JCR: 3.347]
[110]
SilverstoneA L, TsengT S, SwainS M, DillA, JeongS Y, OlszewskiN E, SunT P. Functional analysis of SPINDLY in gibberellin signaling in Arabidopsis. Plant Physiol, 2007, 143: 987-1000[本文引用:1][JCR: 6.555]
[111]
YangS F, HoffmanN E. Ethylene biosynthesis and its regulation in higher plants. Annu Rev Plant Physiol, 1984, 35: 155-189[本文引用:1]
[112]
ChiwochaS D S, CutlerA J, AbramsS R, AmbroseS J, YangJ, RossA R S, KermodeA R. The etr1-2 mutation in Arabidopsis thaliana affects the abscisic acid, auxin, cytokinin and gibberellin metabolic pathways during maintenance of seed dormancy, moist-chilling and germination. Plant J, 2005, 42: 35-48[本文引用:3][JCR: 6.582]
[113]
HermannK, MeinhardJ, DobrevP, LinkiesA, PesekB, HessB, MachackovaI, FischerU, Leubner-MetzgerG. 1-amino-cyclopropane-1-carboxylic acid and abscisic acid during the germination of sugar beet (Beta vulgaris L. ): a comparative study of fruits and seeds. J Exp Bot, 2007, 58: 3047-3060[本文引用:1][JCR: 5.242]
PetruzzelliL, CoraggioI, Leubner-MetzgerG. Ethylene promotes ethylene biosynthesis during pea seed germination by positive feedback regulation of 1-aminocyclopropane-1-carboxylic acid oxidase. Planta, 2000, 211: 144-149[本文引用:1][JCR: 3.347]
[116]
PetruzzelliL, SturaroM, MainieriD, Leubner-MetzgerG. Calcium requirement for ethylene-dependent responses involving 1-aminocyclopropane-1-carboxylic acid oxidase in radicle tissues of germinated pea seeds. Plant Cell Environ, 2003, 26: 661-671[本文引用:1][JCR: 5.135]
[117]
Iglesias-FernándezR, MatillaA. Genes involved in ethylene and gibberellins metabolism are required for endosperm-limited germination of Sisymbrium officinale L. seeds. Planta, 2010, 231: 653-664[本文引用:2][JCR: 3.347]
[118]
MatillaA J, Matilla-VázquezM A. Involvement of ethylene in seed physiology. Plant Sci, 2008, 175: 87-97[本文引用:3][JCR: 2.922]
[119]
LinkiesA, Leubner-MetzgerG. Beyond gibberellins and abscisic acid: how ethylene and jasmonates control seed germination. Plant Cell Rep, 2012, 31: 253-270[本文引用:6][JCR: 2.509]
[120]
MorrisK, LinkiesA, MüllerK, OraczK, WangX, LynnJ R, Leubner-MetzgerG, Finch-SavageW E. Regulation of seed germination in the close Arabidopsis relative Lepidium sativum: a global tissue-specific transcript analysis. Plant Physiol, 2011, 155: 1851-1870[本文引用:1][JCR: 6.555]
[121]
RentzschS, PodzimskaD, VoegeleA, ImbeckM, MüllerK, LinkiesA, Leubner-MetzgerG. Dose- and tissue-specific interaction of monoterpenes with the gibberellin-mediated release of potato tuber bud dormancy, sprout growth and induction of α-amylases and β-amylases. Planta, 2012, 235: 137-151[本文引用:1][JCR: 3.347]
[122]
RohdeA, RuttinkT, HostynV, SterckL, VanDriessche K, BoerjanW. Gene expression during the induction, maintenance, and release of dormancy in apical buds of poplar. J Exp Bot, 2007, 58: 4047-4060[本文引用:1][JCR: 5.242]
[123]
HoldsworthM J, Finch-SavageW E, GrappinP, JobD. Postgenomics dissection of seed dormancy and germination. Trends Plant Sci, 2008b, 13: 7-13[本文引用:2][JCR: 11.808]
[124]
RomanelE A, SchragoC G, CounagoR M, RussoC A, Alves-FerreiraM. Evolution of the B3 DNA binding superfamily: new insights into REM family gene diversification. PloS One, 2009, 4(6): DOI: 10.1371/journal.pone.0005791[本文引用:1][JCR: 3.73]
[125]
SeoM, HanadaA, KuwaharaA, EndoA, OkamotoM, YamauchiY, NorthH, Marion-PollA, SunT P, KoshibaT, KamiyaY, YamaguchiS, NambaraE. Regulation of hormone metabolism in Arabidopsis seeds: phytochrome regulation of abscisic acid metabolism and abscisic acid regulation of gibberellin metabolism. Plant J, 2006, 48: 354-366[本文引用:1][JCR: 6.582]
[126]
DongT T, TongJ H, XiaoL T, ChengH Y, SongS Q. Nitrate, abscisic acid and gibberellin interactions on the thermoinhibition of lettuce seed germination. Plant Growth Regul, 2012, 66: 191-202[本文引用:1][JCR: 1.67]
[127]
CalvoA P, NicolásC, NicolásG, RodríguezD. Evidence of a cross-talk regulation of a GA20-oxidase (FsGA20ox1) by gibberellins and ethylene during the breaking of dormancy in Fagus sylvatica seeds. Physiol Plant, 2004, 120: 623-630[本文引用:1][JCR: 6.555]
[128]
Iglesias-FernándezR, MatillaA. After-ripening alters the gene expression pattern of oxidases involved in the ethylene and gibberellin pathways during early imbibition of Sisymbrium officinale L. seeds. J Exp Bot, 2009, 60: 1645-1661[本文引用:1][JCR: 5.242]
[129]
BarrocoR M, VanPoucke K, BergervoetJ H W, DeVeylder L, GrootS P C, InzeD, EnglerG. The role of the cell cycle machi-nery in resumption of postembryonic development. Plant Physiol, 2005, 137: 127-140[本文引用:1][JCR: 6.555]
[130]
MeiY Q, SongS Q. Early morphological and physiological events occurring during germination of maize seeds. Agric Sci China, 2008, 7: 950-957[本文引用:1][JCR: 0.527][CJCR: 0.4431]
[131]
Gimeno-GillesC, LelievreE, ViauL, Malik-GhulamM, RicoultC, NiebelA, LeducN, LimamiA M. ABA-mediated inhibition of germination is related to the inhibition of genes encoding cell-wall biosynthetic and architecture-modifying enzymes and structural proteins in Medicago truncatula embryo axis. Mol Plant, 2009, 2: 108-119[本文引用:1][JCR: 6.126][CJCR: 0.9487]
[132]
SliwinskaE, BasselG W, BewleyJ D. Germination of Arabidopsis thaliana seeds is not completed as a result of elongation of the radicle but of the adjacent transition zone and lower hypocotyls. J Exp Bot, 2009, 60: 3587-3594[本文引用:2][JCR: 5.242]
[133]
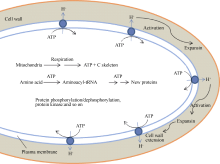
SchopferP. Biomechanics of plant growth. Am J Bot, 2006, 93: 1415-1425[本文引用:1][JCR: 2.586]
[134]
BethkeP C, LibourelI G L, AoyamaN, ChungY Y, StillD W, JonesR L. The Arabidopsis aleurone layer responds to nitric oxide, gibberellin, and abscisic acid and is sufficient and necessary for seed dormancy. Plant Physiol, 2007, 143: 1173-1188[本文引用:1][JCR: 6.555]
[135]
ChenF, BradfordK J. Expression of an expansin is associated with endosperm weakening during tomato seed germination. Plant Physiol, 2000, 124: 1265-1274[本文引用:2][JCR: 6.555]
[136]
ChenF, DahalP, BradfordK J. Two tomato expansin genes show divergent expression and localization in embryos during seed development and germination. Plant Physiol, 2002, 127: 928-936[本文引用:1][JCR: 6.555]
 , 黎妮
, 黎妮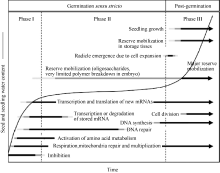



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}