 , 陈奕涵, 何义亮
, 陈奕涵, 何义亮
上海交通大学环境科学与工程学院, 上海 200240
收稿日期: 2017-07-03; 修回日期: 2017-09-08; 录用日期: 2017-09-26
基金项目: 上海交通大学-新加坡国立大学重大国际合作项目(CREATE项目);国家水体污染控制与治理科技重大专项(No.2014ZX07206001)
作者简介: 江月(1993-), 女, E-mail: Jiangyue0524@sjtu.edu.cn
通讯作者(责任作者): 何义亮, E-mail: ylhe@sjtu.edu.cn
摘要: 抗生素抗性基因是水环境中的新型污染物.为了探究夏季时期长江下游某水源型水库抗生素抗性基因(Antibiotic Resistance Genes,ARGs)污染赋存特征,分别采集了水体和沉积物样本并使用高通量荧光定量PCR和实时荧光定量PCR进行研究.结果表明,长江下游水体中检测出104种ARGs,水库水体中检测出118种ARGs,沉积物中检测出124种ARGs.β-内酰胺类、多重抗药类、其他类共54种ARGs为水库中占主导的抗性基因.抗性基因绝对丰度在水体中呈现从长江下游到库内富集的趋势,这表明水环境中的ARGs在水库中富集;相对丰度特征呈现长江下游水介质与库内水介质聚类,水介质与沉积物介质聚类.水体样本的整合子(CintⅠ-1(class 1)、intⅠ-1(clinical))与其ARGs总量之间存在显著的相关性(r=0.750,p < 0.05;r=0.971,p < 0.05),说明整合子对ARGs转移和富集具有重要作用.
关键词:长江下游某水源型水库抗生素抗性基因高通量定量PCR实时荧光定量PCR
Study on antibiotic resistance genes contamination in a reservoir located at Yangtze River downstream
JIANG Yue
, CHEN Yihan, HE Yiliang School of Environmental Science and Engineering, Shanghai Jiao Tong University, Shanghai 200240
Received 3 July 2017; received in revised from 8 September 2017; accepted 26 September 2017
Supported by the Campus for Research Excellence and Technological Enterprise(CREATE) Program and the National Major Science and Technology Program for Water Pollution Control and Treatment(No. 2014ZX07206001)
Biography: JIANG Yue(1993—), female, E-mail: Jiangyue0524@sjtu.edu.cn
*Corresponding author: HE Yiliang, E-mail: ylhe@sjtu.edu.cn
Abstract: Antibiotic resistance genes (ARGs) are known as emerging contaminations in the aquatic environment. In order to explore the levels of ARGs contaminations in a reservoir downstream of the Yangtze River during summer, water and sediment samples were collected and analysed using high throughput quantitative PCR (HT-qPCR) and real-time quantitative PCR (RT-qPCR). The results show that 104, 118 and 124 ARG targets were detected positive in the lower reaches of the Yangtze River, reservoir and sediment respectively. β-lactamase genes, multidrug resistance genes and 54 other kinds of ARGs were identified to be the dominant genes in the reservoir. The absolute abundances of the ARGs in the reservoir were found to be higher than that in the lower reaches of Yangtze River, which indicates an accumulation of ARGs in the reservoir. A heatmap with cluster analysis on the relative ARGs abundance shows clustering between the lower reaches of the river and the reservoir, and between the water columns and the sediment. Relative abundances of integrons (CintⅠ-1 (class 1), intⅠ-1 (clinical)) are significantly correlated to the levels of total ARGs (r=0.750, p < 0.05;r=0.971, p < 0.05), which elucidates that integrons can play an important role in ARGs transfer and accumulation.
Key words: reservoir in downstream of the Yangtze Riverantibiotic resistance geneshigh throughput quantitative PCRreal-time quantitative PCR
1 引言(Introduction)抗生素被长期用于预防和治疗人类和动物疾病, 并在动物养殖业当中广泛使用, 主要作为生长促进剂以亚治疗剂量加入动物饲料中(周启星等, 2007).大多数抗生素只有15%可以被吸收利用, 剩余的通过尿液和粪便排入自然生态系统(Zhang et al., 2011).有研究表明, 在2013年, 中国大约有53800 t抗生素释放到河流和水道中(Zhang et al., 2015).进入到环境中的抗生素除了直接对环境造成污染外, 还可能诱导环境中耐药性细菌和抗生素抗性基因(Antibiotic Resistance Genes, ARGs)的产生(Kim et al., 2004).抗生素的广泛生产和滥用促进了ARGs在环境中的富集和传播, 2010年发现的超级细菌, 其携带耐药基因NDM-1编码的β-内酰胺酶有消解大多数抗生素的能力(Zhang et al., 2011).
国内外****已对地下水、地表水、污水处理厂废水、饮用水中抗生素和ARGs的含量展开研究.在我国的珠江、黄浦江、黄河、海河流域中都检测到抗生素与ARGs, 例如, Lin等(2015)对长江入海口及沿海沉积物中的ARGs展开研究, 发现sul类、tet类、intⅠ基因在这一区域内普遍存在, 含量从河口到海岸呈现出下降的趋势;Jiang等(2013)以黄浦江流域和流域内饮用水水源地为研究目标, 发现水源地ARGs的含量高于黄浦江流域的其他地区;Huerta等(2013)对水源地水库抗生素、抗性基因与微生物群落之间的关系进行了研究.
鉴于国内外现阶段研究目标聚焦在单一水环境, 针对ARGs从河流系统到水库系统赋存特征的研究较少, 且夏季时期藻类含量高, 微生物活跃, 是水库水质的高风险时期.因此, 本研究选择夏季时期长江下游某水源型水库为研究对象, 分别采集水样和沉积物样本并使用高通量定量PCR和实时荧光定量PCR方法, 检测水体和沉积物中的八大类共285种目前已知的主要类型抗生素抗性基因, 并对其赋存分布特征进行分析, 以期为水库ARGs污染控制及水质安全保障提供基础依据.
2 材料与方法(Materials and methods)2.1 样品采集该水源型水库位于长江下游(31°48'N, 121°57'E), 供水规模超过719万m3·d-1.本研究于2016年5月和6月赴水库采样, 围绕整个水库共设置4个采样点(图 1).
图 1(Fig. 1)
 |
| 图 1 水库地理位置示意图 Fig. 1Location of reservoir |
S1样点位于长江下游(水库外), 是水库的入水口, 入水口水流速度和浊度变化明显(5、6月样品分别命名为5-S1、6-S1);S2样点位于水库内下游水闸, 该泵站具有维持水库蓄水量恒定的功能(5、6月样品分别命名为5-S2、6-S2);S3样点位于库内输水泵站, 该泵站将水库内水输送至自来水厂(5、6月样品分别命名为5-S3、6-S3);S4样点位于库内中游位置, 是水库重要的一个巡视位置(5、6月样品分别命名为5-S4、6-S4).在S1~S4样点距离水面0.5 m处采集10 L水样, 立即运回实验室采用0.22 μm滤膜进行过滤并于-80 ℃冰箱保存;在库内中游处用抓泥斗采集表层沉积物, 命名为SE4(5、6月样品分别命名为5-SE4、6-SE4), 采取的沉积物立即用铝箔纸封存并于-80 ℃冰箱保存.
2.2 水样及沉积物DNA提取水样和沉积物的DNA分别用E.Z.N.A.? Water DNA Kit和E.Z.N.A.? Soil DNA Kit (OMEGA, USA)提取, 根据试剂盒的说明书, 按操作步骤进行提取.提取出的DNA用1%琼脂糖凝胶电泳和Thermo Nanodrop2000微量分光光度计(Thermo Scientific, USA)进行检测, DNA样品用QuantiFluor? dsDNA System (Promega Corporation, USA)试剂盒测定双链DNA浓度, 根据测定的样品DNA浓度, 用灭菌的超纯水统一稀释至30 ng·μL-1.
2.3 高通量荧光定量PCR使用SmartChip Real-Time PCR Systems(WaferGen Inc., USA)高通量荧光定量反应平台测定DNA样品的ARGs.该SmartChip纳米孔平台可用于大规模基因表达研究, 每次运行可处理5184个纳米孔反应(Zhu et al., 2013).使用的293对引物在其他研究者的研究中已经被有效验证, 新加入3对引物, 1对用于测定超级细菌抗生素抗性基因blaNDM-1, 1对用于测定intⅠ1整合子基因(class1 intergon), 1对用于测定CintⅠ1临床医学意义上的整合子基因(clinical class1 intergon)(黄福义等, 2016).
每个纳米孔用于PCR扩增反应的总体积为100 nL, 由1×LightCycler 480 SYBR? GreenⅠMaster Mix(Roche, USA)、1 mg·mL-1牛血清蛋白(BSA)、nuclease-free PCR-grade water、500 nmol·L-1上下游引物和5 ng·μL-1 DNA模板组成.使用SmartChip Multisample Nanodispenser (MSND)将PCR混合物释放到芯片的纳米孔中, 随后用SmartChip Cycler进行实时qPCR.每个芯片都有无模板的阴性对照.反应条件:初始95 ℃下预变性10 min;95 ℃变性30 s, 60 ℃退火延伸30 s, 40个循环.仪器程序自动升温进行熔解曲线分析, 导出qPCR反应得到的扩增效率介于1.8~2.2的数据.根据该系统的精确度和灵敏度, 将仪器的检测限设为循环次数CT=31(Wang et al., 2014).
2.4 实时荧光定量PCR使用PikoReal-Time PCR System(Thermo Scientific., USA)进行qPCR定量分析.通过TA克隆, 构建带有pMD? 18-T Vetor (TaKaRa Biotechnology, China)的质粒载体, 将10倍浓度梯度稀释的质粒作为定量的标准品, 用于制作绝对定量的标准曲线.qPCR反应体系如下:12.5 μL SYBR? Premix Ex TaqTM Ⅱ(2×) (TaKaRa Biotechnology, China), 正向、反向引物(10 μmol·L-1)各0.5 μL, DNA模板(10 ng·μL-1)1 μL, 总反应体积为25 μL.qPCR反应条件为:95 ℃下预变性30 s;95 ℃变性5 s, 60 ℃退火30 s, 循环40次;72 ℃延伸3 min.
2.5 数据分析根据Looft等(2012)的研究, 用公式(1)、(2)分别计算基因的相对拷贝数和绝对丰度.
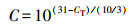 | (1) |
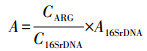 | (2) |
利用Microsoft Excel 2010进行计算分析, 利用SPSS 21.0进行相关性分析, 利用Origin Pro 9.0做图, Circos图利用Circos software (http://circos.ca/)制作, 热图利用MeV制作.
3 结果与讨论(Results and discussion)3.1 水库抗生素抗性基因多样性按照抗性基因对应的抗生素类型将295种ARGs分为九大类抗生素抗性基因(黄福义等, 2016), 分别为磺胺类抗生素(Sulfonamide)抗性基因(7种)、四环素类抗生素(Tetracycline)抗性基因(39种)、氯霉素类抗生素(Chloramphenicol)抗性基因(3种)、β-内酰胺类抗生素(β-Lactamase)抗性基因(54种)、大环内脂类-林肯酰胺类-链阳性菌素B类抗生素(MLSB)抗性基因(45种)、氨基糖苷类抗生素(Aminoglycoside)抗性基因(36种)、万古霉素类抗生素(Vancomycin)抗性基因(32种)、多重抗药类抗生素(Multidrug)抗性基因(52种)和其他类抗生素(Others)抗性基因(27种).以上九大类ARGs均在水体和沉积物中检测到.
如图 2所示, 5-S1样品检出104种ARGs, 5-S2样品检出118种, 5-S3样品检出115种, 5-S4样品检出118种, 5-SE4样品检出122种;6-S1样品检出101种ARGs, 6-S2样品检出116种, 6-S3样品检出114种, 6-S4样品检出117种, 6-SE4样品检出124种.有研究人员对水体和沉积物中的抗性基因污染情况展开了研究, 例如, Xu等(2016)检测了钱塘江流域下游饮用水厂进水、出水和自来水中ARGs的种类, 分别检出168、115、117种ARGs;Wang等(2014)研究了公园沉积物中ARGs的分布情况, 选取北京、石家庄、济南、武汉、昆明、包头和乌鲁木齐地区7个使用再生水灌溉植物的公园, 结果显示, 7个公园共检出147种ARGs.Zhu等(2017)对辽宁、河北、天津、浙江、福建、广东、广西共18个河口沉积物的ARGs开展研究, 各河口检测到的ARGs种类为85~158种, 平均为118.与这些研究结果相比, 长江下游及其蓄水水库的抗性基因污染程度处于中等水平.
图 2(Fig. 2)
 |
| 图 2 研究区不同样点检测到的抗生素抗性基因种类数 Fig. 2Number of detected antibiotic resistance genes in different sampling sites |
3.2 抗生素抗性基因的相对丰度ARGs的相对丰度表明, β-内酰胺类、多重抗药类、其他类(包含转座子基因、整合子基因)抗生素抗性基因在样品中的相对丰度较高(图 3).β-内酰胺类ARGs在10个样品中的相对丰度分别为4.58×10-3(5-S1)、7.53×10-2(5-S2)、6.26×10-2(5-S3)、7.74×10-2(5-S4)、5.68×10-2(5-SE4)、3.25×10-2(6-S1)、6.85×10-2(6-S2)、3.55×10-2(6-S3)、5.70×10-2(6-S4)、5.12×10-2(6-SE4).本实验主要研究bla和ampC两类基因, 实际检测到5种ampC基因, 相对丰度为7.48×10-6~7.43×10-4, ampC-06只在沉积物中检出;14种bla基因的相对丰度为1.05×10-6~1.08×10-2, 其中, blaPAO、blaSHV-02只在沉积物中检出, blaVEB只在水体中检出.在澳大利亚的饮用水及医院和城市废水中都检测到ampC, 德国的废水中该ARG检出率为78% (Schwartz et al., 2003).bla通常在动物来源的病原体(气单胞菌、肠杆菌、沙门氏菌)中被检测到, 在地表水中同样被发现(Zhang et al., 2009).β-内酰胺类抗生素具有低毒性的特点, 在人类医疗和农耕养殖业中被广泛使用, 人类使用的抗生素药物(青霉素、头孢菌素)中有2/3都为这一大类, 也是德国兽药用量最大的一类抗生素(Stoll et al., 2012).因此, 该类ARGs的高相对丰度与检出率可能是由于医疗废水、生活污水、养殖废水排放进入环境所导致.
图 3(Fig. 3)
 |
| 图 3 水体和沉积物中ARGs相对丰度(a.5月,b.6月) Fig. 3Relative abundance of ARGs in water and sediment samples |
多重抗药类ARGs的相对丰度较高(4.66×10-3~9.55×10-2), 5-S1处相对丰度为4.66×10-3, 5-S4处为4.04×10-2, 5-SE4处为7.56×10-2, 在水库中的最大富集倍数(基因相对拷贝数的比值)为9.5, 沉积物中为1.87;6月库外相对丰度为3.53×10-2, 最高相对丰度出现在S2处, 为9.55×10-2, 6-SE4处为9.30×10-2, 在水库中的最大富集倍数为2.7, 沉积物中为2.0.这类抗生素在医院和养殖业中被广泛使用, 残余的抗生素会诱导抗性病菌和多重抗药基因产生, 九龙江流域中有70.6%的细菌菌株对抗生素呈现多重耐药性.有研究发现, 废水经过污水处理厂, 多重耐药性水平提高(Wang et al., 2014), 这也可能是多重抗药类ARGs进入水体的一个重要途径.中国抗生素耐药性增长速度很快, 因此, 合理使用抗生素, 避免多重耐药病原体在环境中的传播显得尤为重要.
氯霉素类ARGs相对丰度与其他几类ARGs相对丰度相比处于中等污染水平(8.24×10-3~2.10×10-2).这可能是因为从20世纪50年代起, 该类抗生素就被长时间应用于治疗动物和人类疾病.尽管现如今很多国家都禁止在家禽中使用该类抗生素, 但因为它具有抗生素效力稳定、价格低廉的特点, 在中国水产养殖和畜牧业中仍部分存在非法使用该类抗生素的现象, 因此, 这类抗生素对应的ARGs在环境中仍有存在.
其他类抗生素抗性基因包括转座子基因、整合子基因和喹诺酮类基因, 转座子和整合子可以捕获整合及表达抗性基因.在下游水闸采样点处(S2)相对丰度高达9.24×10-2, 高于S4处相对丰度(2.11×10-2).本研究发现, 8个水体样品测得的CintⅠ-1(class 1)、intⅠ-1(clinical)的相对含量与ARGs总量之间存在显著相关性(分别为r=0.750, p<0.05;r=0.971, p<0.05).在城市表层水中分离的弧菌菌株中发现, CintⅠ-1(class1)和甲氧苄氨嘧啶、氨基糖苷、β-内酰胺形成具有抗性的共轭原件(Zhang et al., 2009).Zhu等(2017)研究发现, ARGs的丰度与整合子、转座子之间分别存在显著相关性(p<0.01).Luo等(2011)研究发现, 海河沉积物中intⅠ和sulⅠ相对含量之间有显著相关性, 并且证明intⅠ参与了环境中sulⅠ的传播.一些研究表明, intⅠ-1可以通过基因盒整合, 表达超过100种ARGs(包括氨基糖苷类类、β-内酰胺类、磺胺类)(Gillings et al., 2008), 这说明CintⅠ-1(class 1)、intⅠ-1(clinical)可能对ARGs的水平传播起到关键作用.
3.3 抗生素抗性基因的分布特征图 4显示了检测到的ARG相对丰度在每个采样点的分布特征.比较发现, 库外ARGs种类与相对丰度均小于库内, 其中, β-内酰胺类ARGs在库中富集了4.75~16.89倍, 多重抗药类ARGs在库中富集了3.61~9.51倍, 其他类ARGs在库中富集了2.18~5.05倍;比较S4与SE4发现, 沉积物的ARGs种类多于水体(SE4>S4), β-内酰胺类ARGs在沉积物中富集了0.73~1.58倍, 沉积物中氯霉素类、多药类和万古霉素类ARGs相对丰度高于水体, 说明ARGs在沉积物中得到富集.mexF、intⅠ-1(clinic)、cphA-01是3种优势抗性基因, 其中, intⅠ-1(clinic)是对ARGs水平迁移有关键作用的整合子基因.5-S1与6-S1聚类, 5-S3与6-S3聚类, 5-S4与6-S4聚类, 5-SE4与6-SE4聚类, 库外采样点(S1)与库内3个点(S2、S3、S4)聚类, 沉积物与水样聚类, 表明水库沉积物的抗性分布格局不同于水库内和水库内的3个水样.
图 4(Fig. 4)
 |
| 图 4 水体和沉积物中ARGs分布格局 Fig. 4Profile of ARGs in water and sediment samples |
有12种ARGs (erm(34)、acrA-02、acrA-03、acrF、adeA、mexA、yceL_mdtH-01、vanC2_vanC3、vanHB、tnpA-05、aac、blaVIM)只在水库内检出, 库内ARGs多样性大于库外.而本课题组前期研究发现, 水库对总氮、总磷等常规指标, 以及药品和个人护理用品类污染物(PPCPs), 如咖啡因、双酚A、抗生素等有削减作用, 并未出现库内ARGs丰度高于库外的现象.出现这种差异的原因可能是ARGs作为一种生物特性的新型污染物, 有别于传统的化学特性的新型污染物, 其生物体间具有的传播机制和独特的环境行为尚需进一步的研究.但已有研究表明, ARGs丰度会随着采样点处流速的增大而减小(Jiang et al., 2013).而本研究中水库上游进水闸(S1)流速(0.2 m·s-1)>库区流速(0.01~0.19 m·s-1)>下游水闸(S2)流速(0.01~0.03 m·s-1)(袁建忠等, 2012), 显示长江下游的水流速度明显高于库内流速.此外, 16S rDNA绝对定量结果显示, 从库外S1到库内S2、S3、S4及SE4的16S rDNA的均值分别为4.79×108、1.13×1010、1.89×1010、4.18×1010、6.56×109 copies·g-1.表明库内及其沉积物中的细菌数量显著高于库外1~2个数量级, 而丰富的生物量有助于ARGs在不同细菌间的转移和积累.因此, 水流速度和细菌数量可能是造成这种现象的主要原因.
10种ARGs只在沉积物中被检出, 分别为vgaA-01、tetM-01、tetPB-01、tetS、bacA-02、catB3、aacC1、ampC-06、blaPAO、blaSHV-02.环境中残留的抗生素可能会诱导ARGs的出现, 且抗生素特性的差异会影响ARGs在环境中的分布.大环内酯类抗生素易被水解且吸附于沉积物中, 喹诺酮类抗生素同样易被光降解和吸附于沉积物中(Luo et al., 2011), 这两种抗生素对应的ARGs在相对丰度与种类上均显示出沉积物大于水体的特征.这说明这类易水解、光解且吸附于沉积物的抗生素对应的ARGs在水体中的相对丰度与种类均小于对应沉积物.以上结果显示, 沉积物介质具有选择性富集大环内酯类和喹诺酮类抗性基因的功能, 也进一步证实了水体介质与沉积物介质之间ARGs分布特征存在差异.
3.4 抗生素抗性基因的绝对丰度每一大类ARGs和每个样品中ARGs绝对丰度和分布特征见图 5.图 5显示, 从6-S1到5-SE4样品, ARGs的总含量依次为1.60×108、1.31×108、1.12×109、2.70×109、2.53×109、3.50×109、5.40×109、6.30×109 copies·L-1和1.80×109、7.86×108 copies·g-1.
图 5(Fig. 5)
 |
| 图 5 每种ARG在5个采样点中的分布 Fig. 5Distribution of each ARG type in five Sampling points |
氨基糖苷类(36种)抗性基因在水样中的绝对丰度总和为4.22×108 copies·L-1, 其中, 6-S1为4.85×106 copies·L-1, 5-S1为6.16×106 copies·L-1, 6-S2为3.62×107 copies·L-1, 5-S2为3.57×107 copies·L-1, 6-S3为5.98×107 copies·L-1, 5-S3为5.99×107 copies·L-1, 6-S4为1.17×108 copies·L-1, 5-S4为1.03×108 copies·L-1;在沉积物中的绝对丰度总和为4.45×107 copies·g-1, 其中, 6-SE4为3.44×107 copies·g-1, 5-SE4为1.02×107 copies·g-1.其他各类ARGs绝对丰度分布度情况如图 5所示.
β-内酰胺类(54种)抗性基因检出率为44%, 水体中浓度为3.44×107~2.49×109 copies·L-1, 沉积物中含量分别为2.36×108 copies·g-1和4.59×108 copies·g-1.其中, blaTEM绝对丰度为6.68×104~1.96×107 copies·L-1, blaCARB、blaSHV、blaSHV-5未被检出.Jiang等(2013)在黄浦江流域检测到blaTEM浓度为1.4×106~2.0×107 copies·L-1, 是最高浓度S4处(1.46×107 copies·L-1)的0.10~1.37倍;Hu等(2008)研究发现, 超过95%的环境样本中检测到blaTEM, 而blaCARB、blaSHV、blaSHV-5的检出率低于1%.该类ARGs在本水库的检出情况与大部分环境样本一致, 且blaTEM含量低于黄浦江流域.
研究结果显示, 磺胺类ARGs和四环素类ARGs绝对丰度不高, 但这两类ARGs在环境中广泛传播(Gillings et al., 2008).sul Ⅰ、sul Ⅱ在本实验中的检出率均为100%, 磺胺类ARGs的含量高于四环素类ARGs的含量, 这一结果与黄浦江流域研究结果一致(沈群辉等, 2012).dfrA1浓度为0~7.95×106 copies·L-1, dfrA12浓度为0~2.83×105 copies·L-1, dfrA12检出率较低, 分布特征与在德国、澳大利亚的研究结果一致(Stoll et al., 2012).这两种ARGs在环境中的分布特征可能是由于sul Ⅰ、sul Ⅱ属于在环境中比较常见的二氢叶酸合成酶基因, 而dfrA1、dfrA12属于在环境中比较少见的二氢叶酸还原酶基因, 这使得sul Ⅰ、sul Ⅱ基因被广泛检出.
11种四环素类(39种)抗性基因被检出, 其中, tetA-02、tetD-02、tetG-01、tetG-02、tetL-02、tetPB-02、tetR-02检出率为100%, 水体中tetA-02浓度为5.98×104~1.20×107 copies·L-1, 低于在太湖中的浓度(107~108 copies·L-1), tetM-01、tetPB-01、tetS只在沉积物中检出.Zhang等(2011)在中国和其他地区发现, 13种四环素类ARGs中, tetG的丰度最高, 其次是tetA、tetC.Hu等(2008)在自然水体中检测了20种四环素类ARGs, 结果只检测到tetA、tetB、tetM 3种.这可能是由于tetA、tetB、tetG由可迁移的载体, 如转座子和较小的质粒携带, 而质粒具有移动性较强的特点, 因此, 可以在耐药细菌之间稳定传播, 而tetO、tetS普遍存在于不可移动的大的共轭质粒或染色体中, 所以这类基因转移和传播的可能性较低(Schwartz et al., 2003).从S1到S4 ARGs的绝对丰度依次递增, 绝对丰度最高的3类ARGs为多重抗药类、β-内酰胺类、其他类ARGs.本水库与其他地区磺胺类和四环素类检出率都较高, 但长江下游和水库内ARGs浓度与其他流域相比较低.
4 结论(Conclusions)1) 夏季时期长江下游某水源型水库平均检测到118种ARGs, 与我国其他地区相比, 检出ARGs种类数处于中等水平.
2) 九大类ARGs中, β-内酰胺类、多药类、氯霉素类和其他类ARGs相对丰度与绝对丰度均高于其他五大类;四环素类ARGs和磺胺类ARGs绝对丰度低于其他几大类ARGs, 但因为sulⅠ、sulⅡ在环境中常见, tetA、tetG由移动性强的质粒携带, 检出率高于其他ARGs.
3) 移动性强的ARGs在环境中检出频率较高, 水体中整合子CintⅠ-1(class 1)、intⅠ-1(clinical)与ARGs总量之间存在显著的正相关性, 具有促进ARGs水平转移的作用, 是ARGs传播的重要途径.
4) 夏季时期ARGs的分布特征受环境介质影响, 使得水库内外, 以及水体与沉积物之间ARGs分布存在差异.低流速环境易于微生物生长, 细菌多样性多, 使得ARGs易在不同细菌间传播;水库内检测到的ARGs种类和丰度均大于长江下游, 这说明缓流水体易成为ARGs高风险区.
参考文献
| Gillings M, Boucher Y, Labbate M, et al. 2008. The evolution of class 1 integrons and the rise of antibiotic resistance[J]. Journal of Bacteriology, 190(14): 5095–5100.DOI:10.1128/JB.00152-08 |
| Huerta B, Marti E, Gros M, et al. 2013. Exploring the links between antibiotic occurrence, antibiotic resistance, and bacterial communities in water supply reservoirs[J]. Science of the Total Environment, 456: 161–170. |
| 黄福义, 李虎, 安新丽, 等. 2016. 城市生活污水和生活垃圾渗滤液抗生素抗性基因污染的比较研究[J]. 环境科学, 2016, 37(10): 3949–3954. |
| Hu J, Shi J, Chang H, et al. 2008. Phenotyping and genotyping of antibiotic-resistant Escherichia coli isolated from a natural river basin[J]. Environmental Science & Technology, 42(9): 3415–3420. |
| Jiang L, Hu X L, Xu T, et al. 2013. Prevalence of antibiotic resistance genes and their relationship with antibiotics in the Huangpu River and the drinking water sources, Shanghai, China[J]. Science of the Total Environment, 4(38): 267–272. |
| Kim S R, Nonaka L, Suzuki S. 2004. Occurrence of tetracycline resistance genes tet (M) and tet (S) in bacteria from marine aquaculture sites[J]. FEMS Microbiology Letters, 237(1): 147–156.DOI:10.1111/fml.2004.237.issue-1 |
| Lin L, Liang X M, Chen X, et al. 2015. Occurrences and distribution of sulfonamide and tetracycline resistance genes in the Yangtze River Estuary and nearby coastal area[J]. Marine Pollution Bulletin, 8(36): 304–310. |
| Luo Y, Xu L, Rysz M, et al. 2011. Occurrence and transport of tetracycline, sulfonamide, quinolone, and macrolide antibiotics in the Haihe River Basin, China[J]. Environmental Science & Technology, 45(5): 1827–1833. |
| Looft T, Johnson T A, Allen H K, et al. 2012. In-feed antibiotic effects on the swine intestinal microbiome[J]. Proceedings of the National Academy of Sciences of the United States of America, 109(5): 1691–1696.DOI:10.1073/pnas.1120238109 |
| 苏建强, 黄福义, 朱永官. 2013. 环境抗生素抗性基因研究进展[J]. 生物多样性, 2013, 21(4): 481–487. |
| Schwartz T, Kohnen W, Jansen B, et al. 2003. Detection of antibiotic-resistant bacteria and their resistance genes in wastewater, surface water, and drinking water biofilms[J]. FEMS Microbiol Ecology, 43(3): 325–335.DOI:10.1111/fem.2003.43.issue-3 |
| Stoll C, Sidhu J P S, Tiehm A, et al. 2012. Prevalence of clinically relevant antibiotic resistance genes in surface water samples collected from Germany and Australia[J]. Environmental Science & Technology, 46(17): 9716–9726. |
| 沈群辉, 冀秀玲, 傅淑珺, 等. 2012. 黄浦江水域抗生素及抗性基因污染初步研究[J]. 环境科学, 2012, 21(10): 1717–1723. |
| Wang F H, Miao M, Su J Q, et al. 2014. High throughput profiling of antibiotic resistance genes in urban park soils with reclaimed water irrigation[J]. Environmental Science & Technology, 48(16): 9079–9085. |
| Xu L, Ouyang W Y, Qian Y Y, et al. 2016. High-throughput profiling of antibiotic resistance genes in drinking water treatment plants and distribution systems[J]. Environmental Pollution, 213: 119–126.DOI:10.1016/j.envpol.2016.02.013 |
| 袁建忠, 李杰. 2012. 青草沙水库库区流态及淤积分布特征研究[J]. 城市道桥与防洪, 2012(9): 181–183. |
| 周启星, 罗义, 王美娥. 2007. 抗生素的环境残留、生态毒性及抗性基因污染[J]. 生态毒理学报, 2007, 2(3): 243–251. |
| Zhang D D, Lin L F, Luo Z X, et al. 2011. .Occurrence of selected antibiotics in Jiulongjiang River in various seasons, South China[J]. Journal of Environmental Monitoring, 13(7): 1953–1960.DOI:10.1039/c0em00765j |
| Zhang Q Q, Ying G G, Pan C G, et al. 2015. Comprehensive evaluation of antibiotics emission and fate in the river basins of China:source analysis, multimedia modeling, and linkage to bacterial resistance[J]. Environ Environmental Science & Technology, 49(11): 6772–6782. |
| Zhu Y G, Johnson T A, Su J Q, et al. 2013. Diverse and abundant antibiotic resistance genes in Chinese swine farms[J]. Proceedings of the National Academy of Sciences of the United States of America, 110(9): 3435–3440.DOI:10.1073/pnas.1222743110 |
| Zhu Y G, Zhao Y, Li Bing, et al. 2017. Continental-scale pollution of estuaries with antibiotic resistance genes[J]. Nature Microbiology, 2: 1–7. |
| Zhang X X, Zhang T, Fang H. 2009. Antibiotic resistance genes in water environment[J]. Applied Microbiology and Biotechnology, 82(3): 397–414.DOI:10.1007/s00253-008-1829-z |
| Zhang X, Zhang T. 2011. Occurrence, abundance, and diversity of tetracycline resistance genes in 15 sewage treatment plants across China and other global locations[J]. Environmental Science & Technology, 45: 2598–2604. |
