Bacterial diversity in surface sediments of Baiyangdian lake and its influencing factors
YU Yan1,,, WANG Yueyue1, FANG Duxian1, REN Jie1, WANG Ying2 1.School of Chemical and Environmental Engineering, China University of Mining and Technology (Beijing), Beijing 100083, China 2.Key Laboratory of Water and Sand Science, Ministry of Education, College of Environmental Sciences, Beijing Normal University, Beijing 100875, China
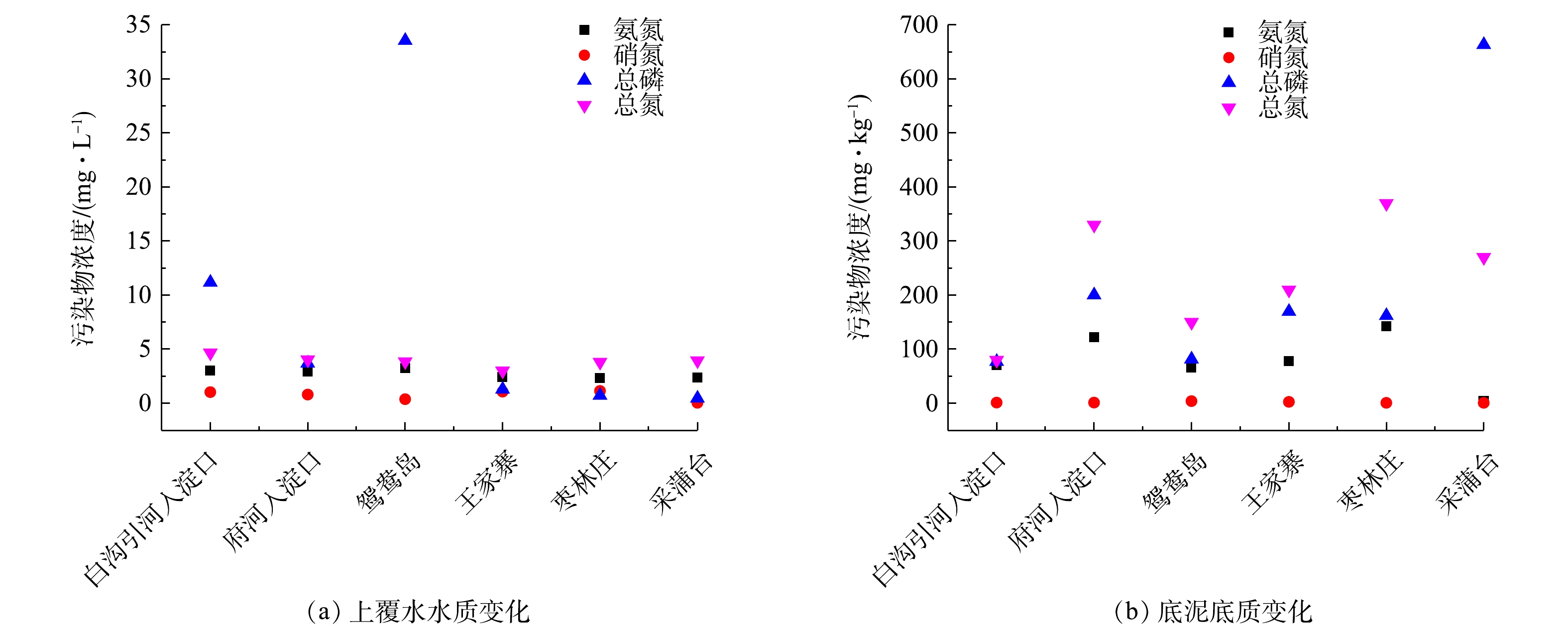
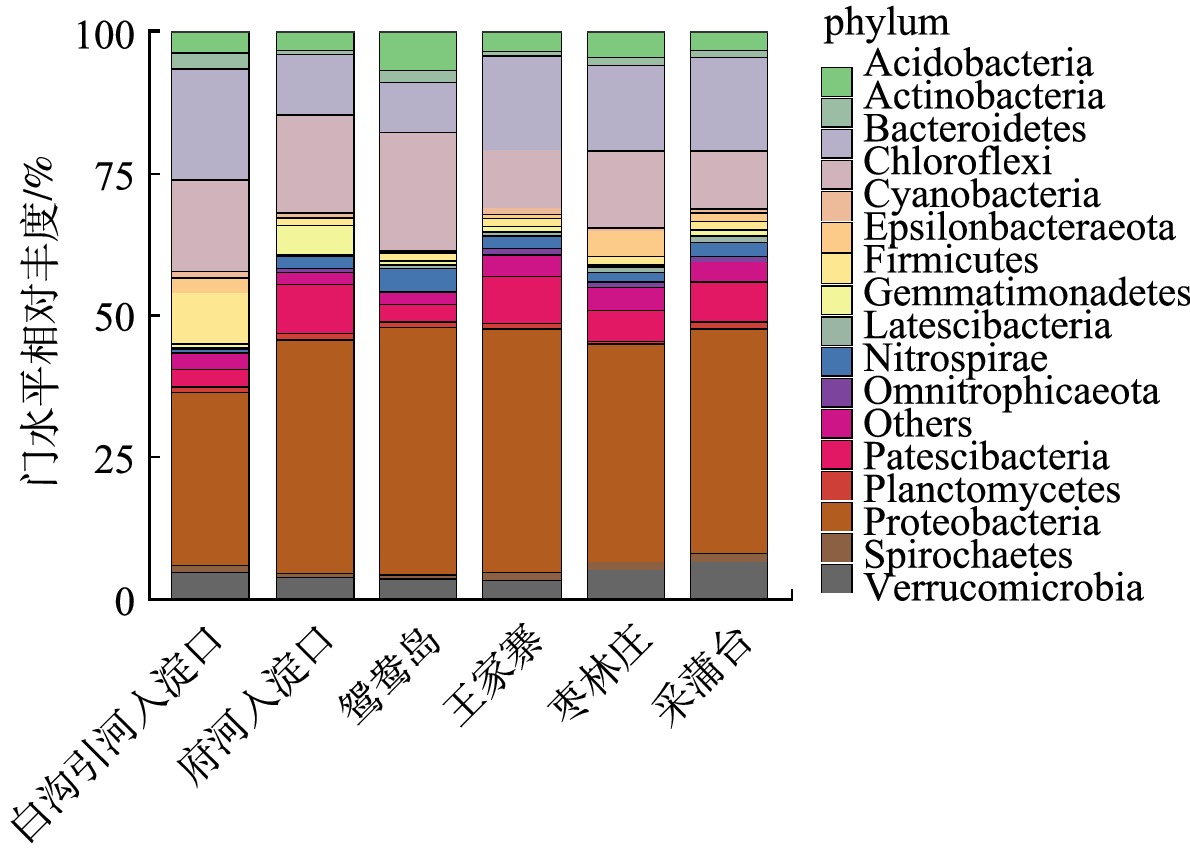
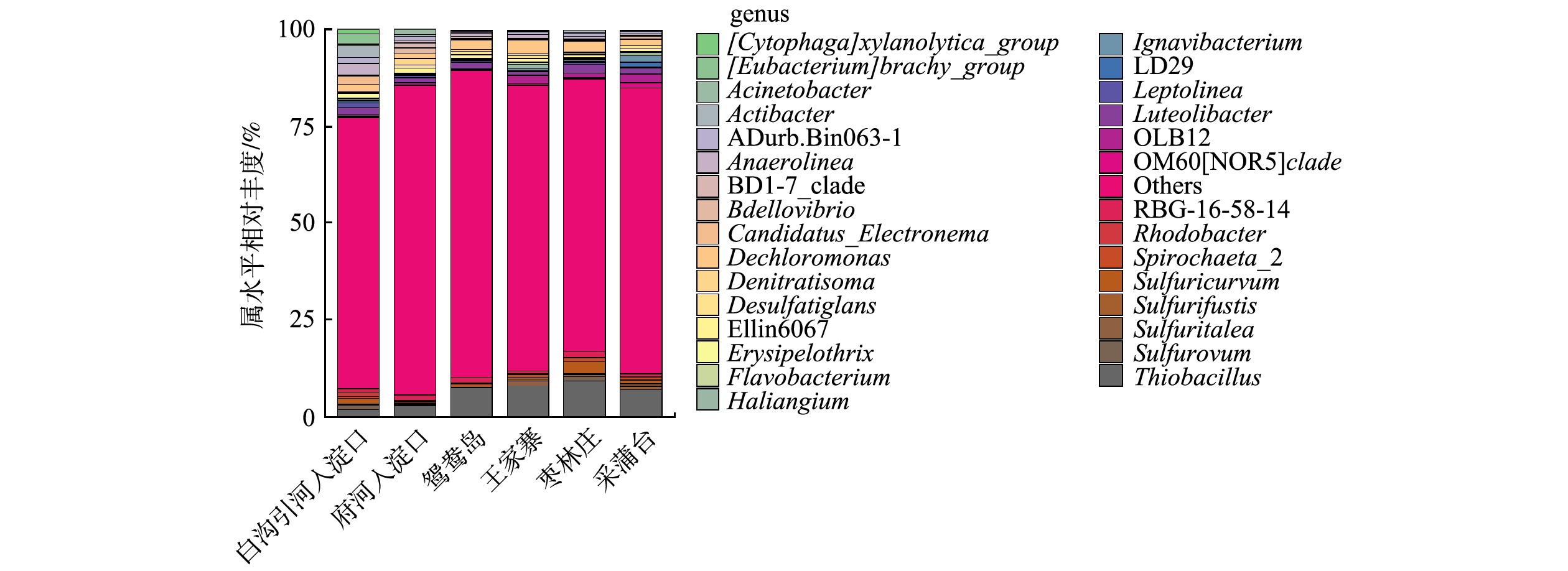
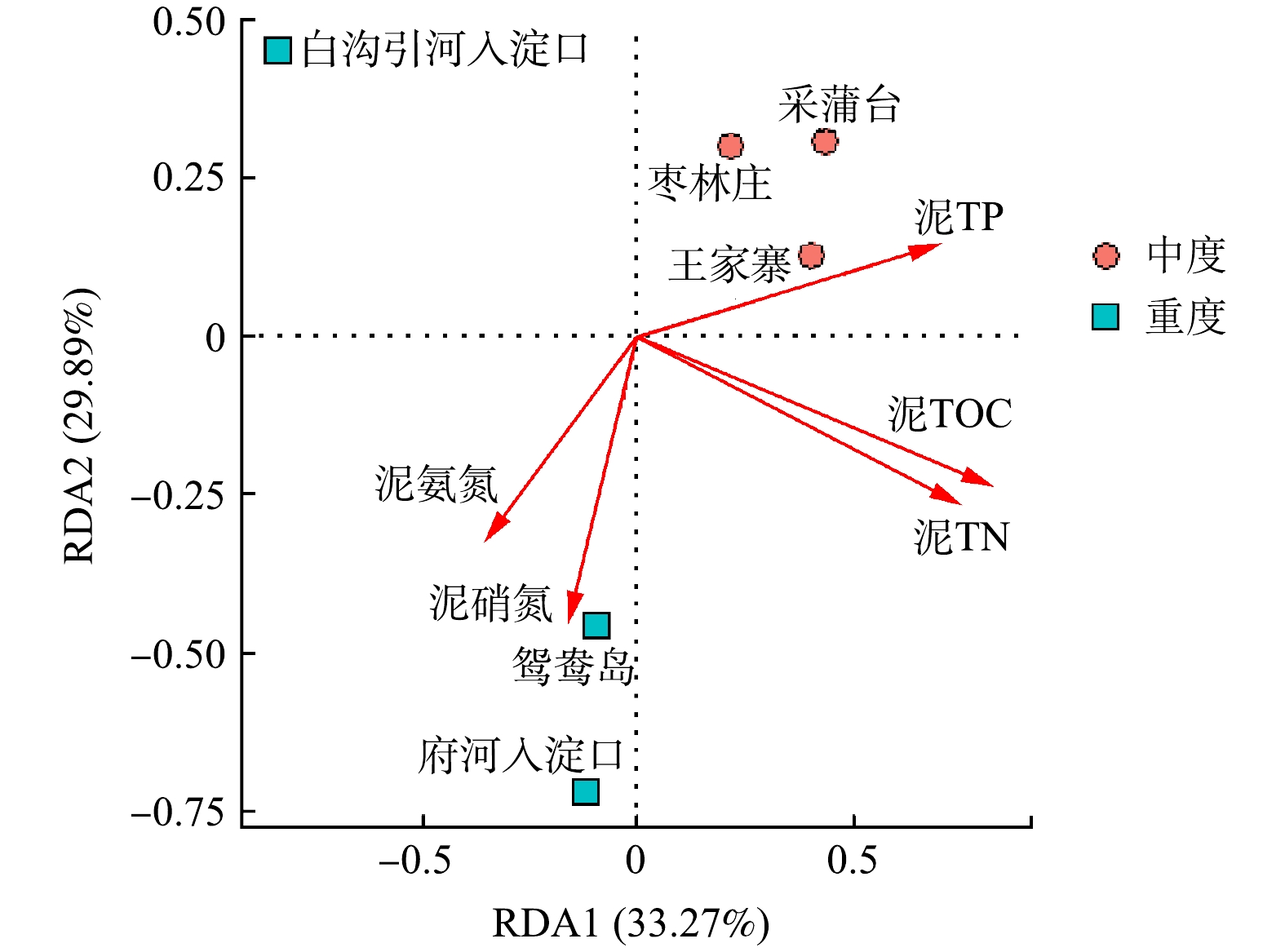
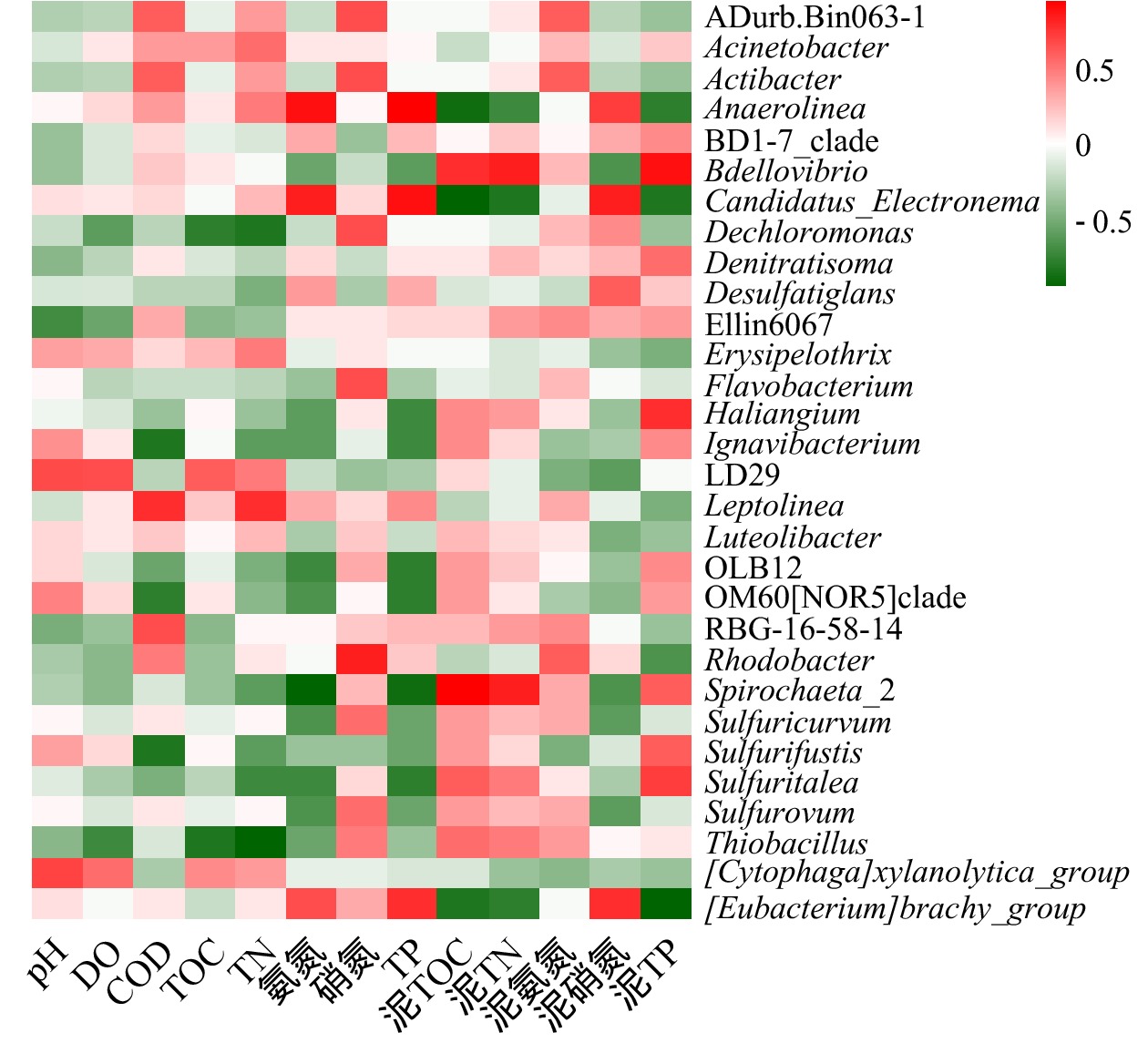
Abstract:The purpose of this study was to explore the differences and driving factors of bacterial community structure in sediments of different eutrophic water bodies in Baiyangdian Lake. The composition and diversity of bacterial community structure under two eutrophication levels in surface sediment, as well as their correlation with environmental factors, were studied through high-throughput sequencing. The results showed that the diversity of bacterial communities decreased with the increase of the eutrophication level. The bacterial community structure was different under the severe and moderate eutrophication levels, and the abundance of Chloroflexi increased significantly under the severe eutrophication level. However, there was no significant difference in the bacterial community structure in Phylum level with different eutrophication in generally. The predominant phylum in the sediments were Proteobacteria (31%~43%), Chloroflexi (10%~21%) and Bacteroidetes (9%~20%), respectively. The heatmap of the correlation between microbial communities and environmental factors revealed that TP and ${\rm{NH}}_4^ + $-N in surface water, TP and ${\rm{NO}}_3^ - $-N in sediments were the main influencing factors of bacteria community diversity in the Baiyangdian Lake. It can be concluded that the diversity of bacteria in sediment was dissimilar in different eutrophic water, and the phosphorus content was identified as the limiting factor of bacterial community structure in Baiyangdian Lake. Key words:baiyangdian Lake/ eutrophication bodies/ sediments/ bacterial community structure/ environmental factors.
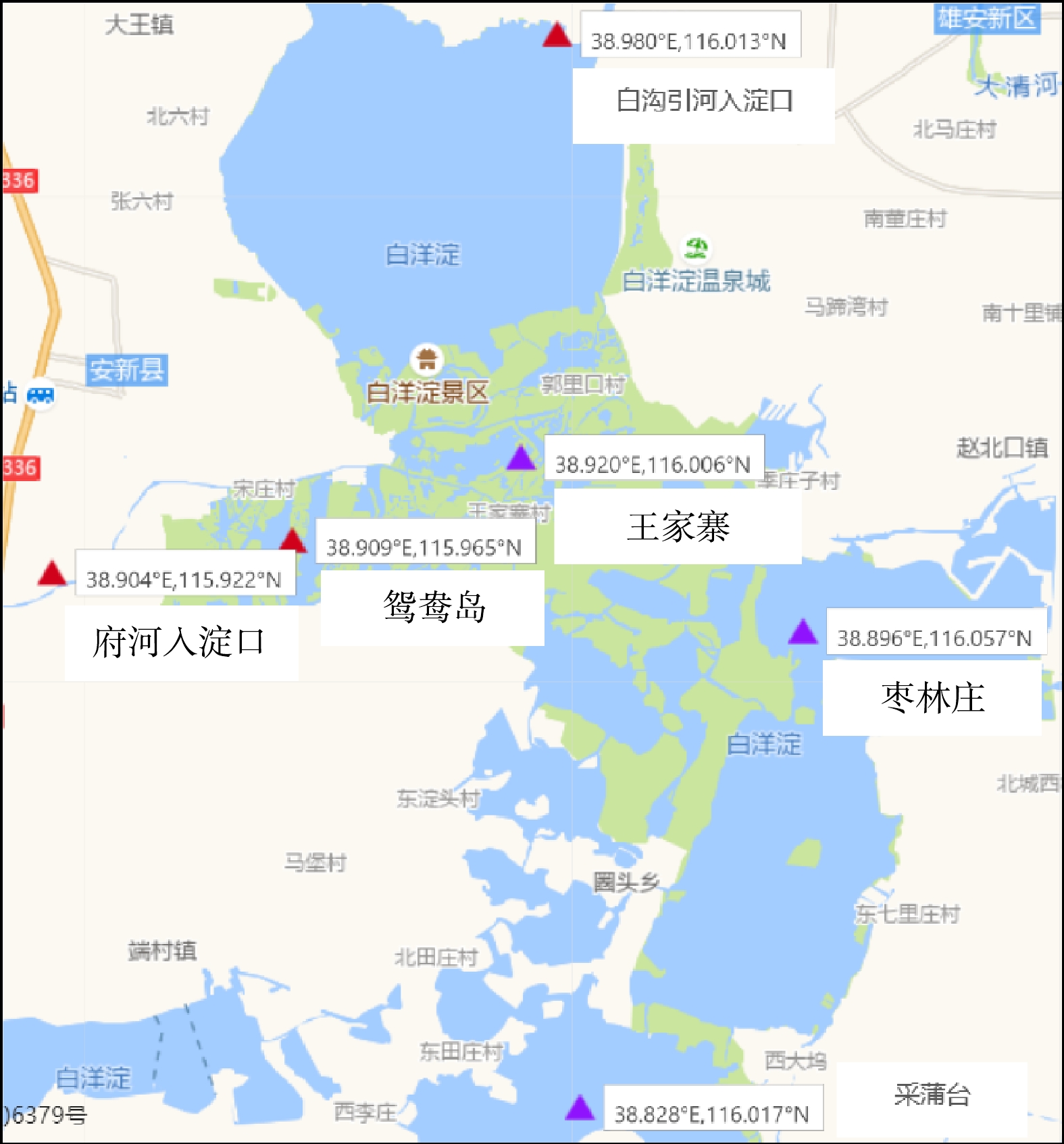
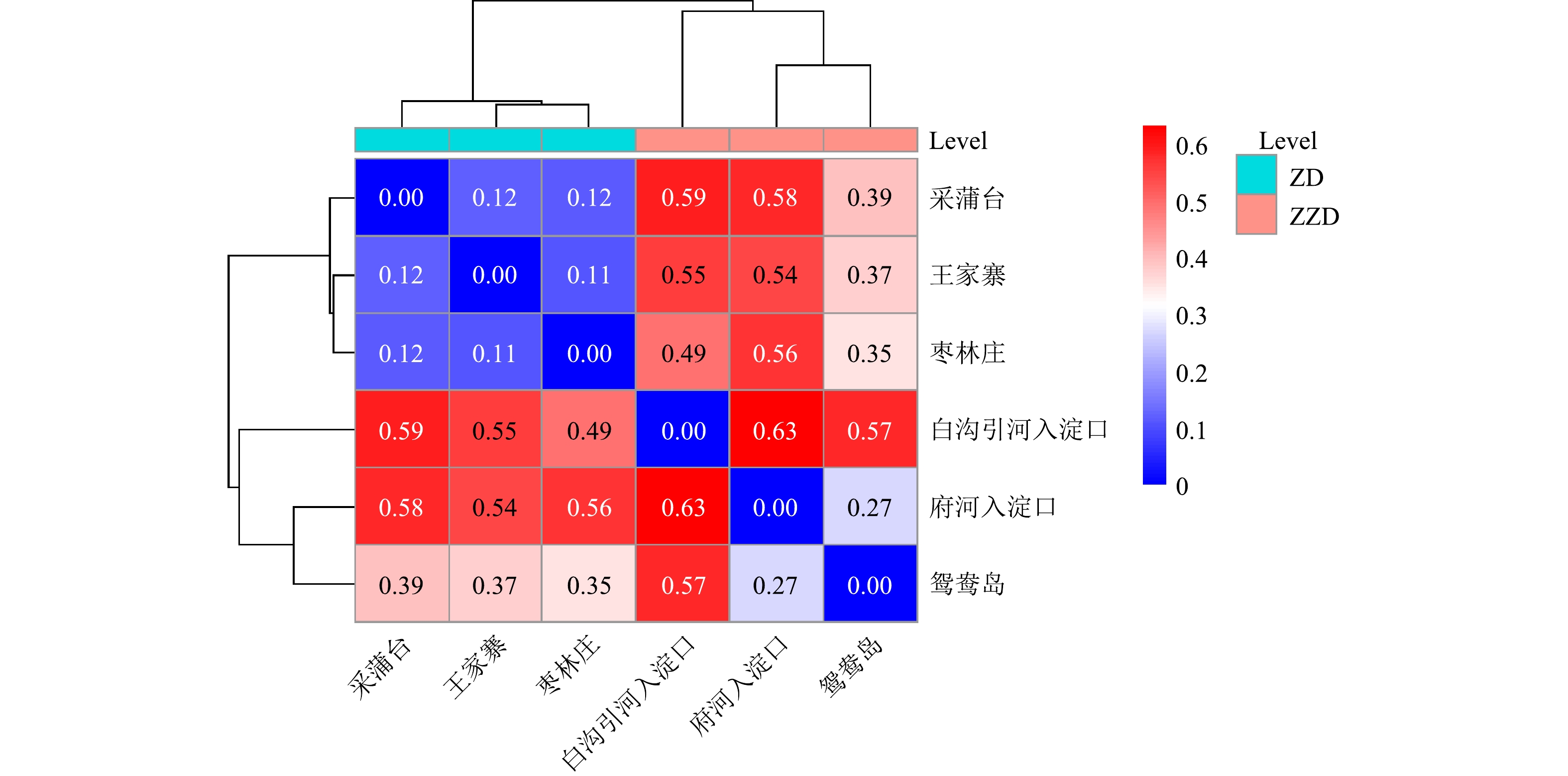
图1采样点分布 Figure1.Sample distributions of different eutrophication levels
图7细菌群落与上覆水、沉积物理化指标相关性分析热图 Figure7.Heatmap of correlation analysis between bacterial communities and physical-chemical indexes of overlying water and sediments
SHENG P, YU Y Z, ZHANG G H, et al. Bacterial diversity and distribution in seven different estuarine sediments of Poyang Lake, China[J]. Springer Berlin Heidelberg, 2016, 75(6): 425-436.
[2]
HUANG R, ZHAO D Y, JIANG C L, et al. Heterogeneity of bacterial community compositions in surface sediments of three lake zones in Lake Taihu[J]. Springer India, 2015, 85(2): 34-42.
JIN X, MA Y, KONG Z Y, et al. The variation of sediment bacterial community in response to anthropogenic disturbances of Poyang Lake, China[J]. Wetlands: Official Scholarly Journal of the Society of Wetland Scientists, 2019, 39(1): 63-73.
[7]
WAN Y, BAI Y, HE J, et al. Temporal and spatial variations of aquatic environmental characteristics and sediment bacterial community in five regions of Lake Taihu[J]. Aquatic Ecology, 2017, 51(3): 35-45.
DAI Y, YANG Y Y, WU Z, et al. Spatiotemporal variation of planktonic and sediment bacterial assemblages in two plateau freshwater lakes at different trophic status[J]. Applied Microbiology and Biotechology, 2016, 100(9): 4161-4175. doi: 10.1007/s00253-015-7253-2
[21]
YANG J, JIANG H, WU G, et al. Distinct factors shape aquatic and sedimentary microbial community structures in the lakes of western China[J]. Frontiers Microbiology, 2016, 42(7): 1772-1782.
SCHREIER H J, MIRZOYAN N, SAITO K. Microbial diversity of biological Filters in recirculating aquaculture systems[J]. Current Opinion in Biotechnology, 2010, 21(3): 318-325. doi: 10.1016/j.copbio.2010.03.011
[24]
ZHANG J X, YANG Y Y, ZHAO L, et al. Distribution of sediment bactetial and archaeal communities in plateau freshwater lakes[J]. Applied Microbiology and Biotechnology, 2015, 99(7): 3291-3302. doi: 10.1007/s00253-014-6262-x
BOWMAN J P, MCCUAIG R D. Biodiversity, community structural shifts, and biogeography of prokaryotes within antarctic continental shelf sediment[J]. Applied and Environmental Microbiology, 2003, 69(5): 2463-2483. doi: 10.1128/AEM.69.5.2463-2483.2003
[27]
LEE S H, KA J O, CHO J C. Members of the phylum Acidobacteria are dominant and metabolically active in rhizosphere soil[J]. FEMS Microbiology Letters, 2008, 285(2): 263-269. doi: 10.1111/j.1574-6968.2008.01232.x
[28]
ZENG Y H, BAUMBACH J, BARBOSA E G V, et al. Metagenomic evidence for the presence of phototrophic Gemmatimonadetes bacteria in diverse environments[J]. Environmental Microbiology Reports, 2016, 48(3): 139-149.
[29]
GOFFREDI S K, ORPHAN V J. Bacterial community shifts in taxa and diversity in response to localized organic loading in the deep sea[J]. Environmental Microbiology, 2010, 34(12): 344-363.
[30]
MARTINS G, TERADA A, RIBEIRO D C, et al. Structure and activity of lacustrine sediment bacteria involved in nutrient and iron cycles[J]. FEMS Microbiology Ecology, 2011, 77(3): 666-679. doi: 10.1111/j.1574-6941.2011.01145.x
1.School of Chemical and Environmental Engineering, China University of Mining and Technology (Beijing), Beijing 100083, China 2.Key Laboratory of Water and Sand Science, Ministry of Education, College of Environmental Sciences, Beijing Normal University, Beijing 100875, China Received Date: 2020-04-17 Accepted Date: 2020-10-15 Available Online: 2021-03-24 Keywords:baiyangdian Lake/ eutrophication bodies/ sediments/ bacterial community structure/ environmental factors Abstract:The purpose of this study was to explore the differences and driving factors of bacterial community structure in sediments of different eutrophic water bodies in Baiyangdian Lake. The composition and diversity of bacterial community structure under two eutrophication levels in surface sediment, as well as their correlation with environmental factors, were studied through high-throughput sequencing. The results showed that the diversity of bacterial communities decreased with the increase of the eutrophication level. The bacterial community structure was different under the severe and moderate eutrophication levels, and the abundance of Chloroflexi increased significantly under the severe eutrophication level. However, there was no significant difference in the bacterial community structure in Phylum level with different eutrophication in generally. The predominant phylum in the sediments were Proteobacteria (31%~43%), Chloroflexi (10%~21%) and Bacteroidetes (9%~20%), respectively. The heatmap of the correlation between microbial communities and environmental factors revealed that TP and ${\rm{NH}}_4^ + $-N in surface water, TP and ${\rm{NO}}_3^ - $-N in sediments were the main influencing factors of bacteria community diversity in the Baiyangdian Lake. It can be concluded that the diversity of bacteria in sediment was dissimilar in different eutrophic water, and the phosphorus content was identified as the limiting factor of bacterial community structure in Baiyangdian Lake.




 下载:
下载:  点击查看大图
点击查看大图