 ,1,2, 白炜鹏1,2,3, 张立民1,2,3
,1,2, 白炜鹏1,2,3, 张立民1,2,3The first description of Rhinocerotidae (Perissodactyla, Mammalia) from Xinyaozi Ravine in Shanxi, North China
DONG Wei,1,2, BAI Wei-Peng1,2,3, ZHANG Li-Min1,2,3通讯作者: *dongwei@ivpp.ac.cn
收稿日期:2021-03-26
| 基金资助: |
Corresponding authors: *dongwei@ivpp.ac.cn
Received:2021-03-26

摘要
20世纪80年代在桑干河盆地一带考察泥河湾层时,在山西省天镇县南高崖乡的辛窑子沟一带发现了很多哺乳动物化石地点并出土了大量哺乳动物化石。最近对其中犀科化石的研究表明在辛窑子沟一带的早更新世地层中产出的犀类化石至少有两个种,裴氏板齿犀(Elasmotherium peii )和泥河湾披毛犀(Coelodonta nihowanensis ), 可能还存在过第三个种。由于标本保存状况不理想,暂时鉴定为梅氏犀相似种(Stephanorhinus cf. S. kirchbergensis )。虽然后者的形态大小与梅氏犀最接近,但和泥河湾披毛犀也有相似之处,因此也有可能是这两个种之一的种内变异类型。同样,产于下沙沟地点被德日进和皮维托鉴定为有疑问的中国犀(Rhinoceros sinensis (?))也可能是梅氏犀或泥河湾披毛犀的种内变异类型。迄今为止在广义泥河湾盆地发现的早更新世犀类有两个确定的种和两个不确定的种。出土裴氏板齿犀的地点为下沙沟、山神庙咀、大黑沟及辛窑子沟;出土泥河湾披毛犀的地点为下沙沟、大南沟、东谷坨、山神庙咀及辛窑子沟。有疑问的中国犀仅出现在下沙沟,梅氏犀相似种仅出现在辛窑子沟。
关键词:
Abstract
Abundant mammalian fossils were uncovered during the field exploration for Nihewan beds at the beginning of the 1980s along Xinyaozi Ravine at Nangaoya Township of Tianzhen County, Shanxi Province in North China and the studied taxa indicate an age of the early Early Pleistocene. Recent studies on the rhino material not yet described show that there are at least two species of rhinocerotids: Elasmotherium peii and Coelodonta nihowanensis . There might be a third taxon provisionally named as Stephanorhinus cf. S. kirchbergensis due to incompleteness of the specimens. Since its morphometric characters are between S. kirchbergensis and C. nihowanensis , it might be a variety of one of the two species although it is more similar to the former than the latter. In the same way, The rhino specimens from Xiashagou named as Rhinoceros sinensis (?) by
Keywords:
PDF (3880KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
董为, 白炜鹏, 张立民. 山西天镇辛窑子早更新世犀科化石新材料. 古脊椎动物学报[J], 2021, 59(4): 273-294 DOI:10.19615/j.cnki.2096-9899.210715
DONG Wei, BAI Wei-Peng, ZHANG Li-Min.
1 Introduction
The extension of field exploration for Nihewan beds from Yangyuan County of Hebei Province to its western adjacent area at the beginning of the 1980s by the team of Wei Qi (1997) resulted in the discovery of many mammalian fossil localities along Xinyaozi Ravine at Nangaoya Township of Tianzhen County, Shanxi Province in North China. These localities are geographically very close to Xiashagou (=Hsia-sha-kou) locality yielding classical Nihewan fauna (Teilhard de Chardin and Piveteau, 1930). The uncovered fossils systematically studied include Postschizotherium intermedium (Qiu et al., 2002), Hesperotherium sinense (Qiu, 2002), Elaphurus bifurcatus , E. davidianus predavidianus (Dong et al., 2019), Muntiacus bohlini , Cervavitus cf. C. huadeensis , Axis shansius , Nipponicervus elegans , Cervus (Elaphus ) elaphus (Dong et al., 2020) as well as Spirocerus wongi (Bai et al., 2019). The rest of the fossil collection are still in study. The geologic age of the fauna can roughly be considered as contemporary with that of the classical Nihewan fauna (Qiu, 2002), with an estimated age of ca. 2.2-1.7 Ma (Liu et al., 2012). Here we systematically describe the rhinocerotid material from Xinyaozi Ravine at Tianzhen and discuss on the taxonomy of rhinocerotids from the Early Pleistocene in the generalized Nihewan Basin. The dental terminology follows that of Guérin (1980), Deng (2004), Qiu and Wang (2007) and Tong and Wang (2014). The specimens described are housed at the Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences (IVPP).2 Systematic paleontology
Mammalia Linnaeus, 1758Perissodactyla Owen, 1848
Rhinocerotidae Owen, 1840
Rhinocerotinae Owen, 1845
Elasmotheriini Dollo, 1885
Elasmotherium Fischer, 1808
Elasmotherium peii Chow, 1958
(Fig. 1; Table 1)
Elasmotherium sp. Teilhard de Chardin and Piveteau, 1930, p. 19-21
Elasmotherium inexpectatum Chow, 1958, p. 138-139
Elasmotherium sp. Chow, 1958, p. 139
Elasmotherium peii Chow, 1958, p. 139-141
Elasmotherium cf. inexpectatum Chia and Wang, 1978, p. 35-37
Elasmotherium caucasicum Antoine, 2002, p. 43-45
Elasmotherium caucasicum Deng and Zheng, 2005, p. 110-121
Elasmotherium peii Tong et al., 2014, p. 369-388
Material A right mandibular fragment with m2-3 (IVPP V 27428) from Siyangou at Xinyaozi Ravine.
Description The specimen V 27428 (Fig. 1) is a right adult mandibular fragment with m2-3. The preserved fragment indicates that the mandibular body is massive, deep (high) but not very thick. The part in front of the anterior fracture just below the anterior border of m2 of the mandibular body is broken off. The lingual side of the body is slightly convex, and its buccal side is somewhat concave. The ventral margin on the buccal side of the body forms a protrusion similar to a horizontal torus. The depth or height from the alveolar margin to ventral one of the mandibular body measured at m2 is 141.18 mm on buccal side and 153.38 mm on lingual one. And those at m3 are 154.46 and 159.92 mm respectively. The mandibular ramus is partially preserved. The masseteric tuberosity (tuberositas masseterica) is well developed and protrudes laterally (Fig. 1B-C), indicating the powerful muscle of mastication. The mastication fossa (fossa masseterica) is large and somewhat profound, and it extends to the posterior lobe of m3. The pterygoid fossa (fossa pterygoidea) is also deep but less large and just extends to the posterior border of m3. The rest parts of the mandibular ramus, such as coronoid process, condyle process, angular process and mandibular angle are all missing. The specimen is heavily mineralized and resistant to the X ray of CT scanner and the slices obtained are not very clear.
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1A mandibular fragment (IVPP V 27428) of Elasmotherium peii from Xinyaozi Ravine in lingual (A), occlusal (B) and buccal (C) views
The m2 is large, hypsodont and without any cingulid. The exposed crown height and the total crown height estimated on CT images are given in Table 1. The crown is worn, but the occlusal structure can still be seen clearly. The anterior lobe is evidently shorter than the posterior one. The paraconid, protoconid and metaconid are developed. The paralophid, protolophid and metalophid are thick and they connect with each other forming a “U” shape. The trigonid basin, or anterior valley, is moderate and open lingually. The anterior enamel wall of the trigonid basin is smooth, but the buccal wall and posterior wall are folded with many enamel wrinkles. An enamel fold from the paraconid and another one from the protoconid in the trigonid basin fused with each other and form a small vertical shaft in the basin. The buccal enamel layer of the protolophid is also vertically folded. The hypoconid and entoconid of the posterior lobe are also developed. The hypolophid and entolophid are thick, too. They connect with each other forming a flipped “L” shape. The anterior crest of hypoconid, or anterior part of hypolophid, fused with protoconid that links the anterior lobe and the posterior one to form a continuous enamel structure in occlusal view. The talonid basin, or posterior valley, is surrounded by metalophid in the anterior, hypolophid on the buccal side and entolophid in the posterior. It is well developed and widely open lingually. The anterior and buccal enamel walls of talonid basin are strongly folded, but the posterior enamel wall of the talonid basin is just mildly undulated. The buccal enamel layer of the hypolophid is also vertically folded. Both anterior and posterior valleys are filled with cement. The buccal and lingual sides of the crown are covered with cement, but the cement layer on the buccal wall of ectoloph is broken off and only some residue preserved.
Table 1
Table 1Measurements of lower cheek teeth of Elasmotherium peii from Xinyaozi and comparison (mm)
| Taxon | Elasmotherium peii | E. cf. E. peii | E. caucasicum | E. sibiricum | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Location | Xinyaozi | Shanshenmiaozhui1) | Xihoudu2) | Stavropol Krai3) | Salcia4) | Taman5) | Stavropol Krai3) | |||
| m2 L | 74.44 | 75 | 75.5 | 70-77 | 70-77 | 70-95 | 64.2-84 | |||
| m2 W | 40.58 | 40 | 43 | 41.5-60 | 34-36 | 39-50 | 34-43.5 | |||
| m2 H | 29.12(92) | 104 | 44-77 | |||||||
| m3 L | 82.36 | 72-84 | 60-98 | |||||||
| m3 W | 37.78 | 33 | 30-44 | |||||||
| m3 H | 31.28(140) | 129-130 | ||||||||
新窗口打开|下载CSV
The m3 is also large, but longer while narrower than m2 in occlusal view. The morphological structure of m3 is generally the same as that of m2. Nevertheless, the trigonid basin is larger, there is no enamel fold fusing with each other to form any shaft in the basin. The talonid basin is longer than that in m2 and the enamel walls of talonid basin are less folded than those of m2. The entoconid is less developed than that in m2. The total crown height of m3 estimated on CT images is much larger than that of m2 (Table 1).
Comparison and determination The specimen (V 27428) is evidently a large rhinocerotid. The hypsodont crown and strongly folded enamel layers in anterior and posterior valleys or trigonid and talonid basins indicate that it is an Elasmotherium .
Compared with the Elasmotherium peii from Shanshenmiaozhui in Nihewan Basin (Tong et al., 2014), Xinyaozi specimen is very close to those of Shanshenmiaozhui. For example, the anterior lobe is shorter than the posterior one, the trigonid basin or anterior valley of lower molars is much smaller than talonid basin or posterior valley, and the enamel folds in both basins or valleys and on buccal side are well developed. The dimensions of crowns are also close to each other (Table 1).
Compared with the specimens from Xihoudu (Hsihoutu) identified as Elasmotherium cf. inexpectatum by Chia and Wang (1978) and reassigned into Elasmotherium peii by Tong et al. (2014), the comparable material is the m2. The morphology of the specimens from the two localities is close to each other, the crown dimensions are also similar (Table 1). However, the trigonid basin is larger in Xihoudu specimen, and enamel folds are less developed in trigonid basin in Xihoudu specimen. The enamel folds on the anterior wall of talonid basin are much more developed in Xihoudu specimen than in Xinyaozi one, and those on the buccal wall of talonid basin are on the contrary. The difference is likely due to the different degree of the crown abrasion with age, Xinyaozi specimen is younger than that of Xihoudu.
Compared with the Elasmotherium cf. E. peii from Salcia in Moldova (David and Eremeico, 2003), Xinyaozi specimen is also similar to the counterpart specimen numbered 52-563 from Salcia, e.g. developed enamel folds in both anterior and posterior valleys of lower molars, anterior lobe is evidently shorter than the posterior one, the dental dimensions are similar (Table 1), etc. Nevertheless, the metaconid, paraconid and paralophid in Xinyaozi specimen are more developed than in Salcia specimen, the trigonid basin or anterior valley is smaller in Xinyaozi specimen than in Salcia one.
As to other species of Elasmotherium , the dental measurements of Elasmotherium peii from Xinyaozi are within the ranges of those of both E. caucasicum from Taman and E. sibiricum from Stavropol Krai (Table 1). Xinyaozi specimen differs from Taman ones (Borissiak, 1914) by longer length of m3 than that of m2, less enamel folds in trigonid and talonid basins, smaller trigonid basin in m2. It differs from those of E. sibiricum (Antoine, 2002) by less enamel folds on posterior side of paralophid and anterior side of entolophid in m2.
The first Elasmotherium from Nihewan Basin was reported as some limb bone fragments of radius, semilunar, metacarpus and metatarsus, etc. but without any dental material. The morphological information the specimens offered is so limited that it allowed only generic taxonomic determination as Elasmotherium sp. (Teilhard de Chardin and Piveteau, 1930). Some additional materials were collected later from the same locality and were noticed as “Elasmotherium sp.” by Teilhard de Chardin and Leroy (1942), including a molar fragment identified as a posterior buccal fragment of a left M2 (Chow, 1958). Deng and Zheng (2005) described the limb bone material from Nihewan Basin at the collection of Tianjin Natural History Museum and included them into E. caucasicum as suggested by Antoine (2002). Tong et al. (2014) described elasmothere material from Shanshenmiaozhui in Nihewan Basin and assigned the specimen into E. peii . The latter was erected by Chow (1958) for some elasmothere specimens from Shanxi Province together with another new species E. inexpectatum . E. peii was reported again found at Xihoudu Paleolithic Site (Chia and Wang, 1978). Nevertheless, Chinese specimens were all regrouped into E. caucasicum by Antoine (2002). Tong et al. (2014) compared Chinese material with Russian one and indicated that E. peii differed clearly from both E. caucasicum and E. sibiricum and its taxonomic status should be retained. Based on the present work on E. peii from Xinyaozi and related comparisons, the present authors agree to maintain the taxonomic status of E. peii . Elasmotheres are very large rhinos and their taxonomic diversity is reasonably limited by their size and food requirement. It may be true that all Elasmotherium species might represent a single genetically valid species, but for the cases in paleontology with fossils, it is reasonable to classify the specimens according to their morphology with geographic consideration.
Dicerorhinini Loose, 1975
Stephanorhinus Kretzoi, 1942
Stephanorhinus cf. S. kirchbergensis (Jäeger, 1839)
(Figs. 2-6; Tables 2-4)
Material A right juvenile maxillary fragment (IVPP V 27429.1) from Xinyaozi Ravine; A pair of broken mandibles (V 27429.2) from Taijiaping at Xinyaozi Ravine; three broken left mandibles (V 27429.3-5) from Xinyaozi Ravine.
Description The right juvenile maxillary fragment (V 27429.1) is preserved with complete DP1-3 and broken DP4 (Fig. 2). Its dental measurements are listed in Table 2.
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2A right juvenile maxillary fragment (IVPP V 27429.1) of Stephanorhinus cf. S. kirchbergensis from Xinyaozi Ravine in lingual (A), occlusal (B) and buccal (C) views
Table 2
Table 2Measurements of upper deciduous cheek teeth of Stephanorhinus cf. S. kirchbergensis from Xinyaozi and comparison (mm)
| V 27429.1 | Xiashagou1) | Rhino Cave2) | Huludong3) | Zhoukoudian4) Loc. 1 | Zhoukoudian5) Loc. 13 | West Europe6) | |
|---|---|---|---|---|---|---|---|
| DP1 L | 25.38 | 26 | 31.8-32.9 | 28-29 | 27.5 | 35 | 26-32 |
| DP1 W | 20.06 | 24 | 26-27.9 | 28 | 24-25 | 32 | 31-34 |
| DP1 H | 24.34 | 26 | |||||
| DP2 L | 35.08 | 37 | 44-47.3 | 38-39 | 36.5 | 41 | 34-39.5 |
| DP2 W | 32.76 | 40 | 42.6-47.9 | 40-42 | 39.5 | 40 | 31-43 |
| DP2 H | 28.48 | 25 | |||||
| DP3 L | 41.08 | 50.6-55.8 | 47-48 | 43.5 | 51 | 41.5-50.5 | |
| DP3 W | 39.12 | 48.2-52 | 50-52 | 49 | 53 | 42-54.5 | |
| DP3 H | 35.18 | 36.5 | |||||
| DP4 L | 49.52 | 52 | 56.6-60 | 48-52 | 47 | 53 | 48-57 |
| DP4 W | 34.24 | 51 | 52.6-54.5 | 54 | 48.5 | 56 | 51-57.5 |
| DP4 H | 42.56 | 42 | |||||
| L DP1-4 | 145.32 | 172.8-183 | 175-177 | 147 | 150 |
新窗口打开|下载CSV
The DP1 is clearly composed of four main cusps, e.g. paracone, protocone, metacone and hypocone. Metacone is the largest cusp and its anterior crest fused with the posterior crest of paracone forming ectoloph. The parastyle is located in the anterior of the crown and its posterior crest fused with the anterior one of the paracone that the buccal side of the crown is formed by parastyle, paracone, ectoloph and metacone in a consecutive chain. The protocone is moderate in size and oriented obliquely in occlusal view. Its anterior crest extends buccally and fused with the anterior crest of parastyle forming protoloph. A posterior crest connects metacone and hypocone forming metaloph. A medisinus separates protocone from hypocone. A large medifossette is encircled by protocone, protoloph, parastyle, paracone, ectoloph, metacone, metaloph and hypocone. The crochet is absent that medifossette is directly connected to the medisinus. A postfossette is located posterior to the metaloph between hypocone and metacone. It is much smaller than medifossette. Precingulum is present, but very weak. Postcingulum is present and moderate.
The DP2 is well molarized and in “π” shape in occlusal view. The crown is composed of four main cusps and three main lophs. Both parastyle and precingulum are developed. The crista protruding from paracone is present but weak. It divides the central depression between protoloph and metaloph into prefossette (anterior fossette) 1) and medifossette. The crochet extending from anterior side of metaconule is developed and set between medifossette and medisinus, but does not reach the crista and the medifossette is still widely open. Antecrochet and metastyle are absent. Ectoloph is much higher than protoloph and metaloph. Postfossette and postcingulum are developed. Entocingulum is present and moderate.
The DP3 is similar to DP2, but its dimensions are much larger, and its prefossette and medifossette are relatively small, while the medisinus is much larger.
The DP4 is not well preserved. The anterior lingual part of the crown, including protocone, protoloph, precingulum, etc. are broken off in a fragment. And the fragment was incorrectly adhered to its original location by lied down position during preparation. However, its morphology can still be observed in general. It is similar to DP3 but with larger dimensions.
The enamel layers of all upper deciduous cheek teeth are generally smooth, the crowns are relatively hypsodont and quite ectolophodont.
The specimens V 27429.3-5 are three left mandibular fragments with deciduous cheek dentitions (Figs. 3-4). Specimen V 27429.3 is relatively well preserved with large part of mandibular body, a small part of mandibular ramus and complete deciduous dentition (Fig. 3). The body is thick in occlusal view, posteriorly deep (high) but anteriorly shallow (low) in lateral view. The dp4 is located just behind the symphysis. The specimens V 27429.4-5 are more fragmental, with dp2-4 and dp3-4 respectively (Fig. 4). The enamel and dentine layers of lower deciduous cheek teeth are thin, while the cavities are large (Fig. 4F). The lower deciduous dental measurements are listed in Table 3.
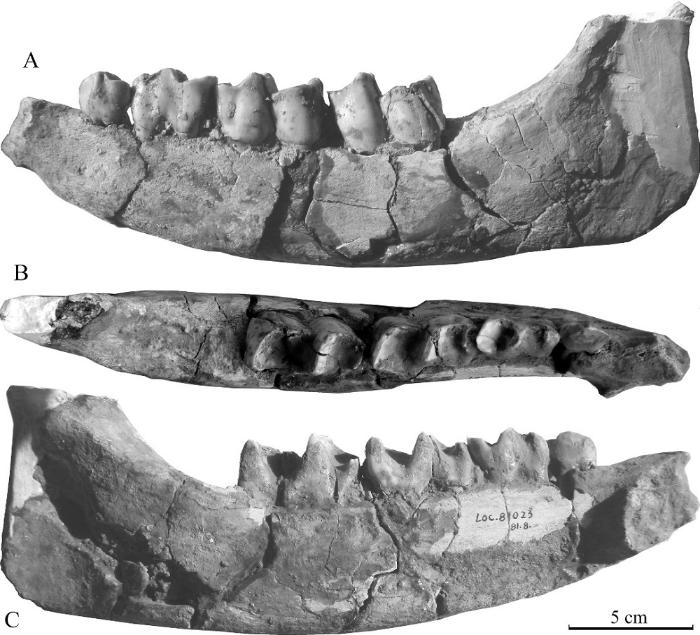 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3A broken mandible (IVPP V 27429.3) of Stephanorhinus cf. S. kirchbergensis from Xinyaozi Ravine in buccal (A), occlusal (B) and lingual (C) views
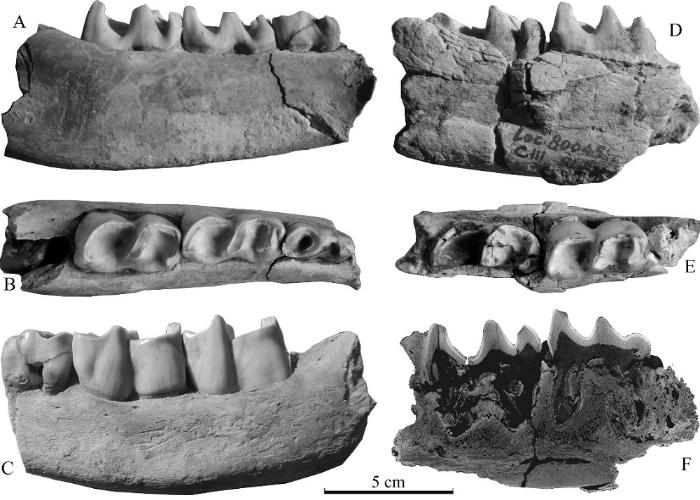 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 4Mandibular fragments (IVPP V 27429.4-5) of Stephanorhinus cf. S. kirchbergensis from Xinyaozi Ravine
A-C. V 27429.4; D-F. V 27429.5; A, D. lingual view; B, E. occlusal view; C. buccal view; F. sagittal slice of CT image
Table 3
Table 3Measurements of lower deciduous cheek teeth of Stephanorhinus cf. S. kirchbergensis from Xinyaozi and comparison (mm)
| V 27429.3 | V 27429.4 | V 27429.5 | Rhino Cave1) | Loc. 13 Zhoukoudian2) | West Europe3) | |
|---|---|---|---|---|---|---|
| dp1 L | 20.56 | 23-23.4 | 21 | 17.5-23 | ||
| dp1 W | 11.38 | 11.6-13 | 13 | 14 | ||
| dp1 H | 19.82 | 18.5 | ||||
| dp2 L | 32.42 | 32.32 | 30.5-38.8 | 35 | 28-38 | |
| dp2 W | 18.44 | 18.94 | 16.7-20 | 21 | 16.5-21.5 | |
| dp2 H | 25.02 | 25.12 | ||||
| dp3 L | 44.12 | 46.98 | 45.78 | 49.2 | 44 | 38-44.5 |
| dp3 W | 23.22 | 24.98 | 25.44 | 25.1 | 26 | 21-28.5 |
| dp3 H | 28.84 | 30.26 | 32.82 | 28.5 | ||
| dp4 L | 44.82 | 45.18 | 47.68 | 49.5-52 | 47 | 42-51 |
| dp4 W | 23.22 | 27.46 | 21.54 | 27.1-27.4 | 29 | 21-31 |
| dp4 H | 32.12 | 34.14 | 23.14 | 30-35.5 | ||
| L dp1-4 | 141.68 | 182-201 |
新窗口打开|下载CSV
The dp1 is relatively small and composed of a main cusp, protoconid, and some minor cusps such as metaconid and hypoconid. Talonid basin or posterior valley is present, but weak. Ectoflexid is absent.
The dp2 is composed of five main cusps and three lophids. The protoconid and hypoconid are relatively larger. The parastylid is developed. The entolophid is wider than metalophid, and the paralophid is the shortest. Metalophid is obliquely oriented and its angle with mandibular body is less than 45°. Talonid basin is larger than trigonid one. Cingulid and ectoflexid are not evident.
The dp3 is also composed of five main cusps and three lophids, but paralophid and metalophid are much wider than in dp2. Parastylid is well developed. Metalophid is also obliquely oriented but its angle with mandibular body is larger than 45°. The posterior lobe of the dp3 is somewhat selenodont in occlusal view. The metalophid is evidently higher than the hypolophid and the ectoflexid is developed.
The dp4 is similar to dp3, but without parastylid and the trigonid basin is larger than that in dp3.The specimen V 27429.2 is a pair of broken mandibles with complete permanent cheek dentitions (Fig. 5). The anterior part of symphysis is missing, and the posterior end of the symphysis is located below p3. The lower part of mandibular body is not well preserved, and the major part of mandibular ramus is lost. The preserved parts of the specimen indicate that the mandibular body is robust. It curves slightly downward in lateral view and slightly medially in occlusal view. The measurements of lower cheek teeth are listed in Table 4.
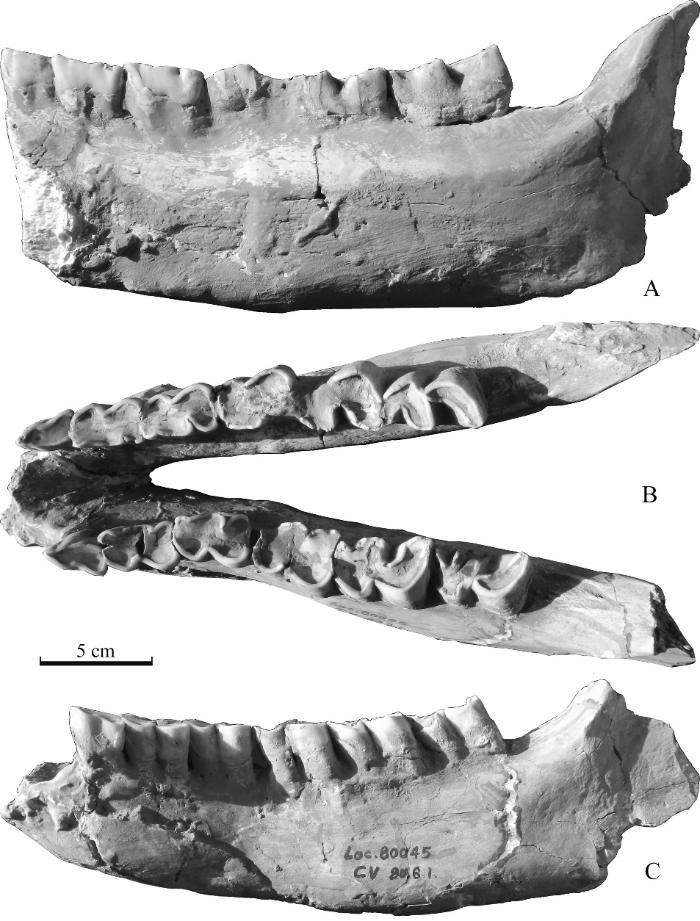 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 5Broken mandible (IVPP V 27429.2) of Stephanorhinus cf. S. kirchbergensis from Taijiaping in Xinyaozi Ravine in lingual (A), occlusal (B) and buccal (C) views
Table 4
Table 4Measurements of lower cheek teeth of Stephanorhinus cf. S. kirchbergensis from Xinyaozi and comparison (mm)
| IVPP V 27429.2 | S. kirchbergensis | |||||||
|---|---|---|---|---|---|---|---|---|
| left | right | Nihewan1) | Rhino Cave2) | Loc. 1 Zhoukoudian3) | Anping4) | West Europe5) | ||
| p2 L | 28.66 | 26.88 | 26.7-29.2 | 30.5-33.5 | 26-32 | 29.1-32.7 | 30.5-37 | |
| p2 W | 17.06 | 17.04 | 17.2-18 | 18.7-20.5 | 17-23 | 18.5-21.8 | 18.5-23 | |
| p2 H | 21.72 | 19.18 | 20-36 | 28-40 | ||||
| p3 L | 32.56 | 31.14 | 26.3 | 34.5-36.8 | 33-38 | 32.6-41.1 | 40-46.5 | |
| p3 W | 22.84 | 22.48 | 24 | 25.9-28.6 | 21-29 | 26.5-28.4 | 23.5-30 | |
| p3 H | 20.58 | 18.52 | 17-32 | 36 | ||||
| p4 L | 35.88 | 35.12 | 38-38.4 | 42-43.4 | 40-49 | 37.9-42.8 | 39-49 | |
| p4 W | 25.02 | 25.36 | 28-28.6 | 30-32.7 | 30-36 | 29.6-33.7 | 25.5-36 | |
| p4 H | 22.68 | 21.92 | 25-34 | 28.5-53 | ||||
| m1 L | 39.34 | 37.52 | 43-43.3 | 41.8-47.9 | 40-45 | 44.1-51.1 | 46.5-56 | |
| m1 W | 30.28 | 29.18 | 31.5-31.9 | 33.1-37 | 29-33 | 30.8-37.1 | 27-37 | |
| m1 H | 15.98 | 15.82 | 17-31 | 40-58 | ||||
| m2 L | 46.86 | 42.42 | 48-48.7 | 51.8-56.5 | 38-53 | 49.5-54.7 | 51-63 | |
| m2 W | 31.92 | 32.08 | 30.9-31.8 | 34.6-36.3 | 29-34 | 32.1-36.3 | 30-40 | |
| m2 H | 23.52 | 22.08 | 22-49 | 42-54 | ||||
| m3 L | 47.02 | 48.76 | 46.3-46.8 | 52.1-60.4 | 47-57 | 52.1-55.8 | 52-68 | |
| m3 W | 28.24 | 29.08 | 29.4-29.5 | 32.6-35.7 | 30-38 | 31.7-32.8 | 28-40 | |
| m3 H | 26.08 | 26.02 | 39-47 | 40-64 | ||||
| L p2-4 | 89.62 | 88.42 | ||||||
| L m1-3 | 130.42 | 129.08 | 130-137 | |||||
| L p2-m3 | 219.96 | 214.38 | 248.1 | |||||
新窗口打开|下载CSV
The lower cheek teeth of the specimen are much worn. The p2 is composed of two lobes, the posterior one is larger than the anterior one and the anterior part of the posterior lobe is largely fused with the posterior part of the anterior lobe. The anterior lobe and the posterior one are separated by moderately sized ectoflexid. The p2 differs from p3 and p4 by the presence of parastylid. The p3 and p4 are both molarized and formed by anterior and posterior lobes, the p4 is larger than p3. The m1-3 are also composed of anterior and posterior lobes; the talonid basin is larger than trigonid basin; the ectoflexid is well developed. A weak precingulid is present on the lingual base of paralophid in the p3-m3, but the most cingulids are generally absent. The enamel layer of cheek teeth is smooth, and the paraconid rib is absent in all check teeth. Although the crowns are much worn, it can still be observed on p4 and m3 that the angle formed by protolophid and metalophid is close to a right angle.
Comparison and determination The morphology of V 27429.1 (Fig. 2) is similar to that of Stephanorhinus kirchbergensis . For example, the enamel layers of cheek teeth are generally smooth, the crown is relatively hypsodont, upper cheek teeth quite ectolophodont, i.e. buccal loph and cusps evidently higher than the lingual ones, and antecrochet and metastyle are absent from upper cheek teeth. It is very similar to that of juvenile maxillary fragments Nos. 246 and 256 from Rhino Cave in Shennongjia (Tong and Wu, 2010). However, the dimensions of V 27429.1 are smaller than those from Rhino Cave, slightly smaller than those from Huludong, Locs. 1 & 13 of Zhoukoudian, and close to the lower range limit of those from West Europe (Table 2).
Compared with S. kirchbergensis from other localities, the morphology and dental dimensions of V 27429.3-5 (Figs. 3-4) are close to those of Loc. 13 of Zhoukoudian and Rhino Cave in Shennongjia, e.g. the enamel surface of cheek teeth is smooth, parastylid is well developed but without paraconid rib on dp2 and dp3. Their dental dimensions are also close to those of West Europe (Table 3).
Compared with S. kirchbergensis from other localities, the morphology of V 27429.2 (Fig. 5) is similar to that of VM.670 from Loc. 1 of Zhoukoudian (Chow, 1979), TNP00315 from Daheigou and SGT2:1 from Shigou in Nihewan Basin (Tong et al., 2014), as well as that of two mandible specimens from Rhino Cave in Shennongjia (Tong and Wu, 2010). On the other hand, the dental dimensions of V 27429.2 are generally smaller than those from other localities, i.e. Anping (Zhang et al., 1980; Xu, 1986) and West Europe (Guérin, 1980) (Table 4). Fig. 6 illustrates that Xinyaozi specimen is closer to Coelodonta than to S. kirchbergensis , but the smooth surface of enamel in both upper and lower dentitions, the absence of paraconid rib in lower cheek teeth distinguish Xinyaozi specimens from those of Coelodonta .
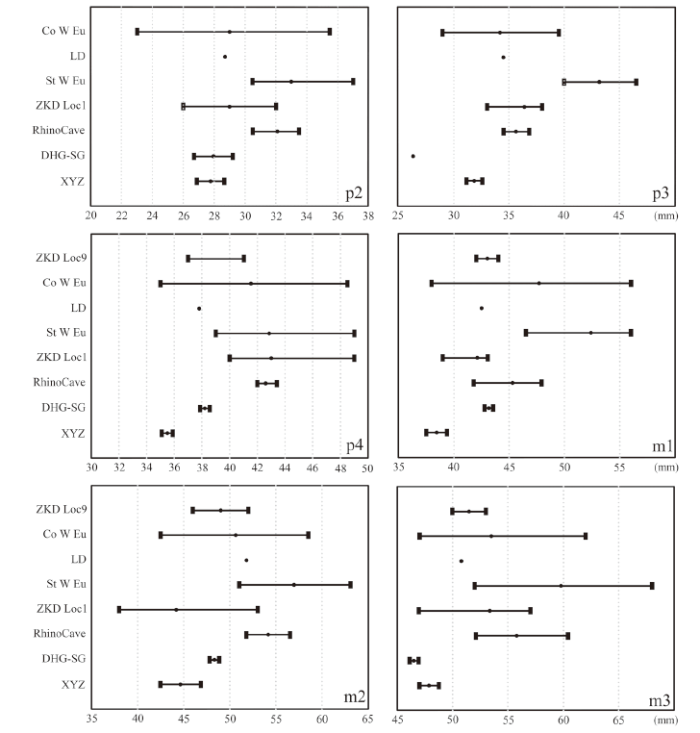 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 6Length range comparison of lower cheek teeth from selected localities
Abbreviations and data sources: ZKD Loc9, Coelodonta antiquitatis from Locality 9 at Zhoukoudian (Teilhard de Chardin, 1936); Co W Eu, C. antiquitatis from West Europe (Guérin, 1980); LD, C. nihowanensis from Longdan (Qiu et al., 2004); St W Eu, Stephanorhinus kirchbergensis from West Europe (Guérin, 1980); ZKD Loc1, S. k. from Locality 1 of Zhoukoudian (Chow, 1979); RhinoCave, S. k. from Shennongjia (Tong and Wu, 2010); DHG-SG, S. k. from Daheigou and Shigou in Nihewan Basin (Tong et al., 2014); XYZ, Xinyaozi Ravine
Compared with the “Rhinoceros sinensis (?)” from Xiashagou (Teilhard de Chardin and Piveteau, 1930), the upper deciduous dentitions from both Xinyaozi and Xiashagou share the following characters such as smooth buccal enamel and convex ectoloph, developed crochet, but the crista is more developed and dimensions are slightly larger (Table 2) in Xiashagou specimens.
In general, the above comparisons indicate that the upper and lower deciduous dentitions are close to S. kirchbergensis from related localities both morphologically and metrically, while the adult lower cheek teeth are morphologically close to those of S. kirchbergensis from related localities, but with certain metrical differences. Since S. kirchbergensis is considered mostly distributed in the Middle Pleistocene, with some survivors to the Last Glaciation (e.g. Guérin, 1980; Fortelius et al., 1993), the discovery of S. kirchbergensis from Shigou in Nihewan Basin extended the earliest appearance of S. kirchbergensis to the late Early Pleistocene (Tong et al., 2014). The geological age of Xinyaozi deposits were dated as contemporary with Xiashagou fauna, i.e. the early Early Pleistocene (Qiu, 2002; Qiu et al., 2002), the appearance of S. kirchbergensis in such age is thus questionable, although if Xinyaozi specimens can be confirmed as true S. kirchbergensis it will extend its earliest appearance to the early Early Pleistocene. The Xinyaozi specimens are morphologically similar to those of S. kirchbergensis , but metrically smaller. We place them therefore as conformis species of S. kirchbergensis for prudence because the only adult specimen is worn, although the juvenile specimens are quite certain to be included into S. kirchbergensis .
Coelodonta Bronn, 1831
Coelodonta nihowanensis Kahlke, 1969
(Figs. 7-9; Tables 5-6)
Rhinoceros cf. tichorhinus Teilhard de Chardin and Piveteau, 1930, p. 17-19
Coelodonta nihowanensis Kahlke, 1969, p. 701-702
Coelodonta nihowanensis Chow, 1978, p. 48
Coelodonta antiquitatis Li, 1984, p. 64-65
Coelodonta antiquitatis Wei et al., 1985, p. 227
Coelodonta antiquitatis Tang et al., 1995, p. 79
Coelodonta nihowanensis Qiu et al., 2004, p. 119-125; p. 186-187
Coelodonta nihowanensis Tong and Wang, 2014, p. 710-724
Material A maxillary fragment (IVPP V 27430.1) with left and right deciduous cheek teeth from Taijiaping at Xinyaozi Ravine; an isolated right dp3 (V 27430.2) and an isolated right dp4 (V 27430.3) from Xinyaozi Ravine.
Description The maxillary fragment (V 27430.1) is preserved with left DP1-4 and right DP1-3 (Fig. 7; Table 5). The palatine is flat. The width between lingual edges of DP1s measures 29.2 mm and that of DP3s 30.9 mm.
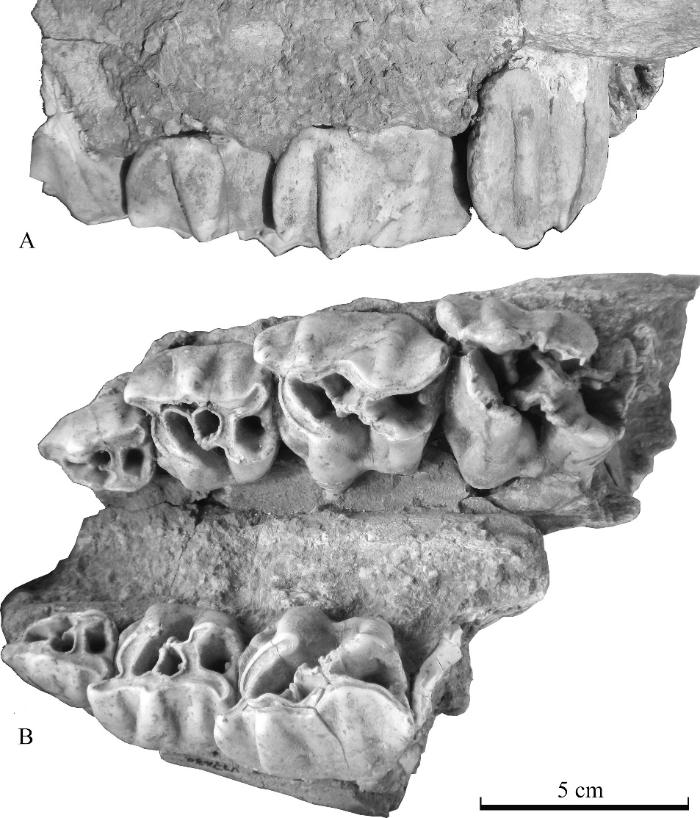 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 7Maxillary fragment (IVPP V 27430.1) of Coelodonta nihowanensis from Xinyaozi Ravine in left buccal (A) and occlusal (B) views
Table 5
Table 5Measurements of upper deciduous cheek teeth of Coelodonta nihowanensis from Xinyaozi and comparison (mm)
| V 27430.1 | C. nihowanensis | C. antiquitatis | C. tologoijensis | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| left | right | Xiashagou1) | Danangou2) | SSMZ3) | Longdan4) | Dingcun5) | W. Europe6) | W. Transbaikalia7) | ||||
| DP1 L | 29.48 | 29.04 | 32 | 27 | 26-27 | 27.3 | 26 | 21-21.5 | 22-27 | |||
| DP1 W | 22.04 | 21.68 | 23 | 20 | 18.2-20 | 21.8 | 19 | 17-18 | 17-20 | |||
| DP1 H | 21.34 | 22.64 | ||||||||||
| DP2 L | 34.78 | 34.32 | 35 | 34.8 | 31.5-32.5 | 34.5 | 27.5 | 28-33 | 28-32 | |||
| DP2 W | 35.08 | 33.76 | 34 | 32 | 29.2-30.3 | 35 | 32 | 26-32.5 | 28-30 | |||
| DP2 H | 26.58 | 27.56 | 23-30 | |||||||||
| DP3 L | 45.82 | 46.46 | 47 | 45 | 42.6-44.7 | 41.7 | 40-48 | 31-45.5 | 43-49 | |||
| DP3 W | 41.02 | 40.18 | 40 | 39 | 37.6-38.3 | 40.8 | 37-45 | 31.5-42 | 35-41 | |||
| DP3 H | 34.22 | 35.28 | 25-33.5 | |||||||||
| DP4 L | 49.54 | 54 | 46.9 | 50.2 | 55-55.5 | 44-57 | ||||||
| DP4 W | 43.46 | 40 | 41.4 | 46 | 40-43 | 39-55 | ||||||
| DP4 H | 35.28 | 33.5-46 | ||||||||||
| L DP1-4 | 145.62 | 152 | 136 | 150 | 128-141 | |||||||
新窗口打开|下载CSV
The DP1 is roughly triangular in occlusal view and composed of four main cusps with the protocone the smallest. The protocone is developed and its postprotocrista extends posteriorly with tendency to close the medisinus. The parastyle is well developed. It fuses with paracone and metacone forming ectoloph. The paracrista from the paracone extends lingually and divides the trigon basin into prefossette and medifossette. The crochet from metaconule is developed and extends forward and nearly closing medifossette. Antecrochet is absent. The metaloph is evidently wider than the protoloph. The postcingulum is very developed forming the posterior wall of postfossette.
The DP2 is well molarized and composed of four main cusps and three main lophs. The parastyle is very developed. The precingulum is well developed. The crista from paracone protrudes forwards and meets the crochet from metaconule to close the medifossette completely. The prefossette is short but very wide and open to medisinus. The postfossette is developed and postcingulum is moderate.
The DP3 is similar to DP2 but with larger dimensions.
The DP4 is similar to DP3, but its dimensions are larger and the crista is relatively low.
The lower dentition is represented by a dp3 and a dp4 only. The measurements are listed in Table 6.
Table 6
Table 6Measurements of lower deciduous cheek teeth of Coelodonta nihowanensis from Xinyaozi and comparison (mm)
| C. nihowanensis | C. antiquitatis | C. tologoijensis | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| V 27430.2 | V 27430.3 | Xiashagou1) | Danangou2) | Shanshenmiaozui3) | Dingcun4) | W. Europe5) | W. Transbaikalia6) | |||
| dp3 L | 40.08 | 38 | 41 | 38.6-40 | 37-38.2 | 34.5-41.5 | 36-38 | |||
| dp3 W | 19.98 | 18 | 21 | 21-23 | 19.5-21.3 | 17-24 | 16-18 | |||
| dp3 H | 32.76 | 24.5-36 | ||||||||
| dp4 L | 45.12 | 46 | 44 | 41.5-45 | 45-23 | 37-51 | 39-43 | |||
| dp4 W | 24.02 | 22 | 21 | 21.2-25 | 19-25.5 | 19-21 | ||||
| dp4 H | 32.66 | 27-46 | ||||||||
新窗口打开|下载CSV
The dp3 (V 27430.2) is composed of an anterior and a posterior lobes. The parastylid is not evident. The paraconid rib is absent or at least very weak. The protoconid rib is present and moderate. The protolophid is concave in buccal view. The trigonid and talonid basins are well developed and widely open. The ectoflexid is also well developed. The angle between the protolophid and metalophid is evidently an obtuse one, as well as that between the hypolophid and entolophid. The dp4 (V 27430.3) is morphologically similar to dp3 (Fig. 8), but with much larger dimensions (Table 6).
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 8Deciduous cheek teeth of Coelodonta nihowanensis from Xinyaozi Ravine
A-C. right dp3 (IVPP V 27430.2); D-F. right dp4 (V 27430.3) A, D. buccal view; B, E. occlusal view; C, F. lingual view
Comparison and determination The described specimen V 27430.1 with DP1-4 is characterized by the undulation of ectoloph, closure of medifossette; crochet, crista, paracone rib and metacone rib, parastyle and metastyle are well developed. The lower deciduous cheek teeth V 27430.2-3 by the presence of protoconid rib and undulation of protolophid and hypolophid. These traits distinguish them from those of S. kirchbergensis and they are the same as those from Shanshenmiaozhui and in accordance with the diagnosis of Coelodonta revised by Qiu et al. (2004). V 27430.1 is very close to the specimen reassigned to C. nihowanensis by Tong and Wang (2014) and that figured in fig. 3 of plate II by Teilhard de Chardin and Piveteau (1930), e.g. the undulation of buccal enamel of ectoloph, the closure of medifossette, absence of antecrochet, etc. Its dental dimensions are also close to those of C. nihowanensis from Xiashangou (Teilhard de Chardin and Piveteau, 1930), Shanshenmiaozhui (Tong and Wang, 2014), Longdan (Qiu et al., 2004), slightly different from those of C. antiquitatis from Dingcun (Pei, 1958) and West Europe (Guérin, 1980), somewhat different from those of S. kirchbergensis from Xinyaozi, Locs. 1 and 13 of Zhoukoudian (Table 5; Fig. 9). Both upper and lower dental dimensions of Xinyaozi specimens (Tables 5-6) are larger than those of C. tologoijensis from Transbaikalia (Beliajeva, 1966). The logarithm dimensions of upper deciduous cheek teeth comparison (Fig. 9) shows that those of V 27430.1 are the closest to those of C. nihowanensis from Longdan, then Xiashagou, and then Shanshenmiaozhui. Those of C. antiquitatis from Dingcun and S. kirchbergensis from Xinyaozi (V 27429.1) are also similar. And those of C. antiquitatis from West Europe are slightly smaller and those of S. kirchbergensis from Locs. 1 and 13 of Zhoukoudian are slightly larger.
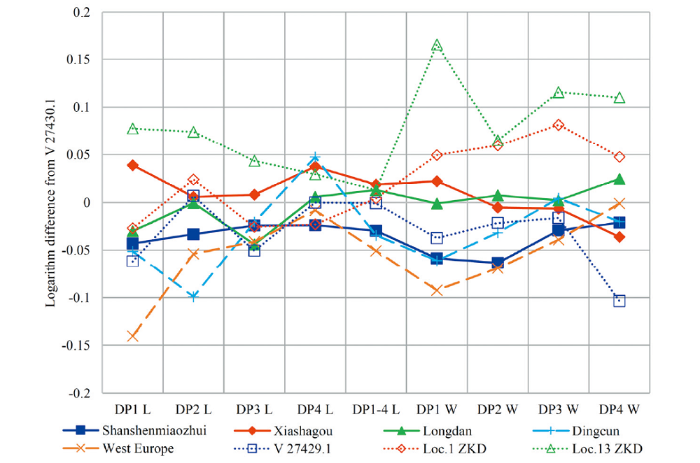 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 9Ratio diagram of dimensions of upper deciduous cheek teeth from related localities, plotted against those of IVPP V 27430.1 (reference 0)
Note: Data source as in Tables 2, 5. L. length; W. width; ZKD. Zhoukoudian; solid line, Coelodonta nihowanensis ; dash line, C. antiquitatis ; dotted line, Stephanorhinus kirchbergensis
3 Discussion and conclusion
Xinyaozi Ravine is located in the west part of generalized Nihewan Basin (s. l. ), or adjacent to the west side of classical Nihewan Basin (s. s. ). The Xinyaozi deposits yielding mammalian fossils were correlated to those of Xiashagou described by Teilhard de Chardin and Piveteau (1930), i.e. the Early Pleistocene (Qiu et al., 2002; Qiu, 2002). The first description of rhinos from the basin was published by Teilhard de Chardin and Piveteau (1930) on the fossils collected in the 1920s. The rhino remains from Xiashagou and surrounding areas were abundant, but mostly limb bones with some fragmental dentitions and without complete cranial material that made the taxonomic determination difficult. Teilhard de Chardin and Piveteau (1930) assigned the rhino specimens into three taxa, i.e. Rhinoceros sinensis (?), Rhinoceros cf. tichorhinus and Elasmotherium sp. The presence of Elasmotherium in the Nihewan Basin was confirmed successively by Deng and Zheng (2005), Tong et al. (2014). The further understanding of the elasmothere from the basin is that the rhino has been precisely assigned to Elasmotherium peii (Tong et al., 2014). The elasmothere specimens are so characteristic, i.e. massive mandible and dentitions, strongly folded enamel layers etc., that they can be distinguished clearly from other rhinos and its presence in the Early Pleistocene deposits of the basin is further confirmed. The elasmothere taxa are not numerous in China and mostly range from the Late Miocene to the Early Pleistocene and distributed in northwestern and northern China (Teilhard de Chardin and Piveteau, 1930; Deng, 2003, 2005, 2008; Deng and Zheng, 2005; Deng et al., 2013; Tong et al., 2014). The “Rhinoceros cf. tichorhinus ” from the basin was reassigned into Coelodonta nihowanensis named by Kahlke (1969) and Chow (1978) independently as a new species of Early Pleistocene Coelodonta , together with the specimens from Gonghe, Lingyi and Zhoukoudian. The Gonghe specimens were later excluded from C. nihowanensis by Zheng et al. (1985). This approach was accepted by later authors (e.g. Qiu et al., 2004; Tong and Wang, 2014). In addition, the specimens from Danangou (Li, 1984) and Donggutuo (Wei et al., 1985) were reassigned from Coelodonta antiquitatis to C. nihowanensis by Tong and Wang (2014). The Xinyaozi specimens can clearly be included into C. nihowanensis . The presence of the species at Xinyaozi Ravine added a new locality yielding C. nihowanensis in the basin besides Xiashagou, Danangou, Donggutuo and Shanshenmiaozhui. The famous wooly rhino (Coelodonta ) widespread in northern Eurasia in the Late Pleistocene (Boule et al., 1928; Qi, 1975; Jiang, 1977; Tang et al., 1983; Prothero et al., 1986; Pei, 2001; Kahlke and Lacombat, 2008; Tong et al., 2011; Hou et al., 2013; Zhang et al., 2016) had an Early Pleistocene ancestor with a large population in the generalized Nihewan Basin. Their farther ancestor can be traced back to the Middle Pliocene (Deng, 2002; Deng et al., 2011).The specimens of the Rhinoceros sinensis (?) described by Teilhard de Chardin and Piveteau (1930) were problematic. Teilhard de Chardin and Piveteau placed some maxillary fragments with deciduous or permanent cheek teeth and some fragmental lower cheek teeth from Xiashagou into “R. sinensis (?)”, but named the specimens as “R . cf. sinensis ” in figure and plate captions. They indicated that Xiashagou specimens differed from R. sinensis from Sichuan (Sze-chuan) of Central China (Colbert and Hooijer, 1953) by their smooth enamel of ectoloph of premolars and the pattern of P2. They considered that Xiashagou specimens differed also from Rhinoceros etruscus (Stephanorhinus etruscus ) and Rhinoceros mercki (S. kirchbergensis ), but the differences from S. kirchbergensis were slight. R. sinensis was not clearly defined when it was named and it served for quite a long time as a “waste basket” taxon for most Pleistocene rhino specimens from southern China (Tong and Moigne, 2000; Yan et al., 2014). It is understandable why Teilhard de Chardin and Piveteau (1930) included Xiashagou specimens into this taxon with a question mark. Yan et al. (2014) redefined the diagnosis of R. sinensis . Xiashagou specimens are in accordance with such diagnosis by some characters such as the well-developed crochet, absence of crista and antecrochet in the adult, and moderately developed postfossette, but the protocone constriction is not distinct on upper molars and protoloph does not extend to the median valley as in typical R. sinensis . On the other hand, Xiashagou specimens have some characters similar to those of S. kirchbergensis , e.g. highly molarized premolars, absence of antecrochet and metastyle, smooth enamel surface, ectolophodont upper cheek teeth. It is, therefore, likely a variety of S. kirchbergensis . R. sinensis ranges in the Middle Pleistocene (Colbert and Hooijer, 1953; Tong and Moigne, 2000; Tong, 2002; Yan et al., 2014) and distributed mostly in central and southern China. If the presence of R. sinensis in the Nihewan Basin can be further proved, it will be its earliest and northernmost records.
Although the specimens of Stephanorhinus cf. S. kirchbergensis from Xinyaozi are morphometrically similar to Coelodonta nihowanensis in a certain way, and somewhat similar to Xiashagou “R. sinensis (?)”, it is the most similar to S. kirchbergensis as mentioned in above section. Due to the incompleteness of the specimens, the morphometric information available is not sufficient for precise determination before the discovery of more complete material.
To conclude, there are at least two forms of rhinos from the Xinyaozi Ravine, Elasmotherium peii and Coelodonta nihowanensis , with a probable third form temporally assigned into S. cf. S. kirchbergensis . If its taxonomic status can be further confirmed, it will be the earliest record of S. kirchbergensis , while it would also be a variety C. nihowanensis . The Early Pleistocene rhinos in generalized Nihewan Basin include therefore two confirmed taxa such as E. peii and C. nihowanensis , as well as two uncertain taxa such as R. sinensis (?) and S . cf. S. kirchbergensis , which might be two varieties of C. nihowanensis or S. kirchbergensis , or of both respectively.
Acknowledgements
The present work was supported by the Strategic Priority Research Program of Chinese Academy of Sciences, Grant No. XDB 26030304. The authors would like to acknowledge Professor Wei Qi for providing the material of the present study, Professors Deng Tao and Tong Haowen for helpful comments and suggestions to improve the manuscript.参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 5]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 4]
[本文引用: 2]
[本文引用: 4]
[本文引用: 7]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 4]
DOIPMID [本文引用: 1]

Ice Age megafauna have long been known to be associated with global cooling during the Pleistocene, and their adaptations to cold environments, such as large body size, long hair, and snow-sweeping structures, are best exemplified by the woolly mammoths and woolly rhinos. These traits were assumed to have evolved as a response to the ice sheet expansion. We report a new Pliocene mammal assemblage from a high-altitude basin in the western Himalayas, including a primitive woolly rhino. These new Tibetan fossils suggest that some megaherbivores first evolved in Tibet before the beginning of the Ice Age. The cold winters in high Tibet served as a habituation ground for the megaherbivores, which became preadapted for the Ice Age, successfully expanding to the Eurasian mammoth steppe.
DOIURL [本文引用: 1]
DOI [本文引用: 1]
Many fossil Elaphurus have been discovered in East Asia and some in Central Asia since the discovery of the first fossil E. bifurcatus of the genus from Nihewan Basin in the 1920s. Some of them are well preserved, but some others are fragmental and it makes some identifications doubtful. Two Elaphurus were identified in recent study based on the unstudied material of fossil deer unearthed from Taijiaping, Tianzhen, Shanxi Province in North China. The taxonomy of Elaphurus was consequently discussed and it is recommended to classify the genus into four species with eight subspecies. Elaphurus davidianus predavidianus subsp. nov. distributed in North China and Taiwan in the Early Pleistocene. E. davidianus tamaensis is a geographic subspecies of the Early Pleistocene in Japan. E. d. menziesianus derived from E. d. predavidianus and distributed in North, Northeast and East China including Taiwan, as well as in Japan, from the Middle Pleistocene to the Holocene and E. d. davidianus restricted in North and East China probably since recent thousands years ago. E. bifurcatus bifurcatus and E. b. shikamai existed in the Early Pleistocene in North China and Japan respectively. The probable ancestor of E. bifurcatus is Elaphurus eleonorae from the Early Pleistocene of Kuruksay, Tadjikistan in Central Asia. The Early Pleistocene E. chinnaniensis chinnaniensis and E. c. lantianensis ranged in North China only and with dubious generic status.
[本文引用: 1]
[本文引用: 1]
[本文引用: 12]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 3]
DOIURL [本文引用: 1]
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 6]
[本文引用: 1]
[本文引用: 2]
[本文引用: 5]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 17]
[本文引用: 2]
[本文引用: 2]
DOIURL [本文引用: 16]
DOIURL [本文引用: 4]
[本文引用: 1]
[本文引用: 8]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOIURL [本文引用: 3]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}