 ,1,2,3,*, 李志恒1,2
,1,2,3,*, 李志恒1,2Reexamination of the oldest pigeon (Aves: Columbidae) from Asia: Columba congi from the Early Pleistocene of Zhoukoudian, Beijing, China
SHEN Wei1,2,3, Thomas A. STIDHAM,1,2,3,*, LI Zhi-Heng1,2通讯作者: *presbyornis@gmail.com
收稿日期:2020-12-7
| 基金资助: |
Corresponding authors: *presbyornis@gmail.com
Received:2020-12-7

摘要
丛氏原鸽(Columba congi)出土于周口店北京猿人遗址早更新世第12地点(~1.7 Ma)。丛氏原鸽共出土11件骨骼碎片,但是由于年代久远,仅遗存4件肱骨标本,且遗存标本的形态学特征并不支持最初的物种鉴定特征。对遗存的4件肱骨标本进行了重新描述和对比,其肱骨气窝腹侧外缘平直和背髁上突位于肱骨更远端的位置这两个特征或为作为灭绝鸽类物种的丛氏原鸽提供稳定的识别特征。丛氏原鸽是亚洲地区已知最古老的鸽属化石,推测其生存于温暖且潮湿的森林环境。对丛氏原鸽的研究或能为解决鸽属物种的古生物地理分布以及目前广泛分布的原鸽的起源等问题提供帮助。
关键词:
Abstract
Columba congi is an extinct species that was described as part of the Early Pleistocene (~1.7 Ma) fauna from locality 12 of the UNESCO Zhoukoudian or “Peking Man” site in Beijing, China. Only four partial humeri of the original type series of 11 bones can be located, and the features present in those specimens do not support the original diagnosis. However, our study and redescription shows that the straight and flat margin of the pneumotricipital fossa rim (in ventral view) and the relative distal position of the dorsal supracondylar tubercle may support the continued recognition of C. congi as a valid extinct species. Columba congi appears to be the oldest fossil of Columba in Asia, and it lived during a warmer and wetter period of time of the Pleistocene with a forested Zhoukoudian. Further study of pigeons from all localities at Zhoukoudian should help to resolve questions about pigeon biogeography and evolution, including possibly the time and center of origin of the globally distributedC. livia.
Keywords:
PDF (716KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
沈韦, Thomas A. STIDHAM, 李志恒. 亚洲最古老鸽子(Columbidae)物种有效性再检验:北京周口店早更新世丛氏原鸽(Columba congi). 古脊椎动物学报[J], 2021, 59(3): 245-256 DOI:10.19615/j.cnki.1000-3118.210304
SHEN Wei, Thomas A. STIDHAM, LI Zhi-Heng.
1 Introduction
Members of the avian genus Columba (Columbidae) commonly are referred to as pigeons or doves (largely depending on their relative body sizes), and they have represented messengers of love and symbols of peace. In addition, these birds are a common dietary resource in cultures around the world. The long history of interactions between hominins and species of Columba extends back tens of thousands of years with records that Neanderthals consumed C. livia 40000 years ago (Blasco et al., 2014), and archeological evidence demonstrating that C. livia has been common food item in the Fertile Crescent more than 10000 years ago (Shapiro and Domyan, 2013). Furthermore, in the formulation of Darwin’s theory of evolution through natural selection, variation in the domestic pigeon,C. livia, played an enlarged part (Baptista et al., 2009). Pigeons continue to have an over-sized role (as compared to many other birds) in fundamental studies of evolutionary processes, interrelationships between the genotype and phenotype, and even teaching basic avian anatomy (e.g., Baptista et al., 2009; Shapiro et al., 2013; Jones et al., 2019). Despite those significant contributions to people and science, basic questions about the interrelationships of the species of Columba and the timing and pattern of their evolution remain elusive.The genus Columba includes 35 extant (and two historically extinct) species found on all continents except Antarctica (Gill et al., 2020). The species of Columbidae are particularly rich in China with records of 31 species placed in seven genera, and in particular 10 of the 35 species of Columba (MacKinnon and Phillips, 2000). Among those species of Columba in China are the sister species of the Domestic Pigeon or Rock Dove (C. livia) and the Hill Pigeon (C. rupestris; Johnson et al., 2001; Soares et al., 2016), along with the high elevation Snow Pigeon (C. leuconata).
In contrast to the abundance of the extant doves and pigeons around the world, fossils of doves and pigeons are much less common. The oldest fossils of Columbidae are from the late Oligocene or Early Miocene of Australia (Worthy, 2012). While Europe has a rich record of pigeon fossils (Weesie, 1984), with a large number published from Italy (Pavia, 1999; Abbazzi et al., 2004; Sardella et al., 2005; Bedetti and Pavia, 2007; Petronio et al., 2008), fossil records are rare in Asia, with published fossil specimens from Mongolia, China, Israel, and Azerbaijan (Hou, 1982, 1993; Rich et al., 1986; Tyrberg, 1998; Martynovich, 2002; Watanabe et al., 2018). The only published fossils attributed to Columbidae in China are specimens of Streptopelia and Columba from the UNESCO Zhoukoudian “Peking Man” site caves in Beijing, which are probably best known for their abundant hominin fossil specimens (particularly “Peking Man”), stone age technologies, and mammalian fauna. However, Rich and coauthors (1986) reported the unpublished occurrence of Columba at a site in Hebei Province (China), but the location of those fossil specimens is unknown.
Hou (1993) published the only overview of the diverse Pleistocene bird fossils found at Zhoukoudian, and he placed the avian fossils into 13 orders and 31 families of birds, representing a great taxonomic, morphological, and ecological diversity. Columbidae is one of the most abundant groups of birds in the Zhoukoudian fossil sites, with more than 700 specimens from localities 1, 3, 4, 12, 15, and Upper Cave allocated to the extant species C. livia, Streptopelia chinensis, and S. orientalis, and the extinct C. congi (Hou, 1993). Those cave localities and their columbid fossils range in age from ~1.7 Ma to ~35.1-33.5 ka (Cheng et al., 1996; Li, 2018), with locality 12 as the oldest fauna with columbid fossils. Hou (1993) erected the taxon Columba congi as an extinct species of pigeon based on the assemblage of larger columbid fossils from locality 12 (occurring with specimens of the smaller Streptopelia). With an estimated age of 1.7 Ma for the locality 12 fauna, the fossils of C. congi are possibly the oldest specimens of Columba in Asia (Tyrberg, 1998) and certainly the oldest in eastern Asia.
In the years since its publication (Hou, 1993), C. congi has been mentioned rarely in the literature, and its validity as part of the Pleistocene Eurasian avifauna appears not to have been challenged (Tyrberg, 1998; Zelenkov, 2015). With the established sister relationship between C. livia and C. rupestris (which both occur in China, and Beijing in particular) and their estimated recent divergence within the Pleistocene (Soares et al., 2016), early fossils of Columba in China may provide a window into the evolutionary origin of these commonly studied extant species. Therefore, given the age and geographic occurrence of C. congi, its further study may provide insight into pigeon evolution and biogeography. Columba congi from locality 12 is unique in the series of Zhoukoudian cave deposits in that Hou (1993) identified all other specimens of Columba from the Zhoukoudian cave localities solely as C. livia, despite the current occurrence of both C. livia and C. rupestris in Beijing and around Zhoukoudian.
There are 27 cave localities collectively called Zhoukoudian. Locality 18 is thought to contain the earliest fauna dating back to 1.9 Ma (Cheng et al., 1996), and the Upper Cave (locality 26) is the youngest with an estimated age of 35.1-33.5 ka (Li, 2018). Of course, columbid fossils are not present in all cave faunas. Hou (1993) considered Columba congi to be a kind of small primitive pigeon, and locality 12, where all known specimens of this extinct species derive, is a pocket-shaped cave with a fauna from the middle Early Pleistocene, ~1.70 Ma (Cheng et al., 1996). The upper part of the deposit is composed of hard dark red sediments, and the lower part is made of sands and gravels (Teilhard de Chardin, 1938; Cheng, 2002). Cheng (1997) interpreted the mammalian fauna from locality 12 as deriving from a hot and humid dense forest. Representative animals from locality 12 include more than 20 species, including rhinoceroses (Rhinoceros sp.), burrowing zokors (Siphneus sp.), an extinct monkey (Procynocephalus wimani), saber-toothed cats (Machairodus sp.), and hyraxes (Postschizotherium chardini) (Huang, 1960). In contrast to the rich mammalian diversity in locality 12 deposits, Columba congi and doves referable to Streptopelia sp. are the only birds reported (Hou, 1993). That mammalian fauna contrasts significantly with the extant fauna of northern China (lacking rhinos, monkeys, and hyraxes), and suggests a distinctly different environmental and climatic regime in the Early Pleistocene.
Institutional abbreviations BMNHC, Beijing Museum of Natural History Collection, Beijing; IOZ, Institute of Zoology, Chinese Academy of Sciences, Beijing; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing; OV, modern osteological specimen in the IVPP; V, fossil specimen in the IVPP.
Materials examined Modern skeletons: Columba livia, BMNHC 21449, IVPP OV 1363, OV 1735; C. rupestris, IOZ S031 and OV 859; Streptopelia chinensis, OV 1846 and OV 1908; S. orientalis, BMNHC 108613, OV 1883, and OV 1882. Fossil specimens: C. congi, V 6436. Measurements were taken with digital calipers and rounded to the nearest 0.1 mm.
Anatomical nomenclature Terminology for osteological features follows Baumel and Witmer (1993) with English equivalents of the Latin terms.
2 Systematic paleontology
Columbiformes Latham, 1790Columbidae Illiger, 1811
Columba Linnaeus, 1758
Columba congiHou, 1993
HolotypeHou (1993) designated the specimen number IVPP V 6436 as the holotype of C. congi. However, that single specimen number refers to 11 fragments of bones from locality 12 including parts of coracoids, ulnae, and humeri. Hou (1993) did not state how many specimens of the total of 11 are humeri, ulnae, or coracoids (or other parts of the skeleton), and he did not indicate which of these 11 specimens is the individual holotype specimen for the species.
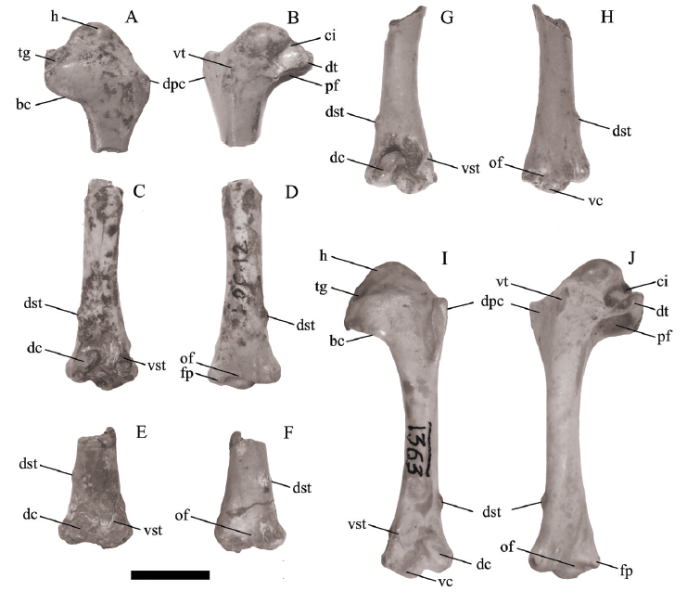
A search of the Zhoukoudian collections housed in the IVPP resulted in only four specimens total labeled as C. congi from locality 12 being located. Those four osteologically adult specimens (with smooth fully ossified bone surfaces) are one proximal left humerus and three distal right humeri representing a minimum of three individuals (Fig. 1). All are labeled as IVPP V 6436. The location of the other pigeon fossil materials mentioned and illustrated by Hou (1993) is unknown at present. None of the four specimens appear to be those that were illustrated by Hou (1993), and thus do not appear to strictly represent the holotype for the species. They can be treated as paratypes since they are part of the original type series designated byHou (1993). We designate the four humeri as IVPP V 6436.1-4 in order to facilitate communication about individual specimens and for future reference (Fig. 1; see description below).
Fig. 1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1Comparative images of the humerus of the extinct Columba congi (A-H) and Columba livia (I, J)
A, B. proximal end of left humerus of IVPP V 6436.1 in cranial (A) and caudal (B) views; C-H. distal end of right humerus of C. congi in cranial (C, E, G) and caudal (D, F, H) views: C, D. IVPP V 6436.2; E, F. V 6436.3; G, H. V 6436.4; I, J. left humerus of IVPP OV 1363 in cranial (I) and caudal (J) views
Abbreviations: bc. bicipital crest; ci. capital incisure; dc. dorsal condyle; dpc. deltopectoral crest; dst. dorsal supracondylar tubercle; dt. dorsal tubercle; fp. flexor process; h. humeral head; of. olecranon fossa; pf. pneumotricipital fossa; tg. transverse ligamental groove; vc. ventral condyle; vst. ventral supracondylar tubercle; vt. ventral tubercle. Scale bar = 10 mm
DiagnosisHou’s (1993:271) original diagnosis in English states, “A more primitive and smaller Columba, the wall thick and bone cavity small. The pneumatic fossa of the humerus not developed, no pneumatic foramen or very small. The humeral shaft flat and out border straight; the internal condyle of the distal end of the humerus not developed. The ulna shortly curved, smaller than that of C. livia.” No further description of the specimens is provided in English, but in Chinese he says (Hou, 1993:201), “the humeri have a small and curved crista deltopectoralis, shallow sulcus ligament transversus, small process supracondylaris dorsalis, large epicondyles, and undeveloped internal condyles. The only remaining coracoid is seemly straight and robust but not well preserved.”
The diagnoses (in Chinese and English) do not actually diagnose this species as separate from any other species of Columba, and many of the features are not present among the available fossils. The size of humerus of C. congi is smaller than C. rupestris and C. livia, and slightly larger than Streptopelia orientalis and S. chinensis (Table 1). That aspect of the original diagnosis appears correct. The statement about the wall thickness does not appear accurate. The bone thickness is not supported as a diagnostic feature since it is not significantly thicker in our analyses (see details below). It is possible that Hou’s (1993) interpretation of a thick-walled bone could have been based on the presence of medullary bone (of an egg-laying female), but that is merely speculation until the original specimens can be located. In addition, the humerus is a pneumatic bone in pigeons, and should not have been filled with red blood cell producing marrow (Schepelmann, 1990). The only proximal humerus among the four bones, IVPP V 6436.1, clearly preserves a pneumatic opening in the pneumotricpital fossa similar in size with that in extant skeletons; contradicting the statement about its absence in this extinct species. The preserved shafts of the humeri also do not differ visually from those of modern skeletons in their flatness and borders. We believe that Hou’s (1993) statement about the ‘internal condyle’ refers to the ventral condyle which is clearly present (except where broken off in V 6436.3) or possibly the ventral supracondyle which appears similar to other pigeons. We are not able to locate the ulna illustrated by Hou (1993), and cannot verify any potential diagnostic features in that skeletal element.
Table 1
Table 1Measurements of fossil and modern columbid humeri (mm)
| Specimen | Bp | Dtd | GBd | GL | SD | TW | |
|---|---|---|---|---|---|---|---|
| Columba congi | IVPP V 6436.1 L | 12.8 | 4.9 | 0.4 | |||
| IVPP V 6436.2 R | 9.5 | 10.2 | 4.8 | 0.4 | |||
| IVPP V 6436.3 R | 10.3 | 4.8 | 0.4 | ||||
| IVPP V 6436.4 R | 9.3 | 10.0 | 4.9 | 0.3 | |||
| Columba livia | BMNHC 21449 L | 14.5 | 10.5 | 46.7 | 5.4 | ||
| BMNHC 21449 R | 14.6 | 10.5 | 46.9 | 5.6 | |||
| IVPP OV 1735 L | 15.2 | 11.3 | 11.8 | 46.5 | 5.3 | ||
| IVPP OV 1735 R | 15.2 | 11.0 | 12.0 | 46.5 | 5.3 | ||
| IVPP OV 1363 L | 12.9 | 9.8 | 10.0 | 44.7 | 4.5 | ||
| IVPP OV 1363 R | 13.0 | 9.6 | 10.0 | 44.6 | 4.5 | ||
| Columba rupestris | IOZ S031 L | 12.7 | 10.0 | 10.3 | 43.5 | 5.2 | 0.4 |
| IOZ S031 R | 12.8 | 10.1 | 10.0 | 43.4 | 5.1 | 0.4 | |
| IVPP OV 859 L | 13.4 | 10.5 | 10.3 | 43.2 | 4.6 | ||
| Streptopelia orientalis | BMNHC 108613 L | 13.1 | 9.3 | 9.7 | 41.3 | 4.9 | |
| BMNHC 108613 R | 12.9 | 9.2 | 9.7 | 41.5 | 5.0 | ||
| IVPP OV 1883 L | 13.1 | 9.2 | 10.5 | 44.0 | 5.0 | ||
| IVPP OV 1883 R | 13.0 | 9.3 | 9.9 | 43.5 | 4.9 | ||
| IVPP OV 1882 L | 12.6 | 9.2 | 9.9 | 40.6 | 4.6 | ||
| IVPP OV 1882 R | 12.6 | 9.1 | 9.7 | 41.0 | 4.5 | ||
| Streptopelia chinensis | IVPP OV 1846 L | 10.9 | 7.9 | 8.9 | 34.4 | 3.8 | |
| IVPP OV 1846 R | 11.1 | 7.6 | 8.9 | 34.3 | 3.9 | ||
| IVPP OV 1908 L | 12.4 | 8.3 | 9.2 | 39.1 | 4.3 | ||
| IVPP OV 1908 R | 12.7 | 8.1 | 9.3 | 39.2 | 4.3 |
新窗口打开|下载CSV
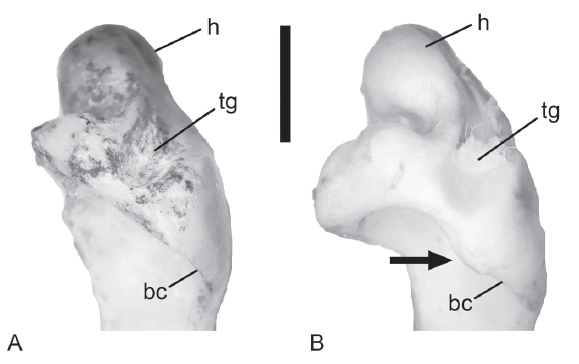
The deltopectoral crest of columbids and pteroclids has a very distinct pointed triangular outline that contrasts with the stated presence of a rounded crest in C. congi (Li et al., 2020). That statement also is contradicted by the shape presented in the photograph of the humerus of C. congi (Hou, 1993:fig. 11 on plate 6) and the morphology in IVPP V 6436.1. The transverse ligamental sulcus is not any smaller than the condition in extant comparative specimens (Fig. 2). The relative sizes of the dorsal and ventral supracondylar regions are consistent with the range of variation in extant skeletons. We could not locate the coracoid to verify any potential diagnostic features of the bone.
Fig. 2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Comparison of proximal pigeon humeri in ventral view illustrating a potential diagnostic feature
A. Columba congi (IVPP V 6436.1); B. C. rupestris (IVPP OV 869)
Arrow indicates the convex margin of the rim of the pneumotricipital fossa
Abbreviations: bc. bicipital crest; h. humeral head; tg. transverse ligamental groove. Scale bar=5 mm
Taken in total, we do not see any of the features listed by Hou (1993) except for size, as actually differentiating these fossils from any extant or extinct species. In part, that is because it appears that many of the features actually are not present in the specimens available. However, our close examination of the fossils reveals morphological characters that are diagnostic at a variety of taxonomic levels, including potentially that of species.
While the humerus exhibits many features common to Columbidae and Pteroclidae such as the large triangular deltopectoral crest, and a dorsal supracondylar tubercle located proximally, the humeral specimens from locality 12 display typical features of Columbidae including having an inset olecranon fossa, and they lack several features present in their close relatives Pteroclidae (like an indistinct olecranon fossa; Li et al., 2020). Comparisons with other taxa help to place the four fossil specimens in Columba using the following characteristics: humerus, proportionately more robust, shaft straighter, proximal and distal ends more expanded (Hume, 2011). Species of Streptopelia have an olecranon fossa that is a wider and shallower area, and these fossils also are distinctly larger than individuals of the dove species inStreptopelia (Table 1).
Tentative revised diagnosis: a couple of features appear to differ from other Pleistocene fossils and extant skeletons of Columba and may be diagnostic for this species. In ventral view, the caudodistal ventral margin or rim of the pneumotricipital fossa is straight in the proximal humerus of C. congi (Fig. 2), but the muscle origination scar on that rim projects distally (making the margin slightly convex) in Columba rupestris and C. livia. Though our sample sizes are small, this may be a diagnostic feature. The dorsal supracondylar tubercle also is positioned more distally in all distal humeri fossils as compared to C. livia and C. rupestris (see description below; Table 1).
The shared absence of these features in C. livia and C. rupestris could suggest that they represent synapomorphies of those species and a phylogenetic position of C. congi outside of the sister relationship between those extant species hypothesized based on molecular data (Johnson et al., 2001; Soares et al., 2016). Conversely, they could be plesiomorphies and indicate a derived character state in C. congi. We are unable to differentiate between the near identical humeri of C. rupestris and C. livia, but other portions of their skeletons exhibit variable characters (like the presence or absence of lateral trabeculae on the sternum). In addition, the great age of the C. congi fossils as compared to the other cave sites at Zhoukoudian (and their fossils ofColumba) also could point to a sister or even anagenetic type of evolutionary relationship with the current species in Beijing.
Locality Locality 12, Zhoukoudian, Fangshan District, Beijing, China; 200 m to the south of locality 1. The cave deposit in a vertical cylindrical pit four meters high with one meter diameter, the upper part of the deposit is hard dark red sediment, the lower part is coarse sand and gravel (Teilhard de Chardin, 1938), and the age of the deposit is middle Early Pleistocene, ~1.70 Ma (Cheng et al., 1996). Columba congi and Streptopelia sp. are the only avian fossils present in Locality 12. However, more than 20 mammalian species have been recorded, most of which are forest-linked mammals including bears (Ursus cf. U. angustidens), saber-toothed cats (Megantereon sp.), leopards (Panthera cf. P. pardus), and rhinoceroses (Rhinoceros sp.; Huang 1960 ; Cheng, 1997).
Description Currently, all specimens of Columba from locality 12 are designated as IVPP V 6436 including those illustrated and discussed by Hou (1993) and the four specimens described here. In order to accurately document the known specimens, we informally refer to each specimen with a number 1-4 after the specimen number. Thus, V 6436.1 refers to the proximal humerus. The three distal right humeri are V 6436.2, V 6436.3, and V 6436.4 (see Fig. 1 for illustrations of each specimen). V 6436.4 and V 6436.2 preserve the most of the humeral shaft (over half of the humeral length), and V 6436.3 is the least complete and missing the distal condyles. The dorsoventral width of the proximal end from the dorsal tubercle to the ventral edge of the bicipital crest is 12.8 mm. The dorsoventral width of the distal end is 10.2-10.3 mm ( Table 1). The smallest diameter of the humeral shaft is 4.8-4.9 mm. The thickness of the humeral shaft bone is 0.3-0.4 mm.
The proximal left humerus (IVPP V 6436.1) is missing the ventral tubercle and the apex of the deltopectoral crest, and the shaft is broken distal to the distal end of the deltopectoral crest. The deltopectoral crest is a large triangular projection with a short proximal side and a longer distal margin. The humeral head projects distinctly proximally and has a slight concavity on its cranial ventral aspect. The caudal face of the humeral head is flush with the adjacent distal humeral surface. The ventral tubercle is an ovoid scar that is flush with the adjacent humeral surface, except for its distal end that is very slightly raised. The capital incisure is a wide U-shaped concavity that does not undercut the head. The dorsal crus of the pneumotricipital fossa is short, and does not extend as far distal as the bicipital crest. A muscle scar (that widens distally) extends from the distal end of the dorsal crus to the broken end of the specimen. The distal end of the bicipital crest is distal to the apex of the deltopectoral crest, and in cranial view the distal margin of the bicipital crest is straight. The pneumotricipital fossa is partially walled, but has a central pneumatic opening (as in other specimens of Columba). In ventral view, the ventral rim of the pneumotricipital fossa has a straight margin that is inclined caudoproximal to craniodistal (Fig. 2). The caudal rim of the transverse ligamental sulcus adjacent to the capital incisure is a relatively thin ridge. The bicipital crest meets the shaft at an obtuse angle.
The distal end of the humerus is characterized by the ventral condyle extending distal to the dorsal condyle. The dorsal supracondylar tubercle is positioned far proximal relative to the distal end. The tubercle is located 9.3-9.5 mm proximal to the distal end in the fossil specimens (as compared to 9.6-11.3 mm in extant specimens;Table 1). The tubercle is not elongate proximodistally, and is positioned proximal to the proximal end of the brachial fossa. There is a shallow, arcuate fossa around the proximal end of the dorsal condyle that connects ventrally with the brachial fossa. The brachial fossa is a proximodistally elongate ovoid that is deepest ventrally. The proximal end of the dorsal condyle is at the same proximodistal level as the ventral supracondylar tubercle. The facet for the ventral collateral ligament has a very rounded subtriangular outline, is slightly convex on its surface, and faces slightly distally (not entirely cranially directed). The ventral supracondylar muscle scar for one of the pronator muscles is located proximal to the level of the ventral supracondylar tubercle. The area ventral to the ventral supracondylar tubercle and facet for the ventral collateral ligament is concave and directed somewhat cranioventrally. The scapulotricipitalis groove is present and extends proximally to about the same level as the proximal end of the dorsal condyle. The ventral margin of the groove projects more caudally than its dorsal margin. The olecranon fossa is a wide shallow area, but contains a distinctly inset concavity that cuts into the caudal side of the ventral condyle and extends towards the flexor process. The dorsal margin of the bone dorsal to the dorsal condyle is slightly concave.
3 Discussion
There has been a lack of reliable evidence for the origin of the crown group of Columbidae (Shapiro et al., 2002). In addition, the temporal and biogeographic origins of Columba and the ubiquitous C. livia are unknown at present. In part, this is the result of a low level of molecular sampling among the species of Columba for phylogenetic analysis (Johnson et al., 2001; Soares et al., 2016). With the exception of the globally distributed C. livia, the species of Columba are found exclusively in the Old World with the most species endemic to Africa and Asia (Gibbs et al., 2001). Given that the distribution of the sister species to C. livia (i.e. C. rupestris) is restricted to Asia, and a large number of species of Columba are endemic to Asia (14 species), it would appear possible, if not likely, that the cosmopolitan C. livia lineage originated in Asia. However, answering that question will require additional phylogenetic and paleontological data.Records of Columba and fossils referred to C. livia have been published from the Pliocene and Pleistocene across the Old World (Tyrberg, 1998). As yet, those identifications (and that of other fossils attributed to species of Columba) have not been tested via phylogenetic or other quantitative analyses. As we see in our study, the potential diagnostic features are few and not yet tested within a larger phylogenetic context of the clade Columba. While there are abundant pigeon fossils from the Pleistocene of Europe (Tyrberg, 1998), the origin of Columba remains elusive. Molecular clock estimation places the split between C. livia and C. rupestris late in the Pleistocene (Soares et al., 2016), and that is much younger than some of the oldest fossils attributed toC. livia (i.e. Pliocene). Either the molecular clock estimates are severely underestimating the time of divergence or the plethora of early fossils identified as C. livia belong to other lineages. We will be able to clearly address these questions only through greater sampling of species of Columba and with the construction of a morphological phylogenetic context for those species (extinct and extant). Our study of C. congi demonstrates both the problems and the potential outcomes from such a broader research approach. While we dismantled most of the features previously used to recognize C. congi as a separate species, we uncovered others that may withstand further sampling and testing and document C. congi as a valid species. In addition, the great age of these fossils along with their apparent morphological differences could, along with a study of the younger specimens identified as C. livia from Zhoukoudian, narrow hypotheses about the evolutionary origin of C. livia, and pigeons in Asia.
The sediments preserved in locality 12 represent a cave deposit. At present, there is not much sediment left at locality 12, and in recent years, the site was backfilled (Cheng, 2002). No additional specimens of C. congi will be found there. Columba congi is only known from locality 12 with its fauna representing a hot and humid climate, with abundant surface water and well-developed forests (Cheng, 1997). That hypothesized Early Pleistocene habitat contrasts with the preferences of the extant C. livia and C. rupestris which prefer more open environments (Goodwin et al., 1983). The occurrence of C. congi in a wet forest habitat along with an extinct monkey and leopards could indicate that its phylogenetic (and biogeographic) relationships might be closer to those columbid species present in southern China (and Asia) today. Hou (1993) identified C. congi only at locality 12 and designated all specimens from all other (younger) cave sites at Zhoukoudian asC. livia. The proposed difference in the inferred habitats of locality 12 may relate to the presence of C. congi during that time period alone, and its absence in other caves (and younger times) that represent more open habitats preferred by the extant species. A study of stable isotopes from the fossils possibly could help to resolve any habitat or ecological differences among the specimens of Columba present through the Pleistocene at Zhoukoudian. Furthermore, we recommend that future study examine the geologically younger fossil columbid material, particularly with reference to its potential placement within C. rupestris, the sister species of C. livia and a current resident species in Beijing.
Acknowledegments
We thank Song Gang and Liu Di for access to modern skeletons at Institute of Zoology, Chinese Academy of Sciences (Beijing) and the Beijing Museum of Natural History Collection. We also thank the reviewers for valuable comments on an earlier draft of the manuscript.参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOIPMID [本文引用: 1]

Feral Pigeons have colonised all corners of the Earth, having developed a close association with humans and their activities. The wild ancestor of the Feral Pigeon, the Rock Dove, is a species of rocky habitats, nesting typically on cliff ledges and at the entrance to large caves. This habit would have brought them into close contact with cave-dwelling humans, a relationship usually linked to the development of dwellings in the Neolithic. We show that the association between humans and Rock Doves is an ancient one with its roots in the Palaeolithic and predates the arrival of modern humans into Europe. At Gorham's Cave, Gibraltar, the Neanderthals exploited Rock Doves for food for a period of over 40 thousand years, the earliest evidence dating to at least 67 thousand years ago. We show that the exploitation was not casual or sporadic, having found repeated evidence of the practice in different, widely spaced, temporal contexts within the cave. Our results point to hitherto unappreciated capacities of the Neanderthals to exploit birds as food resources on a regular basis. More so, they were practising it long before the arrival of modern humans and had therefore invented it independently.
[本文引用: 3]
[本文引用: 4]
[本文引用: 2]
[本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 23]
[本文引用: 2]
DOIURL [本文引用: 1]
DOIURL [本文引用: 3]
DOIPMID [本文引用: 1]
The rock dove (or common pigeon),, is an important model organism in biological studies, including research focusing on head muscle anatomy, feeding kinematics, and cranial kinesis. However, no integrated computer-based biomechanical model of the pigeon head has yet been attempted. As an initial step towards achieving this goal, we present the first three-dimensional digital dissection of the pigeon head based on a contrast-enhanced computed tomographic dataset achieved using iodine potassium iodide as a staining agent. Our datasets enable us to visualize the skeletal and muscular anatomy, brain and cranial nerves, and major sense organs of the pigeon, including very small and fragile features, as well as maintaining the three-dimensional topology of anatomical structures. This work updates and supplements earlier anatomical work on this widely used laboratory organism. We resolve several key points of disagreement arising from previous descriptions of pigeon anatomy, including the precise arrangement of the external adductor muscles and their relationship to the posterior adductor. Examination of the eye muscles highlights differences between avian taxa and shows that pigeon eye muscles are more similar to those of a tinamou than they are to those of a house sparrow. Furthermore, we present our three-dimensional data as publicly accessible files for further research and education purposes. Digital dissection permits exceptional visualisation and will be a valuable resource for further investigations into the head anatomy of other bird species, as well as efforts to reconstruct soft tissues in fossil archosaurs.
DOIURL [本文引用: 2]
DOIURL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 2]
[本文引用: 1]
DOIURL [本文引用: 1]
PMID [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
PMID [本文引用: 6]
Pigeons and doves (Columbiformes) are one of the oldest and most diverse extant lineages of birds. However, the nature and timing of the group's evolutionary radiation remains poorly resolved, despite recent advances in DNA sequencing and assembly and the growing database of pigeon mitochondrial genomes. One challenge has been to generate comparative data from the large number of extinct pigeon lineages, some of which are morphologically unique and therefore difficult to place in a phylogenetic context. We used ancient DNA and next generation sequencing approaches to assemble complete mitochondrial genomes for eleven pigeons, including the extinct Ryukyu wood pigeon (Columba jouyi), the thick-billed ground dove (Alopecoenas salamonis), the spotted green pigeon (Caloenas maculata), the Rodrigues solitaire (Pezophaps solitaria), and the dodo (Raphus cucullatus). We used a Bayesian approach to infer the evolutionary relationships among 24 species of living and extinct pigeons and doves. Our analyses indicate that the earliest radiation of the Columbidae crown group most likely occurred during the Oligocene, with continued divergence of major clades into the Miocene, suggesting that diversification within the Columbidae occurred more recently than has been reported previously.
[本文引用: 1]
[本文引用: 5]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}