 ,1,2, 张鑫俊1,2,3, 程心4,5, 汪筱林1,2,3
,1,2, 张鑫俊1,2,3, 程心4,5, 汪筱林1,2,3A new pteranodontoid pterosaur forelimb from the upper Yixian Formation, with a revision of Yixianopterus jingangshanensis
JIANG Shun-Xing,1,2, ZHANG Xin-Jun1,2,3, CHENG Xin4,5, WANG Xiao-Lin1,2,3通讯作者: jiangshunxing@ivpp.ac.cn
收稿日期:2020-09-7网络出版日期:2021-04-20
| 基金资助: |
Corresponding authors: jiangshunxing@ivpp.ac.cn
Received:2020-09-7Online:2021-04-20

摘要
热河生物群的翼龙都发现于义县组和九佛堂组。金刚山层位于义县组上部,已经报道的翼龙包括最早发现的两件翼龙胚胎,两件古翼手龙超科的标本,以及一件存疑的金刚山义县翼龙。对这一层位发现的一件前肢标本进行了描述,并确定其属于无齿翼龙超科。对现存于本溪地质博物馆的金刚山义县翼龙的正型标本进行了观察,并修订其鉴定特征。无齿翼龙超科成员具有独有的特征组合为:牙齿垂直于咬合面,呈三角形且唇舌向扁,前两对更长更细;第二翼指骨长度约为第一翼指骨的93%。在热河生物群中,古翼手龙超科的标本主要发现于义县组,而古神翼龙超科主要发现于九佛堂组。包括这件新标本和金刚山义县翼龙在内的无齿翼龙超科在九佛堂组发现的略多于义县组,这与之前认为的翼龙分布情况不同。
关键词:
Abstract
Pterosaurs in the Jehol Biota have been found in the Yixian and Jiufotang formations. The Jingangshan bedding is in the upper part of the Yixian Formation. The first two pterosaur embryos ever discovered in the world, two archaeopterodactyloid specimens, and the questionable Yixianopterus jingangshanensis have been reported in previous literature. Here, we describe a forelimb from this horizon and confirm its phylogenetic position in the Pteranodontoidea. The holotype of Y. jingangshanensis, now housed at Benxi Geological Museum, has been examined. The diagnosis of this taxon has been revised without the consideration of the artificial parts as following, a pteranodontoid pterosaur with a distinguished combination of characters: triangular and labiolingually compressed teeth with the first two more slender and longer than the others; teeth vertical to the occlusal surface; the second wing phalanx about 93% the length of the first wing phalanx. In the Jehol Biota, archaeopterodactyloid specimens have been mainly discovered from the Yixian Formation, while tapejaroids are almost found from the Jiufotang Formation. Including the new forelimb and Y. jingangshanensis, the pteranodontoids from the Jiufotang Formation are slightly greater in number than those from the Yixian Formation in species and specimens, differing from the previous thoughts on the distribution.
Keywords:
PDF (2880KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
蒋顺兴, 张鑫俊, 程心, 汪筱林. 义县组上部一件无齿翼龙超科前肢及对金刚山义县翼龙的修订. 古脊椎动物学报[J], 2021, 59(2): 81-94 DOI:10.19615/j.cnki.1000-3118.201124
JIANG Shun-Xing, ZHANG Xin-Jun, CHENG Xin, WANG Xiao-Lin.
1 Introduction
Since the early 1990s, the Early Cretaceous terrestrial Jehol Biota has been famous around the world for producing feathered dinosaurs and many other exceptionally well-preserved vertebrate fossils (Zhou, 2014; Jiang et al., 2016; Wang et al., 2018, 2019; Zhang et al., 2019). It includes, from the oldest to the youngest, the Dabeigou, Yixian and Jiufotang formations, which respectively represent the early, middle, and late periods of this biota (Pan et al., 2013). Pterosaur is a representative group of the Jehol Biota, which includes more than 30 genera and species, all from the middle and late periods (Jiang et al., 2016; Lü, 2016; Wu et al., 2017; Zhang et al., 2019; Zhou et al., 2019; Hone et al., 2020). During the middle period, archaeopterodactyloids were dominating, mainly from the lower Yixian Formation, while in the late period, the dominant pterosaurs were ornithocheiroids (Pteranodontoidea + Tapejaroidea, sensu Kellner et al., 2019) (Wang and Zhou, 2006; Wang et al., 2014a; Wu et al., 2017).The Jingangshan bedding is in the upper part of the Yixian Formation (Smith et al., 1995; Wang et al., 1999), which has yielded some unique specimens. The first two pterosaur eggs with embryos ever discovered in the world were from this horizon (Ji et al., 2004; Wang and Zhou, 2004) as well as the earliest avian embryo (Zhou and Zhang, 2004). In addition, another three pterosaur specimens have been reported, including the holotype of Gladocephaloideus jingangshanensis (Lü et al., 2012), a referred specimen of this taxon with the only basihyal among pterosaurs (Jiang et al., 2020), and the holotype of Yixianopterus jingangshanensis (Lü et al., 2006a), which was previously considered as a questionable taxon (Jiang et al., 2020). In the previous phylogenetic results, Gladocephaloideus was located in the clade of the Ctenochasmatidae slightly varying in different studies (Lü et al., 2016; Vidovic and Martill, 2017). Yixianopterus jingangshanensis was tentatively assigned to the Lonchodectidae (Lü et al., 2006a). Recently, a taxonomic review of the Lonchodectidae has been done without the consideration of Y. jingangshanensis (Averianov, 2020). The brief description and figures of Y. jingangshanensis only provided limited information in the first publication (Lü et al., 2006a). Meanwhile, the Jinzhou Museum of Paleontology (JZMP), where the holotype should have been housed is closed. In August of 2020, two of the authors (J. S. & Z. X.) had an opportunity to examine specimens in the Benxi Geological Museum and found the holotype of Y. jingangshanensis. Here, we describe a pterosaur forelimb representing a pteranodontoid pterosaur, with a revision of the taxonomy of Y. jingangshanensis.
2 Material and methods
The new specimen (IVPP V 26616) has an incomplete right forelimb, housed at the Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences (IVPP). The material was collected from a local farmer at the beginning of the 21st century and was prepared by a professional technician in the IVPP. It was observed under the Carl Zeiss Stereomicroscope Stemi 508. Non-invasive observations were made using three-dimensional images of the outer and inner structures of the regions of interest. However, microtomography was not suitable in this case due to the shape of the specimen, which is composed of two broad and relatively thin plates (ca. 35×35×2 cm3). Micro X-ray computed laminography (micro-CL) was conducted in the Key Laboratory of Vertebrate Evolution and Human Origins, IVPP. Image segmentation and visualization were performed using the software VGStudio version 2.2 and Mimix version 17.0.The holotype of Yixianopterus jingangshanensis was reported to have been housed at a private museum, the Jinzhou Museum of Paleontology (Lü et al., 2006a), which has been closed since 2019. The holotype has been on exhibition in the Benxi Geological Museum for more than ten years under the previous number, JZMP-V-12. One of the museum staff is the collector of this specimen, and he told the authors how he found and “prepared” this fossil. He moved the anterior part of the upper and lower jaws into a new position and took some postcranial elements from the same body to make “a complete skull”, and a new piece of shale was used to “complete” the whole specimen. This specimen was observed under a microscopy and measured with calipers.
Phylogenetic analysis was performed on T.N.T. (Tree Analysis Using New Technology), version 1.5 for Windows (Goloboff and Catalano, 2016). The data matrix of Kellner et al. (2019) was used in this analysis. All characters were unordered and given the same weight. Ornithosuchus woodwardi was considered the primary outgroup. Analysis was performed using the traditional search, with the TBR algorithm, with 1000 random addition sequence replicates. Branches were collapsed when their minimum length was equal to zero. As some replications overflowed after the first round, traditional search was again employed, this time on trees from RAM. The command “best” was run to obtain the most parsimonious trees (MPTs).
3 Systematic paleontology
Pterosauria Kaup, 1834Pterodactyloidea Plieninger, 1901
Pteranodontoidea Kellner, 1996
Pteranodontoidea indet.
Material IVPP V 26166, an incomplete right forelimb in black color and articulately preserved in a hard, grey slab, association with fish, ephemerid, and plant fragments (Figs. 1-3), housed at the IVPP.
Fig. 1
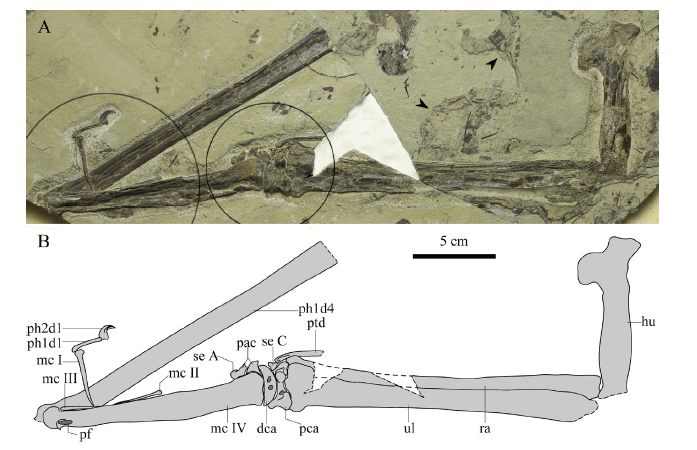 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1Photo (A) and line drawing (B) of IVPP V 26166 from the upper Yixian Formation
Two circles show the regions using the method of micro-CL, and two arrows show the different layers exposed in this specimen Abbreviations: dca. distal syncarpal; hu. humerus; mc I-IV. metacarpal I-IV; pac. preaxial carpal; pca. proximal syncarpal; pf. pneumatic foramen; ph1-2d1. first or second phalanx of manual digit I; ph1d4. first phalanx of manual digit IV; ptd. pteroid; ra. radius; se A. sesamoid A; se C. sesamoid C; ul. ulna
Fig. 2
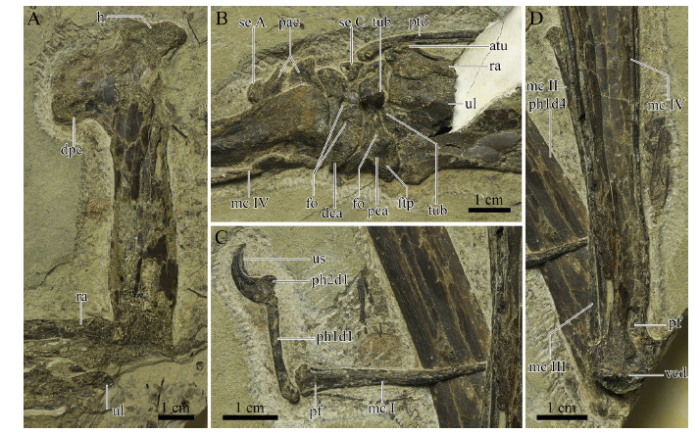 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Close-up of IVPP V 26166 from the upper Yixian Formation
A. The right humerus in medial view; B. The carpal region in ventral view; C. The first metacarpal and manual digit in ventral view; D. metacarpals II-IV Abbreviations: atu. anterior tubercle; dpc. deltopectoral crest; fo. foramen; ftp. flexor tendon process; h. head; tub. tuberculum; us. ungual sheath; vcd. ventral condyle; and the others see the caption of
Fig. 3
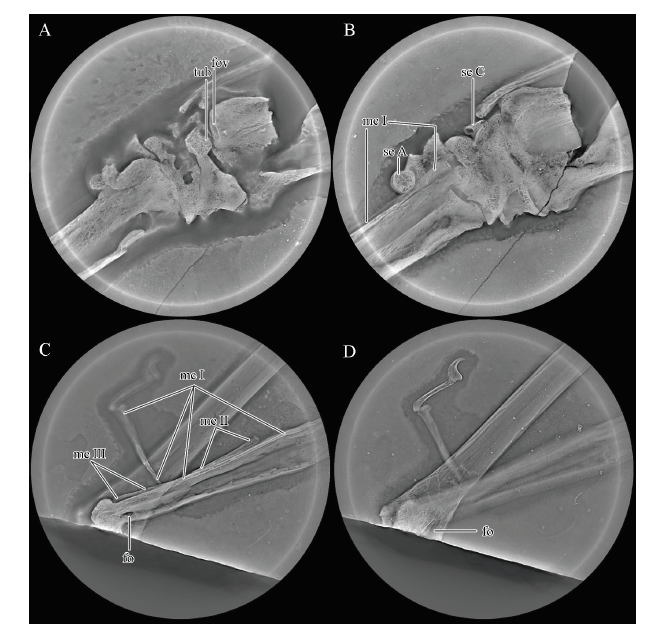 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3Micro-CL images of IVPP V 26166 from the upper Yixian Formation
A, B. the carpal region, same as the region in the right circle of
Abbreviations: fov. fovea; and the others see the captions of Figs. 1, 2
Locality and horizon Jingangshan, Yixian, Liaoning, China; Jingangshan bedding, the upper Yixian Formation, Lower Cretaceous.
Description and comparison The right humerus is straight in medial view (Fig. 2A). The exact length is undetermined because of the crushing of both ends, however, its estimated length is 101.6 mm (Table 1). The shaft of the humerus has no constriction. The head only has an outline, showing a dorsal inclination with the main shaft. The deltopectoral crest is warped and located distally, with an apparent constriction at the base. This warped deltopectoral crest is a pteranodontoid feature, also present in Chinese pteranodontoids, such as Hamipterus, Ikrandraco, and istiodactylids (Wang et al., 2005, 2014b, c; Andres and Ji, 2006; Hone et al., 2020). The length of this crest along the shaft is 25.38 mm, occupying a quarter of the humerus length. The ulnar crest and the distal end are unpreserved, and also no foramina can be detected on the preserved surface.
Table 1
Table 1Measurements of all elements in IVPP V 26166 (mm)
| Length | Mid-width | |
|---|---|---|
| humerus | 98.11a) | 16.55 |
| 101.60b) | ||
| radius | 190.43 | 6.21 |
| ulna | 189.42a) | 12.93 |
| carpals (proximal and distal) | 11.12 (in the middle) | 25.69 |
| metacarpal I | 134.92b)(100.44+34.48) | - |
| metacarpal II | 42.06 | - |
| metacarpal III | 29.67b) | - |
| metacarpal IV | 134.97 | 15.05 |
| 20.29 (max); 9.64 (min) | ||
| pteroid | 30.66a) | - |
| first phalanx of manual digit I | 20.00 | 2.20 |
| second phalanx of manual digit I | 9.71 | - |
| first phalanx of manual digit IV | 204.09a) | 12.96 (min) |
新窗口打开|下载CSV
The ulna and radius are similar in length, and the mid-width of the ulna is slightly greater than twice that of the radius (Table 1), which is a common feature in pteranodontoids (Kellner, 2003; Andres et al., 2014). The proximal end of the ulna is unpreserved, and the distal end is expanded as wide as the proximal syncarpal. A tuberculum is present at the distal end of the ulna (Fig. 2B), which is quite smaller than that of Pteranodon (Bennett, 2001) and Anhanguera araripensis (Wellnhofer, 1985). The ulnar fovea articulated with the tuberculum of the syncarpal, detected in the micro-CL image (Fig. 3A), is unexposed, which differs from the condition in which the fovea is offset ventrally in Dsungaripterus (Bennett, 2001). Due to the absence of the proximal end of the ulna, the proximal end of the radius is exposed, but not expanding as much as in Dsungaripterus or Pteranodon (Young, 1973; Bennett, 2001). The distal end of the radius only has the crushed anterior tubercle exposed, while the rest is overlaid by the ulna.
The pteroid, which is missing the distal part, is incomplete (Fig. 2B). It bends slightly in the proximal part. The diameters of preserved parts are nearly the same. There is probably an articular surface in the proximal end, and it formed a ridge during taphonomy, which is similar to the condition in Anhanguera araripensis (Wellnhofer, 1985). A small triangular element is present between the anterior parts of proximal and distal syncarpals (Fig. 2B), which is probably Sesamoid C as in Pteranodon (Bennett, 2001).
The proximal and distal syncarpals are fused respectively (Fig. 2B). The proximal syncarpal is exposed in ventral view. The flexor tendon process is developed, such as in Pteranodon, Dsungaripterus, Noripterus, and A. araripensis (Young, 1973; Wellnhofer, 1985; Bennett, 2001). A foramen is present between the flexor tendon process and the ventral margin of the ventral articular surface for distal syncarpal, where a small groove was formed in Dsungaripterus, Noripterus, and Pteranodon (Young, 1964, 1973; Bennett, 2001). The tuberculum of the proximal syncarpal is eminent, which is higher than the ventral surface in the flattened preserved specimen.
The ventral surface of the distal syncarpal is flat and crescentic. There are two foramina on the ventral surface: the anterior one is the same as in Dsungaripterus and A. araripensis, but the posterior one is formed by two smaller foramina (Young, 1973; Wellnhofer, 1985). These foramina are absent in Noripterus and Pteranodon (Young, 1973; Bennett, 2001).
The preaxial carpal still articulates with the facet of the distal syncarpal. The preaxial carpal is partially overlaid by metacarpal IV and broken in the middle. It is slightly elongated, differing from the triangular shape in Noripterus, A. piscator, Pteranodon, or Hamipterus (Young, 1973; Kellner and Tomida, 2000; Bennett, 2001; Wang et al., 2014b). The round end is part of the outline of Sesamoid A, and its suture with the preaxial carpal is only visible in the micro-CL image (Fig. 3B)
Using the method of micro-CL scanning, all four metacarpals can be observed. metacarpal IV, the wing metacarpal, is more robust than the others, which is a common feature in pterosaurs. Metacarpals II and III are reduced to less than a third the length of metacarpal IV (Table 1), and they taper proximally to a point (Fig. 3C), indicating the inarticulation with the distal syncarpal. Due to the connection with manual digit I, the slender bone vertical to metacarpal IV is interpreted as the distal part of metacarpal I (Fig. 2C, D), whose proximal part is overlapped by metacarpal IV. Metacarpal I has a flat facet and a small foramen lying on the ventral surface near the distal end (Fig. 2C), as well as in Pteranodon (Bennett, 2001). In the micro-CL images, it has a flat proximal articulation with the distal syncarpal (Fig. 3B). Metacarpals I-III still connect with the distal syncarpal in non-pterodactyloids and less advanced pterodactyloids (Bennett, 2001; Andres et al., 2014). In Anhanguera, Ikrandraco and istiodactylids, one of metacarpals I-III (probably metacarpal I) articulates with the distal syncarpal, while none of metacarpals I-III articulate with the distal syncarpal in Pteranodon or azhdarchids (Andres et al., 2014). Metacarpal II is next to the middle part of metacarpal IV (Fig. 2D), and it rotated 180° during taphonomy. Its distal end has a convex facet. Metacarpal III is in its anatomical position, exposing only its shaft (Fig. 2D).
Metacarpal IV is preserved in ventral view. It has an expanded proximal end, tapering posteriorly. The ventral condyle has a small breakage, and a large pneumatic foramen lies at the base of the condyle on the posterior surface (Fig. 2D), which is also present in Anhanguera piscator, Pteranodon, and Tapejara (Kellner and Tomida, 2000; Bennett, 2001; Eck et al., 2011), but absent in Dsungaripterus, Noripterus, and Ikrandraco (Young, 1964, 1973; Wang et al., 2014c). The metacarpal IV/humerus ratio is 1.33 (Table 1), slightly larger than that of Nurhachius ignaciobritoi, Longchengpterus, Ikrandraco and Istiodactylus sinensis (Wang et al., 2005, 2006, 2014c; Andres and Ji, 2006).
The first manual digit still articulates with metacarpal I (Fig. 2C). It has two phalanges: The proximal part of the first phalanx has a slight bend, and the second is a large, curved ungual. There is a black impression surrounding the distal end of the ungual, which is interpreted as the ungual sheath (Fig. 2C). The first wing phalanx is partially exposed in dorsal view, and the layer containing the distal part of the phalanx lost during the collecting, where the layer below exposed (see the arrows in Fig. 1). Although the proximal end is overlaid by metacarpal IV, with the help of the micro-CL, a large pneumatic foramen on the ventral side was possible to observe (Fig. 3D), which is also present in Haopterus, Nurhachius ignaciobritoi, Longchengpterus, Hamipterus, Ikrandraco, Anhanguera piscator and Pteranodon (Kellner and Tomida, 2000; Bennett, 2001; Wang and Lü, 2001; Wang et al., 2005, 2006, 2014b, c), but absent in Dsungaripterus and Noripterus (Young, 1964, 1973). The shaft of the first wing phalanx has a similar width. Although the first wing phalanx is incomplete, it is still more than twice the length of the humerus (Table 1), which is a feature only present in non-pterodactyloid Rhamphorhynchus and derived pterodactyloids (Andres et al., 2014).
4 Discussion
This new material can be assigned to the Pterodactyloidea because of the elongated wing metacarpal and the reduction of metacarpals II and III (Bennett, 2001; Andres et al., 2014; Kellner et al., 2019). The warped deltopectoral crest is a typical character of the Pteranodontoidea (Andres et al., 2014; Kellner et al., 2019), indicating a determined pteranodontoid pterosaur discovered in the upper Yixian Formation.By the observation of the holotype of Yixianopterus jingangshanensis, one of the authors (J. S.) confirmed the artificial parts of this specimen (see Fig. 4A), as well as some artificial bone surface. Each metacarpal IV is incomplete in the holotype, and the preserved length of the longer one is 118.43 mm, at least 53.0% of the first wing phalanx length, indicating that it is a monofenestratan pterosaur (Kellner et al., 2019). The mid-widths of ulna and radius were the same as noted in the previous publication (Lü et al., 2006a), which has never been reported in other monofenestratans. Based on the new measurements, the mid-widths of ulna and radius are 12.19 and 6.72 mm, respectively. Hence, the ulna/radius mid-width ratio is 1.8, which is a common feature in monofenestratans. The foot, not mentioned previously, is nearly completely preserved (Fig. 4C), and metatarsal III is 28.19 mm long. The tibia is not complete, and the preserved length is 102.63 mm. Therefore, the metatarsal III/tibia length ratio is less than 27.5%, which is a typical feature of the Ornithocheiroidea (Andres et al., 2014; Kellner et al., 2019). The teeth of Y. jingangshanensis are triangular and labiolingually compressed (Fig. 4B), from which it can be inferred that Y. jingangshanensis can be assigned to the Pteranodontoidea (Kellner et al., 2019). The first two teeth are slightly more slender and longer than the others.
Fig. 4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 4The holotype of Yixianopterus jingangshanensis (JZMP-V-12) in Benxi Geological Museum
A. photo of the whole specimen. The two regions in grey color indicate the artificial parts, and the region in the red circle should have been located in the blue circle based on what the person who found this specimen recalls; B. close-up of the teeth; C. close-up of the foot
Abbreviations: fe. femur; ft. foot; mt I-IV. metatarsal I-IV; pd 1-4. pedal digit I-IV; ph1-4d4. first to fourth phalanx of manual digit IV; ti. tibia; and the others see the caption of
Lü et al. (2006b) proposed following diagnosis of Y. jingangshanensis: “a pterodactyloid pterosaur with 1) subequal sized, well-spaced teeth; 2) a relative short metacarpal IV; 3) lengths of wing phalanx 1 and wing phalanx 2 nearly equal; 4) length ratio of wing phalanx 1 to metacarpal IV = 2; 5) length ratio of metacarpal IV to ulna = 0.64.” Based on the new measurements, the first and second wing phalanges are 223.28 and 207.33 mm, respectively, and the length ratio of the second to first wing phalanges is 0.93, which is only similar to Ikrandraco (Wang et al., 2014c), but larger than those of other pteranodontoids (Lü et al., 2006a; Andres et al., 2014). Since metacarpal IV is incomplete, characters 2), 4), and 5) are invalid for this taxon. Regarding the first character, the tooth size has a slight variation, and the distances between successive teeth become larger posteriorly. Hence, the revised diagnosis of Y. jingangshanensis is following: a pteranodontoid pterosaur with a distinguished combination of characters: triangular and labiolingually compressed teeth with the first two more slender and longer than the others; teeth vertical to the occlusal surface; the second wing phalanx about 93% the length of the first wing phalanx.
Two pteranodontoid specimens have been collected from the Jingangshan bedding, the upper Yixian Formation, the new specimen and the holotype of Y. jingangshanensis. However, there are few elements that can be compared between them. The only difference that can be confirmed is the ulna/radius mid-width ratio, which is more than two in the new specimen and less than two in Y. jingangshanensis. These two specimens are regarded as arising from different layers due to the different colors of the shales and bones, and these two specimens are tentatively treated as different taxa here. Five other pteranodontoid taxa have been published from Yixian Formation, including three boreopterid species, istiodactylid Luchibang xingzhe, and family-indetermined Haopterus gracilis(Wang and Lü, 2001; Jiang et al., 2014; Hone et al., 2020). The dentition of Y. jingangshanensis can distinguish this taxon from other members. The anterior teeth are extremely elongated in boreopterids (Jiang et al., 2014), a feature which is absent in Y. jingangshanensis. Typical diamond-shaped teeth of the Istiodactylidae in Luchibang (Hone et al., 2020) are also absent in Y. jingangshanensis. The tooth morphology is similar to that of Haopterus (Wang and Lü, 2001). However, the teeth are vertical to the occlusal surface in Y. jingangshanensis, while the teeth have an anterior inclination in Haopterus (Wang and Lü, 2001). Therefore, Y. jingangshanensis is still a valid taxon distinguished from other members from the Yixian Formation.
To access the detailed position of the new specimen and Y. jingangshanensis, a phylogenetic analysis was carried out. Many researchers have worked on the pterosaur phylogeny since the first publication in 1986 (Howse, 1986). Kellner (2003) and Unwin (2003) published their noteworthy work to evaluate the phylogenetic relationship of the Pterosauria. However, current researchers studying the pterosaur have yet to reach an agreement regarding the evolutionary tree. Despite the differences, the Pteranodontoidea is a relatively stable clade in most phylogenetic results (Bennett, 1994; Andres et al., 2014; Codorniú et al., 2017; Wang et al., 2017; Kellner et al., 2019), except for a small minority (Vidovic and Martill, 2017). Here, we have chosen the matrix of Kellner et al. (2019), and the new specimen and Y. jingangshanensis have been added (See supplementary document “Matrix.tnt”). The twenty-eight most parsimonious trees with a length of 361 steps have been obtained, and the strict consensus tree is shown in Fig. 5A. Despite the fact that the relationships among the Pteranodontoidea are unresolved, with the new specimen, Y. jingangshanensis, Haopterusgracilis, Mimodactylus libanensis, Linlongopterus jennyae, the Pteranodontia (sensu Kellner et al., 2019) and other two clades forming a polytomy in the strict consensus tree, confirming that the membership of the new specimen and Y. jingangshanensis in the Pteranodontoidea. The new specimen is an unstable taxon in this analysis, and the reduced consensus tree without it shows a further position of Y. jingangshanensis (Fig. 5B), which lies in the clade of the Istiodactyliformes (sensu Kellner et al., 2019), but the relationships among this clade have yet to be resolved.
Fig. 5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 5Phylogenetic results based on the matrix of Kellner et al., 2019
A. the strict consensus tree of 28 most parsimonious trees with a length of 361 steps; B. the reduced consensus tree without an unstable taxon (IVPP V 26166)
1. Ornithocheiroidea; 2. Pteranodontoidea; 3. Istiodactylidae
Pterosaurs are the main members of the Jehol Biota, and more than 30 taxa have been reported during the last three decades. The dominated taxa vary between Yixian and Jiufotang formations (Table 2) (Wang et al., 2020). One archaeopterodactyloid, Gladocephaloideus with two specimens, have been found in the Jingangshan bedding in the upper Yixian Formation (Lü et al., 2012; Jiang et al., 2020). More than 60% of archaeopterodactyloid taxa are from the lower-middle Yixian Formation, while nearly 30% of this clade are from the Jiufotang Formation, and there is a larger disparity in the number of specimens between these two formations. On the other hand, nearly all the taxa and tens of specimens of tapejaroids are from Jiufotang Formation, while there is only one species with two specimens from the Yixian Formation (Lü and Zhang, 2005; Lü et al., 2006b). The horizon bearing pteranodontoids is complicated. All three taxa of the Boreopteridae are from the lower Yixian Formation (Jiang et al., 2014), and nearly all taxa of the Istiodactylidae are from the Jiufotang Formation, except for the recently published Luchibang xingzhe (Hone et al., 2020). Other pteranodontoids were yielded from the lower Yixian Formation to the Jiufotang Formation, including two specimens from the Jingangshan bedding. This condition is slightly different from the previous mindset that pteranodontoids were mainly found from Jiufotang Formation (Wang et al., 2014a), or that toothed pterosaurs, including all Chinese pteranodontoids, were mainly found from the Yixian Formation (Lü, 2010). So far, pteranodontoid are reported slightly more from the Jiufotang Formation than from the Yixian Formation in species and specimen number.
Table 2
Table 2Comparison of pterosaur assemblages between Yixian and Jiufotang formations (in species)
| Anu | Arc | Pteranodontoidea | Tapejaroidea | |||||
|---|---|---|---|---|---|---|---|---|
| Bor | Ist | oth | Tap | Cha | ||||
| Jiufotang Formation | 0 | 3 | 0 | 6 | 4 | 5 | 3 | |
| upper Yixian Formation | 0 | 1 | 0 | 0 | 2 | 0 | 0 | |
| lower-middle Yixian Formation | 1 | 7 | 3 | 1 | 1 | 1 | 0 | |
新窗口打开|下载CSV
Acknowledgements
We thank Li Yutong (IVPP) for his preparation and Yin Pengfei (IVPP) for her help with the micro-CL scanning. We are grateful to Zhang Jun and Huo Honggang (Benxi Geological Museum) for access to pterosaur specimens under their care. We also thank Ji Shu’an and Zhou Changfu for their suggestions that improved this manuscript.Appendix can be found on the website of Vertebrata PalAsiatica (http://english.ivpp.cas.cn/sp/PalAsiatica/VP_llist/) in Vol. 59, Issue 2.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 2]
DOIURL [本文引用: 9]

The pterosaurs were a diverse group of Mesozoic flying reptiles that underwent a body plan reorganization, adaptive radiation, and replacement of earlier forms midway through their long history, resulting in the origin of the Pterodactyloidea, a highly specialized clade containing the largest flying organisms. The sudden appearance and large suite of morphological features of this group were suggested to be the result of it originating in terrestrial environments, where the pterosaur fossil record has traditionally been poor [1, 2], and its many features suggested to be adaptations to those environments [1, 2]. However, little evidence has been available to test this hypothesis, and it has not been supported by previous phylogenies or early pterodactyloid discoveries. We report here the earliest pterosaur with the diagnostic elongate metacarpus of the Pterodactyloidea, Kryptodrakon progenitor, gen. et sp. nov., from the terrestrial Middle-Upper Jurassic boundary of Northwest China. Phylogenetic analysis confirms this species as the basalmost pterodactyloid and reconstructs a terrestrial origin and a predominantly terrestrial history for the Pterodactyloidea. Phylogenetic comparative methods support this reconstruction by means of a significant correlation between wing shape and environment also found in modern flying vertebrates, indicating that pterosaurs lived in or were at least adapted to the environments in which they were preserved.
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 13]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
URL [本文引用: 5]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 3]
DOIURL [本文引用: 2]
DOIURL [本文引用: 3]
DOIURL [本文引用: 2]
[本文引用: 3]
DOIURL [本文引用: 12]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
[本文引用: 6]
[本文引用: 2]
DOIURL [本文引用: 2]
A new ctenochasmatoid pterosaur, Gladocephaloideus jingangshanensis gen. et sp. nov. from the Yixian Formation of western Laioning Province is erected based on a complete skull and partial postcranial skeleton. It is characterized by following features: about 50 total teeth with sharp tips; small nasoantorbital opening, occupying approximately 13% of the skull length: ratio of prenarial length to skull length approximately 0.63. The diagnoses of the Ctenochasmatoidea and Gallodactylidae are amended based on the new taxon. Gladocephaloideus jingangshanensis is the first gallodactylid pterosaur found in Asia. Its discovery not only provides much more osteological information about the Gallodactylidae but also indicates that the ctenochasmatoid pterosaurs were highly diverse in the Early Cretaceous. The filamentous structures preserved near the dorsal and posterior margins of the posterior portion of the skull and around the neck indicate that it had an epidermal covering and may have been a warm-blooded animal. Crown Copyright (C) 2011 Published by Elsevier Ltd.
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
Fossils from the "Jehol Biota" have been studied for nearly 90 years, and the term "Jehol Biota" has been extensively used and is well known today, largely thanks to the discovery of feathered dinosaurs, angiosperms, and numerous other exceptionally preserved fossils in the Lower Cretaceous of northeastern China. Ironically, however, what exactly the Jehol Biota represents and how it is defined has hardly been discussed. The lack of a precise definition of the Jehol Biota has hindered our understanding of such important issues as its palaeodiversity, its spatial and temporal distribution, and the pattern of radiation of the biota. In this paper, we propose that the Jehol Biota is defined by using a palaeoecological concept, and by combining ecological and taphonomic aspects, i.e., as organisms that lived in the Early Cretaceous volcanic-influenced environments of northeastern China, and were buried in lacustrine and rarely fluvial sediments, where most turned into exceptionally preserved fossils. The relationship between the Jehol Biota and the Jehol Group is also clarified. According to the revised definition, the Jehol Biota is so far only discovered in deposits of the Yixian and Jiufotang formations of western Liaoning, adjacent Inner Mongolia, and northern Hebei, and the Huajiying Formation of northern Hebei. Temporally it ranges from the Barremian to Aptian, i.e., for at least 10 Ma (130-120 Ma). (C) 2013 Elsevier Ltd.
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
DOIURL [本文引用: 1]
[本文引用: 2]
DOIURL [本文引用: 1]
DOIURL [本文引用: 4]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 3]
[本文引用: 2]
DOIURL [本文引用: 3]
DOIURL [本文引用: 4]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 4]
[本文引用: 2]
[本文引用: 3]
[本文引用: 8]
DOIURL [本文引用: 2]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}