 ,1,2,*, 易鸿宇,1,2,*, 卢静,1,2,*
,1,2,*, 易鸿宇,1,2,*, 卢静,1,2,*Reanalysis of Oculudentavis shows it is a lizard
LI Zhi-Heng1,2, WANG Wei3, HU Han4, WANG Min,1,2,*, YI Hong-Yu,1,2,*, LU Jing,1,2,*通讯作者: *wangmin@ivpp.ac.cn;yihongyu@ivpp.ac.cn;lujing@ivpp.ac.cn
第一联系人:
收稿日期:2020-08-17网络出版日期:2021-04-20
| 基金资助: |
Corresponding authors: *wangmin@ivpp.ac.cn;yihongyu@ivpp.ac.cn;lujing@ivpp.ac.cn
Received:2020-08-17Online:2021-04-20

摘要
最近在白垩纪晚期缅甸琥珀中报道了一件小型头骨化石,原论文作者将其鉴定为一蜂鸟大小的鸟翼类(Avialae)恐龙,并命名为宽娅眼齿鸟(Oculudentavis khaungraae)。基于这件标本,原论文提出了中生代恐龙(包括鸟翼类)体型演化和生态类型的种种假设。但原论文的插图和描述均显示这件头骨呈现出多个鳞龙类的典型形态学特征,这些特征很少在主龙类中出现。原论文先验地将标本鉴定为鸟翼类,继而进行仅包括鸟翼类作为内类群的系统发育分析。该分析的取样过于局限,不能对其系统位置归属提供有价值的参考。本文对宽娅眼齿鸟正型标本的计算机断层扫描原始数据重新进行了重建、渲染和分析,结果揭示了多个新的头骨形态学特征,包括侧生齿、开放的下颞孔以及头骨腭面着生有腭齿等,这些均为典型的鳞龙类/有鳞类特征,不支持其主龙类,遑论鸟翼类的系统位置归属。基于上述新的形态学发现,使用一双孔类(Diapsida)形态特征矩阵重新分析了眼齿鸟的系统演化位置。数据矩阵的分类单元涵盖了蜥蜴、鸟类和鳞龙形类与主龙形类中的主要支系。分析所得的严格合意树明确了眼齿鸟属于鳞龙形类中的有鳞类。因此,宽娅眼齿鸟的正型标本显示了足够丰富的形态学信息,完全能够指示眼齿鸟的系统分类位置应为有鳞类,而不是鸟翼类或非鸟恐龙。
关键词:
Abstract
The recent finding of a fossil entombed in a Late Cretaceous amber - Oculudentavis khaungraae - was claimed to represent a humming bird-sized dinosaur. Regardless of the intriguing evolutionary hypotheses about the bauplan of Mesozoic dinosaurs (including birds) posited therein, this enigmatic animal demonstrates various morphologies resembling lizards. If Oculudentavis was a bird, it challenges several fundamental morphological differences between Lepidosauria and Archosauria. Here we reanalyze the original computed tomography scan data of the holotype of Oculudentavis khaungraae (HPG-15-3). Morphological evidences demonstrated here highly contradict the avian or even archosaurian phylogenetic placement of the species. In contrast, our analysis revealed multiple skull morphologies of HPG-15-3 resembling those of squamates, including pleurodont marginal teeth, an open infratemporal fenestra, and the presence of palatal dentition. Based on these new morphological information, the phylogenetic position of Oculudentavis was analyzed in a data matrix sampling across the Diapsida. Taxon sampling of the data matrix included multiple species of lizards, birds, and major clades in Lepidosauromorpha and Archosauromorpha. In the strict consensus tree, Oculudentavis is nested within Squamata. These results show that morphology of the Oculudentavis khaungraae holotype supports a squamate rather than avian or dinosaurian affinity of the species.
Keywords:
PDF (3560KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
李志恒, 王维, 胡晗, 王敏, 易鸿宇, 卢静. 眼齿鸟再分析显示其蜥蜴属性. 古脊椎动物学报[J], 2021, 59(2): 95-105 DOI:10.19615/j.cnki.1000-3118.201020
LI Zhi-Heng, WANG Wei, HU Han, WANG Min, YI Hong-Yu, LU Jing.
1 Introduction
Oculudentavis khaungraae (HPG-15-3) was reported to be the smallest bird ever known, representing a previously unknown bauplan and ecology in Archosauria (Xing et al., 2020a). However, the holotype of O. khaungraae shows multiple features resembling squamates, including shape of the scleral ring and a possibly open infratemporal fenestra (Bolet et al., 2020; Li et al., 2020; Xing et al., 2020a). Four months later, the original paper (Xing et al., 2020a) was retracted for providing inaccurate phylogenetic hypothesis of Oculudentavis (Xing et al., 2020b). However, the authors claimed that their morphological descriptions of O. khaungraae (HPG-15-3) remained accurate.In an updated phylogenetic analysis sampling broadly in Diapsida, the authors argued HPG-15-3 was a bird (O’Connor et al., 2020). They further noted that the palatal dentition and infratemporal fenestra in the holotype (HPG-15-3) was poorly preserved, which may render the phylogenetic assessments of the species equivocal (O’Connor et al., 2020).
Here we reanalyzed the original computed tomography (CT) scan data and provide detailed morphological descriptions of the holotype of Oculudentavis khaungraae (HPG-15-3). Using three-dimensional reconstructions and CT slices, we reconstructed the complete orbit, jugal, and quadrate on the right side of the skull. In addition, we reconstructed the marginal and palatal dentition of HPG-15-3 to compare with the dentition of lizards and early birds. Based on the new morphological information, we analyzed the phylogenetic position of O. khaungraae in a data matrix sampling across Diapsida to test whether it belongs to Aves or Squamata.
As the indicated in the title, the current paper in Vertebrata palAsiatica represents a reevaluation of available CT-data from a withdrawn paper. No new Myanmar amber was involved in this study.
2 Material and methods
The original CT scan data was obtained upon request from the authors of original paper (Xing et al., 2020a). Two 3D format files (9.5 G in total) were combined into one and re-rendered in Drishti 2.7 (Hu et al., 2020). Scan data were analyzed in Avizo 9.0, VG Studio Max 3.4, and imaged in Adobe photoshop. Re-analysis 3D reconstruction files see Source data 1. More scanning settings, data information and original 3D reconstructions are available in Xing et al. (2020a).Phylogenetic position of Oculudentavis khaungraae was analyzed in a morphological data set sampling across Diapsida, including 70 taxa coded with 337 morphological characters. Taxon sampling follow Pritchard and Sues (2019) and O’Connor et al. (2020). Morphological coding of O. khaungraae was based on our observations of the CT data and a newly reconstructed 3D model of the holotype (HPG-15-3). The morphological matrix is provided as an executable TNT file in Appendix 1.
The matrix was analyzed in TNT (Goloboff et al., 2008) with Petrolacosaurus kansasensis as the outgroup. Implied weighting (k=12) was used in the maximum parsimony analysis of the phylogenetic relationship (Goloboff, 1993). This method is suitable for analyzing taxa with strong morphological convergences, such as Aves (Zhou and Zhang, 2002; Xu, 2018). The morphological matrix was subjected to two rounds of heuristic searches in TNT: 1) 1000 replications of TBR, keeping 10 trees in each replication, saving all trees to the RAM; 2) for all trees in the RAM, a second round of heuristic search was performed with 5 rounds of ratchet, 5 rounds of drifting, and 3 rounds of fusing. For the resulting strict consensus of the most parsimonious trees, branch support was calculated in TNT using 1000 replications of bootstrap (Felsenstein, 1985).
3 Systematic paleontology
Diapsida Laurenti, 1768Lepidosauria Haeckel, 1866
Squamata Oppel, 1811
Oculudentavis khaungraae Xing et al., 2020a
Holotype HPG-15-3 (Hupoge Amber Museum, Tengchong, China). A complete skull in amber.
Type locality and horizon Angbamo site, Kachin Province, northern Myanmar (Xing et al., 2020a). Cretaceous (Cenomanian) in age (Shi et al., 2012).
Revised diagnosis Oculudentavis khaungraae is a small squamate characterized with a unique combination of skeletal morphologies: an elongate snout with retract naris; posterior margin of maxilla terminating at the mid-level of the orbit; more than six posterior maxillary teeth located beneath the orbit; marginal teeth tightly packed with tooth count exceeding 30; plicidentine absent at the base of marginal teeth; palatal teeth present as a single row on the medial branch of the anterior process of pterygoid.
3.1 Morphological descriptions
The skull of O. khaungraae (HPG-15-3) is embedded in amber with all bones preserved in articulation. Re-analyses of the CT scan data of the holotype skull revealed anatomical features those are key to understanding the phylogenetic position of O. khaungraae.At the anterior tip of the snout, the premaxilla is fused with a long nasal process. The external naris opens dorsally on the snout, surrounded with the premaxilla, nasal, and maxilla (Fig. 1). The frontal appears to be fused to a single element with no longitudinal suture. In addition, the frontal demonstrates short ventral downgrowth that do not contact along the midline of the skull (Fig. 1J). This suggests the olfactory lobe of the brain was not fully encased in bony structures, which resembles non-burrowing lizards and differs from birds and dinosaurs (Gauthier et al., 1988; Benton, 2014). Posterior to the frontal, the parietal shows a possible parietal foramen near the dorsal apex of the skull roof (Fig. 1I).
Fig. 1
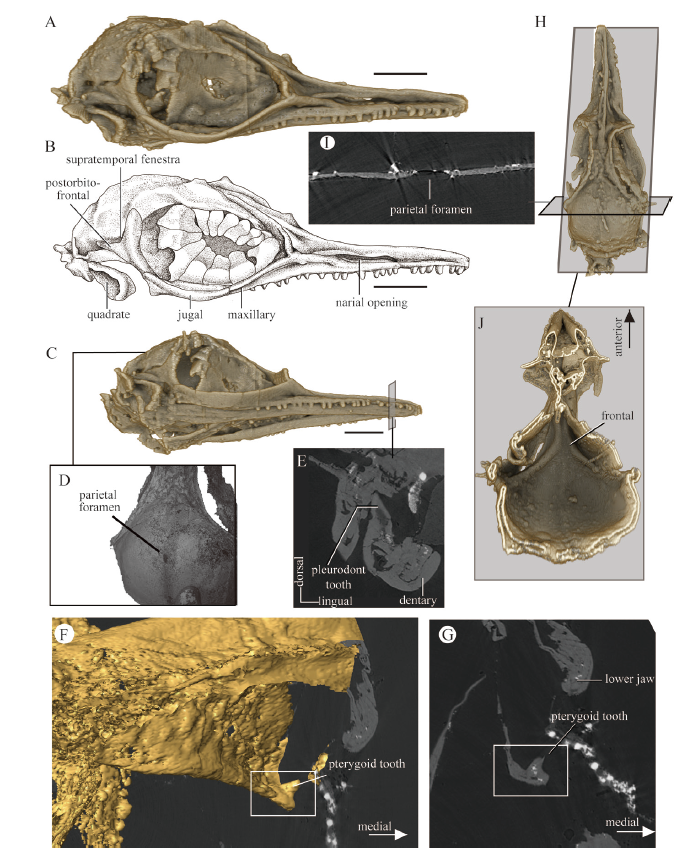 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1Cranial anatomy of the holotype of Oculudentavis khaungraae (HPG-15-3) based on re-analyses of the original propagation phase-contrast synchrotron radiation X-ray microtomography data
A, B. three-dimensional CT reconstruction (A) and line drawing (B) of the skull in right lateral view, showing the absence of quadratojugal; C. ventrolateral view; D. dorsal view, showing the parietal foramen; E. a two-dimensional CT slice through the dentary, showing a typical pleurodont tooth; F, G. pterygoid tooth shown in three-dimensional reconstruction (F) and in a coronal plane (G); H. dorsal view; I. a coronal CT slice through the skull roof showing the parietal foramen; J. ventral view, with the lower jaw and palate removed to show the ventral surface of the frontal. A-C, scale bars: 2 mm; D-J, not to scale
The maxilla contacts the premaxilla anteriorly and extends posteriorly to the mid-level of the orbit. There are more than 30 tightly packed maxillary teeth; posteriorly in the tooth row, at least six teeth are located beneath the orbit. The maxilla articulates with the jugal posteriorly. The jugal is complete on the right side of the skull but broken on the left side (Fig. 1A, B). The jugal has a smooth posterior margin, lacking contact with the quadrate. The quadratojugal is absent (Fig. 1A, B), which means the infratemporal fenestra is open in O. khaungraae - a condition shared with all squamates but not dinosaurs or birds (Estes et al., 1988; Gauthier et al., 2012).
In the palate of O. khaungraae, small conical teeth are present on the pterygoids (Figs. 1F, G; 2C, D), contrary to the observations in Xing et al. (2020a) and Bolet et al. (2020). Pterygoid teeth are observed on the right and left pterygoids, forming a single row on the medial branch of its anterior process (Fig. 2C, D).
Fig. 2
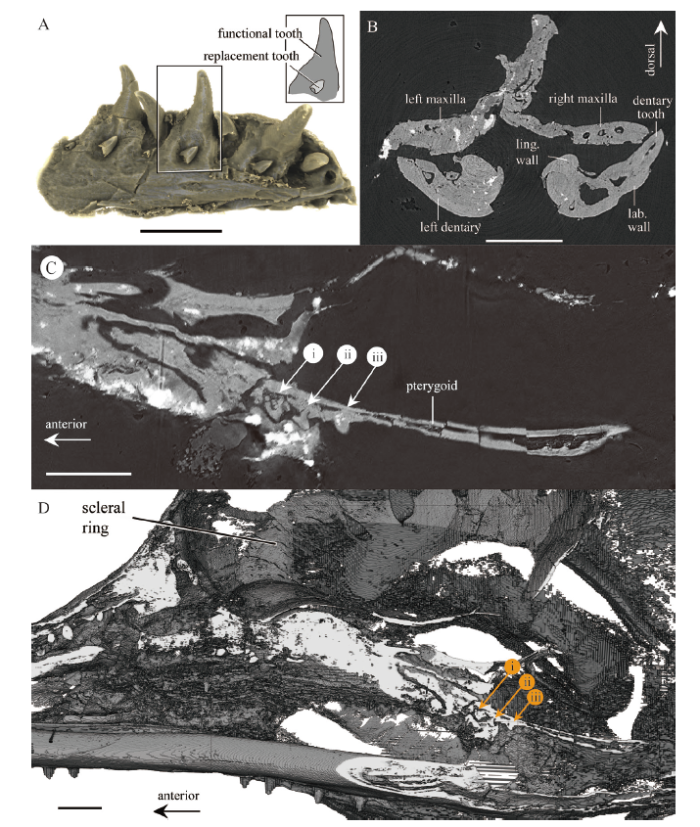 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Dentition of the holotype of Oculudentavis khaungraae (HPG-15-3)
A. three-dimensional reconstruction of the lower jaws showing patterns of tooth attachment and replacement in the dentary. The line drawing at top right corner depicts the relative position of the replacement tooth and functional tooth. B. transverse CT slice of the skull showing a cross-section through the mandibles. C, D. two dimensional (C) and three dimensional (D) sagittal CT slices of the skull through the right pterygoid, revealing three pterygoid teeth (i-iii) on the anterior process of pterygoid Abbreviations: lab. wall. labial wall of the groove for tooth attachment; ling. wall. lingual wall of the groove for tooth attachment. Scale bars: 2 mm
The marginal dentition of O. khaungraae is pleurodont (Fig. 2A, B) rather than “acrodont to pleurodont” suggested by Xing et al. (2020a). Among reptiles, acrodont teeth are ankylosed to the crest of jawbone (Estes et al., 1988), which is present in rhynchocephalians and a small group of lizards (Chamaeleonidae and Agamidae). In O. khaungraae, the maxillary and dentary teeth are ankylosed to the jaw with their labial side, demonstrating typical pleurodont condition (Fig. 2A, B). Furthermore, three-dimensional reconstruction of the dentary shows that replacement teeth develop posterolingual to the functional teeth (Fig. 2A). This is consistent with tooth replacement patterns in lizards (Gauthier et al., 2012).
3.2 Morphological comparisons with other squamates
The Squamata comprises four major clades of lizards: Iguania, Gekkota, Scincomorpha, and Anguimorpha (higher-taxon names sensu Gauthier et al., 2012). In addition, the Serpentes (snakes), Amphisbaenia, Dibamidae, and the extinct aquatic clade Mosasauria represent lineages with highly modified skeletons and controversial relationships with the four lizard clades (Gauthier et al., 2012; Pyron et al., 2013).Oculudentavis khaungraae demonstrates an elongate snout with a retracted naris, which differs from species of Iguania, Gekkota, Scincomorpha, Amphisbaenia, Dibamidae, and Serpentes. In Squamata, narial retraction is a shared derived feature of Varanoidea (a clade in Anguimorpha, sensu Gauthier et al., 2012), which is convergently developed in Mosasauria. However, O. khaungraaediffers from varanoids and mosasaurs in multiple skeletal features. The frontal is fused in O. khaungraae but paired in varanoids. The maxilla extends below the orbit in O. khaungraae but terminates anterior to the orbit in varanoids. Marginal teeth are tightly packed in O. khaungraae but loosely arranged in varanoids and mosasaurs. Plicidentine (enamel infoldings at the base of teeth) is absent in the marginal teeth of O. khaungraae but present in the teeth of varanoids and mosasaurs (Gauthier et al., 2012). A cemented section at the base of marginal teeth is a derived feature shared among mosasaurs (Caldwell, 2007), but this feature is absent in the teeth of O. khaungraae.
3.3 Phylogenetic position of Oculudentavis khaungraae
Phylogenetic analysis using implied weighting found two most parsimonious trees with 60.11 steps. In the strict consensus tree (Fig. 3), O. khaungraae is nested within Squamata, showing unresolved relationships with crown-group squamates and the fossil Huehuecuetzpalli mixtecus. Oculudentavis khaungraae shares with other squamates the following characteristics (character number - state number):Fig. 3
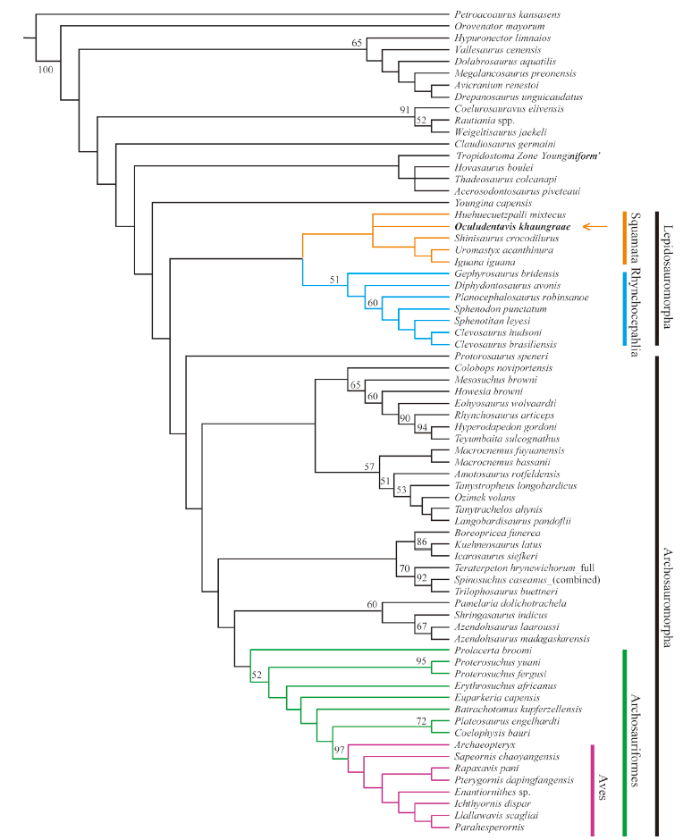 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3Phylogenetic position of Oculudentavis khaungraae
The data matrix follows Pritchard and Sues (2019) and O’Connor et al. (2020, preprint). In the strict consensus of the two most parsimonious trees, O. khaungraae is nested within Squamata. Tree length = 60.11, consistency index = 0.273, retention index = 0.674. Branches with bootstrap values above 50% are labeled
32-1, frontal exhibit gradual transverse expansion throughout anteroposterior length;
58-0, jugal lack posterior process;
85-0, pterygoid lack tooth on the lateral branch;
88-0, pterygoids do not contact along the midline of the skull;
89-1, long axis of the lateral process of pterygoid oriented anterolaterally.
Oculudentavis khaungraae shares with other lepidosauromorphs the following characteristics (character number - state number):
153-1, pleurodont dentition.
4 Discussion
Instead of demonstrating unequivocal synapomorphies of the Aves, O. khaungraae shows multiple characters that have rarely been found in any previously known Mesozoic birds or non-avian dinosaurs. One of the most controversial characters is the absence of an antorbital fenestra (Fig. 1A, B). Closure of the antorbital fenestra is widely distributed in various archosaurs such as crocodilians, ornithischian, and a few crown birds (e.g., shoebills) (Baumel and Witmer, 1993; Carpenter, 2011; Turner, 2015; Xing et al., 2017); however, presence of antorbital fenestrae is likely plesiomorphic among paravians, as all the known Cretaceous birds as well as their closest non-avian paravian relatives (e.g., dromaeosaurids, troodontids) preserve separate antorbital fenestrae (Rauhut, 2003; Norell et al., 2006; Turner et al., 2012; Wang and Hu, 2017).New reconstructions of the mandibles show O. khaungraae has pleurodont dentition. The dentary of O. khaungraae shows a shallow wall lingual to the tooth row, but this wall does not encase the base of dentary teeth (Fig. 2B). In Diapsida, pleurodont dentition occurs primarily in lepidosaurian-line reptiles, including early rhynchocephalians and the majority of lizards (Bertin et al., 2018). Dinosaurs and crocodylomorphs have thecodont teeth (Fig. 4) that develop in tooth sockets and grooves (Bertin et al., 2018). For instance, the Late Cretaceous ornithuromorph bird Hesperornis retain teeth in a groove formed from tooth sockets fusing together and set along the crest of the tooth-bearing bone (Dumont et al., 2016). The labial and lingual walls of the dental groove extend dorsally to the same height in Hesperornis (Dumont et al., 2016). However, the marginal teeth of O. khaungraae are set in a shallow groove whose lingual wall is distinctly lower than the labial wall (Fig. 2B). In Xing et al. (2020a), non-archosaurian dentition of O. khaungraae was interpreted as the result of miniaturization. To our best knowledge, there is no concrete evidence suggesting such drastic change of dentition in miniaturized archosaurs.
Fig. 4
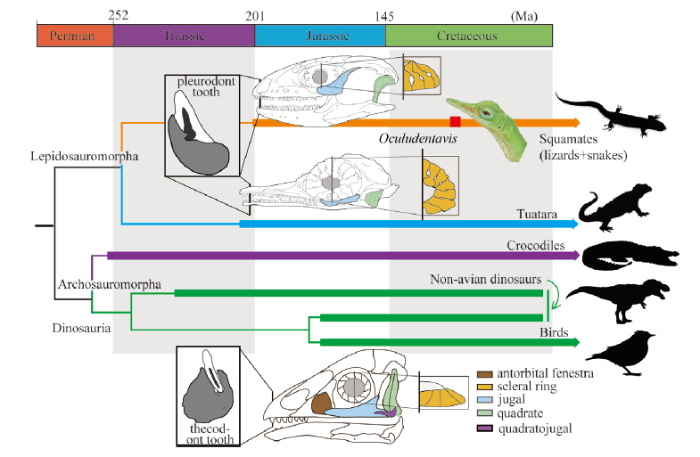 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 4Simplified reptile family tree
Illustrative drawings show comparisons of Oculudentavis khaungraae, a squamate (green lizard Lacerta bilineata) with a Cretaceous bird (Sapeornis chaoyangensis)
The phylogenetic analysis recovered O. khaungraae in Squamata with unresolved relationship with crown-group squamates and the fossil Huehuecuetzpalli mixtecus. The unique combination of long snout, retracted naris, and tightly arranged marginal teeth in O. khaungraae finds few parallels among squamates, which renders its phylogenetic position difficult to resolve in Squamata. In future analyses, adding crown squamates will help resolve the phylogenetic position of O. khaungraae within Squamata.
5 Conclusion
Oculudentavis means “eye-teeth bird”, yet neither the eyes (scleral ring) nor the teeth suggest this new species was a bird. Xing et al. (2020a) assigned this enigmatic animal to Aves based on superficial resemblances, such as the exterior contour of the dome-shaped cranium and slender rostrum (Xing et al., 2020a). However, our reanalysis of the CT data of O. khaungraae confirms multiple unambiguous characteristics supporting the squamate affinity of this animal, including the loss of the infratemporal bar, pleurodont marginal teeth, and presence of palatal teeth (Figs. 1-4). Our phylogenetic analysis with these new morphological information supports O. khaungraae as a squamate rather than bird or even archosaur.Acknowledgements
We thank Li Gang for providing the original CT scan data for reanalyzing, Yang Hong-Yu for drawing a life reconstruction of HPG-15-3, Guo Rui for drawing illustrations of HPG-15-3 and the green lizard. We thank colleagues at Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences for their great help during preparing and writing the manuscript. This work was supported by the Strategic Priority Research Program of Chinese Academy of Sciences Grant No. XDB26000000.Data Availability Statement
3D reconstruction files, re-analysis 3D reconstruction files of O. khaungraae can be downloaded through figshare: https://figshare.com/s/cb449dbbb1d93f6dfbda.The morphological matrix is provided as an executable TNT file in Appendix 1, which can be found on the website of Vertebrata PalAsiatica (http://english.ivpp.cas.cn/sp/PalAsiatica/VP_llist/) in Vol. 59, Issue 2.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 2]
URL [本文引用: 2]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
[本文引用: 2]
DOIPMID [本文引用: 1]

The recently-developed statistical method known as the "bootstrap" can be used to place confidence intervals on phylogenies. It involves resampling points from one's own data, with replacement, to create a series of bootstrap samples of the same size as the original data. Each of these is analyzed, and the variation among the resulting estimates taken to indicate the size of the error involved in making estimates from the original data. In the case of phylogenies, it is argued that the proper method of resampling is to keep all of the original species while sampling characters with replacement, under the assumption that the characters have been independently drawn by the systematist and have evolved independently. Majority-rule consensus trees can be used to construct a phylogeny showing all of the inferred monophyletic groups that occurred in a majority of the bootstrap samples. If a group shows up 95% of the time or more, the evidence for it is taken to be statistically significant. Existing computer programs can be used to analyze different bootstrap samples by using weights on the characters, the weight of a character being how many times it was drawn in bootstrap sampling. When all characters are perfectly compatible, as envisioned by Hennig, bootstrap sampling becomes unnecessary; the bootstrap method would show significant evidence for a group if it is defined by three or more characters.? 1985 The Society for the Study of Evolution.
[本文引用: 1]
DOIURL [本文引用: 6]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
DOIURL [本文引用: 1]
” doi: https://doi.org/10.1101/2020.06.12.147041, preprint
URL [本文引用: 4]
DOIURL [本文引用: 2]
DOIURL [本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
Amber from northern Myanmar has been commercially exploited for millennia, and it also preserves the most diverse palaeobiota among the worlds' seven major deposits of Cretaceous amber. Recent estimated ages vary from Albian to Cenomanian, based on palynology, an arnmonoid, and Mesozoic insect taxa preserved within the amber. The burmite-bearing rock is sedimentary and consists mainly of rounded lithic clasts (0.03 similar to 0.15 mm in diameter), with minor fragments of quartz and feldspar. Among the lithic clasts are mostly volcanic rocks. Zircons separated from the amber matrix form two groups: Group-1 zircons are overgrown and have variable CL patterns, experienced slight geological disturbances after they formed, and their Ion microprobe Pb-206/U-238 ages fall into a very narrow range of similar to 102 Ma- similar to 108 Ma; Group-II zircons are typical magmatic ones with rhythmically flat zones, inferred to be derived from volcanic rock clasts, and yielded a concordia Pb-206/U-238 age of 98.79 +/- 0.62 Ma. The dating on Group-1 zircons is only for their interiors, thus hiding what age excursion might come from the overgrowth. Considering the nearshore marine environment and 1-m thickness of the burmite-bearing sediments, and the syn- and post-eruption deposition of volcanic clasts, the age of 98.79 +/- 0.62 Ma therefore can be used as a maximum limit for the burmite (either at or after), establishing an earliest Cenomanian age for the fossilized inclusions. The age also indicates that volcanic eruption occurred at 98.79 +/- 0.62 Ma in the vicinity of the Hukawng Valley. (C) 2012 Elsevier Ltd.
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 12]
URL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}