1.Institute of Physics and Electronic Engineering, Qilu Normal University, Jinan 250013, China 2.School of Physics, State Key Laboratory of Crystal Materials, Shandong University, Jinan 250100, China
Fund Project:Project supported by the National Natural Science Foundation of China (Grant No. 11674195).
Received Date:27 March 2019
Accepted Date:15 June 2019
Available Online:01 September 2019
Published Online:05 September 2019
Abstract:Due to the exciton migration dynamics playing an important role in the photovoltaic process of organic solar cells, which are usually composed of polymer donor and fullerene (or non-fullerene) acceptor, in this paper we propose a new strategy to achieve the ultrafast exciton migration in polymers. Here, the effects of some nonuniform fields on the exciton migration dynamics in polymers are emphasized, such as the nonuniform electric field and the nonuniform polymer packing configuration field. Both of the two kinds of nonuniform fields can be intrinsically existent or modulated in an actual photovoltaic system. In this work, the nonuniform electric field and the nonuniform configuration field are assumed to be separately created by a confined charge and a linear polymer packing, therefore, their model Hamiltonian is established. In dynamical simulations of the exciton migration dynamics in polymers, an extended version of one-dimensional Su-Schrieffer-Heeger tight-binding model combined with a nonadiabatic evolution method is employed. It is found that the nonuniform electric field and the nonuniform configuration field both can drive exciton to an ultrafast migration process. Compared with the exciton migration speed dominated by the traditional F?rster or Dexter mechanism, the exciton migration speed dominated by the nonuniform electric field and that by the nonuniform configuration field can be increased by one and two orders of magnitude, respectively. In addition, the driving mechanisms of the two kinds of nonuniform fields for the exciton migration dynamics are separately clarified, where the corresponding driving forces are also quantitatively calculated. Finally, in view of the factors affecting the distributions of the two kinds of nonuniform fields (such as the distance d between confined charge and polymer, and the linear packing slope k between polymers), we discuss their effects on the exciton migration dynamics. It is found that the exciton migration in polymer can be apparently accelerated by shortening the distance d between confined charge and polymer, and there exists a critical value of d, beyond which the exciton will be dissociated into free charges in its migration process. For the linear packing slope k between polymers, we find that there exists an optimal value, at which the exciton has the highest migration speed in polymers. Keywords:polymers/ photovoltaic effect/ exciton migration
在实际的聚合物光伏体系内, 非均匀电场总是內禀存在的. 在其D/A界面附近, 聚合物链的取向更为无序, 各种缺陷也不可避免, 这造成D/A 界面附近的部分电荷被限制, 从而形成受限电荷. 另外, 由于D/A分子结构差异及它们的相对位置和取向不同, D/A界面附近电荷密度存在突变. 这些因素都会诱导D/A界面附近存在局域的非均匀电场[29,30]. 除了内禀因素外, 通过向聚合物光伏体系掺杂向电性添加剂(electrotropic additives)或具备极性的分子, 其内部的非均匀电场也可以人为的产生或调控[31,32]. 以往工作中, 通过引入线性梯度的电场已经对激子的超快输运和解离动力学过程进行了模拟, 并基于此解释了有机太阳能电池内观察到的超快电荷分离现象[33]. 本工作选取更接近实际情况的非均匀电场形式. 假定聚合物链的右端点附近存在一个受限正电荷(电荷量为$\left| e \right|$), 如图1所示, d为电荷与链右端点的距离. 根据点电荷的电场形式, 受限电荷诱导的电场沿聚合物链的分布为 图 1 受限正电荷 (电荷量为$\left| e \right|$)相对于聚合物链的位置, d为电荷与链右端点的距离; 曲线为d = 3 nm时电场强度沿分子链的分布, 负号表示电场的方向与分子链的正方向相反 Figure1. Schematic diagram about the position of a confined charge (q = $\left| e \right|$) relative to the polymer chain, d shows the distance between the charge and the right chain-end; the curve describes the distribution of the induced electric field E along the polymer chain with the case of d = 3 nm, where the minus sign means that the direction of the electric field is opposite to that of the chain.
${E_n} = - \frac{1}{{4{\text{π}} {\varepsilon _0}{\varepsilon _{\rm{r}}}}}\frac{{\left| e \right|}}{{r_n^2}},$
在本工作的模型中, 影响非均匀电场在聚合物链上分布的主要因素是受限电荷与链右端点的距离d. 图1中的曲线给出了d = 3 nm时, 开启后(t > tc)的电场沿分子链的分布. 可见, 电场强度沿分子链的方向迅速增加, 呈现出了明显的非均匀特点. 以产生在nc = 30 (nc为激子中心位置)附近的激子为初始态, 一旦开启以上形式的非均匀电场, 激子将经历演化. 如图2所示, 本文给出了激子在演化过程中分子链晶格结构随时间的变化, 即${y_n} = {( - 1)^n}(2{u_n} - {u_{n + 1}} - {u_{n - 1}})/4$. 可以发现, 在非均匀电场的驱动下, 激子自其初始位置沿分子链向右输运, 大约在t = 2000 fs (或2 ps)时, 激子到达分子链的右端并反弹. 就其总体过程而言, 激子在2 ps时间内输运的距离达到了5 nm, 即其平均输运速度高达2.5 nm/ps. 这一速度值比由F?rster或Dexter扩散机制主导的激子输运速度值(约0.1 nm/ps)提高了1个量级[16-19]. 可见, 非均匀电场能够驱动激子沿分子链超快输运. 图 2 激子在非均匀电场(d = 3 nm)驱动下沿聚合物链超快输运对应的晶格动力学演化, 其初始产生时的中心位置为nc = 30 Figure2. Time evolution about the lattice configuration of an exciton in a polymer chain driven by a nonuniform electric field with d = 3 nm, where the exciton is initially generated at a central position of nc = 30.
然而, 激子作为中性态为什么能够在电场的驱动下实现输运呢? 以往的研究表明[34,35]: 激子在均匀电场的作用下一般呈现极化的行为, 即激子内的电子和空穴在电场力的作用下不再完全重合, 而是沿分子链有一定程度的分离, 从而造成激子内出现等量异号的极化电荷. 图3的插图给出了t = 1000 fs时激子在电场下的极化电荷分布${q_n}(t) = e[{\rho _{n,n}}(t) - 1]$. 可见, 激子中心的右侧出现了极化的负电荷, 而其左侧则出现了极化的正电荷. 如果电场是均匀场, 正负极化电荷所处的电场强度相同, 那么激子整体所受的电场力为零, 激子无法输运. 但是, 本工作中所采用的电场为非均匀场, 极化的负电荷所处的电场强度要高于正电荷, 这将导致激子整体受到沿分子链向右的电场力, 这正是激子输运驱动力的来源, 即$F = \sum\nolimits_n {{q_n}(t){E_n}(t)} $. 随着激子沿分子链向右输运, 它感受到的电场强度迅速增加(如图1曲线所示), 这造成激子的极化程度显著增强. 一方面, 激子内极化的正负电荷量增多; 另一方面, 正负电荷所处的电场差异增强. 因此, 驱动力F随着时间或激子的输运将迅速增大. 定量的计算结果如图3所示, 与以上的分析一致. 图 3 非均匀电场诱导激子输运的驱动力F随时间的变化; 插图为t = 1000 fs时激子内极化的正电荷与负电荷的分布 Figure3. Variation of the driving force F as a function of the time. The inset presents the polarized positive charges and negative charges in the exciton at the time t = 1000 fs.
此处, 聚合物链内激子的输运速度可以通过调控非均匀电场沿分子链的分布进一步提高. 由(6)式可知, 受限电荷与链右端点的距离d是影响非均匀电场分布的重要因素. 图4给出了激子的中心位置nc在不同非均匀电场(通过改变d调控)驱动下随时间的演化. 很明显, 随着距离d的减小, 激子将更快地到达分子链的右端. 当d = 1 nm时, 激子中心大约经历1.5 ps就从其初始位置nc = 30到达链的右端, 并且其中心位置可以达到nc = 80, 随后激子将在链端反射呈现振荡的行为. 可见, 当受限电荷与链右端点的距离d减小至1 nm时, 激子的平均输运速度可增至约4 nm/ps. 至于不同d值时激子到达链端所呈现出的不同振荡行为, 这主要取决于不同情况下的激子输运速度. 激子输运速度越大, 激子的振荡行为越明显. 当d = 3 nm或d = 5 nm时, 激子到达链端也会振荡, 但振荡周期和幅度将分别变长和变弱. 另外, 需要补充两点: 一方面, 受限电荷与链右端点的距离d存在临界值, 当d小于此临界值时, 受限电荷在分子链右侧区域诱导的电场将足以使激子解离, 相关图像以往已有报道[34,35], 此处不再赘述; 另一方面, 在模型处理中, 本工作假定受限电荷正好处于分子链的延长线上, 这与实际情况可能不符. 当受限电荷处于任意位置时, 对激子输运起作用的电场实际为受限电荷诱导的电场沿分子链的分量. 这种情况下, 电场分量沿分子链的分布也是非均匀的, 以上图像同样适用. 图 4 不同非均匀电场(通过改变d调控)驱动下激子中心位置nc随时间的演化 Figure4. Time evolution of the exciton center nc along the polymer chain driven by different nonuniform electric fields, which can be modulated by changing the value of d.
23.2.非均匀构型场驱动的激子超快输运 -->
3.2.非均匀构型场驱动的激子超快输运
基于D/A结构的聚合物光伏体系内, 特别是D/A界面处, 由于侧链基团、空间结构及其他无序效应的影响, 聚合物分子之间通常会呈现非均匀的排列[36]. 比如考虑两条聚合物分子链, 在其耦合区域, 非均匀的排列将导致它们之间的相互作用也是非均匀的. 因此, 沿分子链方向必定存在与分子间的排列构型相对应的非均匀场, 本文称之为非均匀构型场. 为简单起见, 本工作构造了两条呈线性排列的分子链, 如图5所示. 每条分子链的总格点数均为N = 100, 沿分子链方向垂直最近邻格点间的距离dn呈线性减小的趋势, 即 图 5 线性排列构型的耦合双分子链, dn为分子链垂直最近邻格点间的距离 Figure5. Schematic diagram of two coupled polymer chains with a linear interchain packing configuration, where dn indicates the distance between the vertical-neighbor sites.
${d_n} = k \cdot \left( {N - n} \right) \cdot a + {d_N},$
其中, k代表分子链之间线性排列的斜率; a = 0.122 nm为晶格常数; dN为分子链右端点对应的垂直最近邻格点间距离, 取dN = 0.24 nm. 引入垂直最近邻格点间的电子跃迁项${t_ \bot }\left( n \right)$描述分子链之间的相互作用, 它的大小取决于dn, 见(12)式[37],
开启以上形式的非均匀构型场后, 激子将经历演化. 在以上模型中, 分子排列的非均匀构型场可通过改变分子链之间线性排列的斜率k进行调控. 图6给出了k = 0.03时激子在演化过程中分子链晶格结构${y_n}$随时间的变化. 发现: 在构型场开启之初(1—50 fs), 初始产生在第1条链上的激子由于链间相互作用迅速向第2条链扩展, 但保持其中心位置nc = 30不变; 当激子在两条链之间扩展至一定程度, 在非均匀构型场的驱动下, 激子自其初始位置沿分子链向右输运, 大约在t = 200 fs (或0.2 ps)时, 激子到达分子链的右端. 总体而言, 激子在0.2 ps时间内输运的距离达到了约6 nm, 即其平均输运速度高达30 nm/ps. 这一速度值不仅比F?rster或Dexter机制主导的激子输运速度提高了2个量级, 也比由前面非均匀电场驱动的激子输运速度提高了1个量级. 图 6 激子在线性链间构型场 (k = 0.03)驱动下在分子间扩展及沿分子链输运的晶格动力学演化, 激子初始产生在第1条分子链上, 中心位置为nc = 30 Figure6. Time evolution about the lattice configuration of an exciton in two coupled polymer chains driven by the nonuniform configuration field with a linear coefficient k = 0.03.
下面将深入分析激子在此线性构型场驱动下超快输运的物理机制. 以往的静态研究已表明分子链间的相互作用强度将会对激子的产生能有重要影响[12]. 具体来说, 激子的产生能会随着分子链间相互作用强度的增强而减小. 本文的模型处理中, 分子链间的相互作用强度${t_ \bot }{\rm{(}}n{\rm{)}}$是通过改变分子链间的距离dn来调控的. 图7中的插图给出了k = 0.03时, 激子的产生能$\Delta E$随分子链间距离dn的变化. 很明显, 链间距越小, 激子产生能越低, 这一能量梯度必将沿分子链方向诱导激子输运的驱动力F, 可以简单地通过$F = - \left( {\Delta {E_{n + {\rm{1}}}} - \Delta {E_n}} \right)/a$求出. 如图7所示, 本文给出了k = 0.03时, 线性构型场所诱导的激子驱动力沿分子链的分布. 一方面, 驱动力为正, 这将驱动激子沿分子链正方向输运. 另一方面, 驱动力沿着分子链的正方向逐渐增强, 这将导致激子沿分子链输运的速度逐渐加快, 这与图6的模拟结果是一致的. 特别是, 通过对比非均匀电场和构型场诱导的激子输运驱动力(图3和图7), 发现线性构型场诱导的驱动力要比前者高2个数量级, 这导致线性构型场驱动的激子输运速度更快. 图 7 线性构型场(k = 0.03)诱导的激子驱动力F沿分子链的分布, 插图为激子产生能$\Delta E$随链间距离dn的变化 Figure7. Distribution of the driving force F along polymer chains driven by a linear configuration with k = 0.03. The inset presents the dependence of the exciton creation energy $\Delta E$ upon the interchain distance dn.
根据前面的模型处理 (见(11)式), 耦合分子链线性构型场的分布可以通过改变其斜率k来调控, 也就是说, 激子沿分子链输运的速度必定与k的取值密切相关. 图8给出了不同k值时激子中心位置nc随时间的演化. 发现: 线性构型场驱动的激子输运速度可以通过改变其斜率k调控. 特别是, 存在最优化的斜率值k = 0.03, 此时激子输运速度最快. 当k < 0.03时, 由于沿分子链方向激子产生能的梯度趋于减小, 驱动力F必将减弱, 因此激子输运的速度将减慢. 极端的情况是分子链均匀排列, 即k = 0时, 激子的产生能在分子链各处均相等, 激子将保持不动. 当k > 0.03时, 由于模型处理中固定了链右端点的距离dN = 0.24 nm, 这导致分子链左侧区域的链间距过大, 这对激子产生能的影响将变小, 因此激子输运的速度也趋于减慢. 同样, 可以考虑极端情况, 即$k \to \infty $时, 模型体系左侧分子链间的距离趋于无限远, 第2条链对第1条链内产生的激子无任何影响, 激子也将保持其初始位置不动. 当然, 实际的分子排列构型一般不是线性的, 形式可能要复杂得多. 但是, 对于耦合的分子链, 其耦合区域的分子间距离总是存在局部的递减趋势, 这一变化必将诱导激子产生能沿分子链方向的梯度(或激子输运的驱动力), 从而驱动激子超快输运. 图 8 不同线性分子排列构型场(通过改变k调控)下激子中心位置nc随时间的演化 Figure8. Time evolution of the exciton center nc along polymer chains driven by different configuration fields, which can be modulated by changing the value of k.















 图 1 受限正电荷 (电荷量为
图 1 受限正电荷 (电荷量为






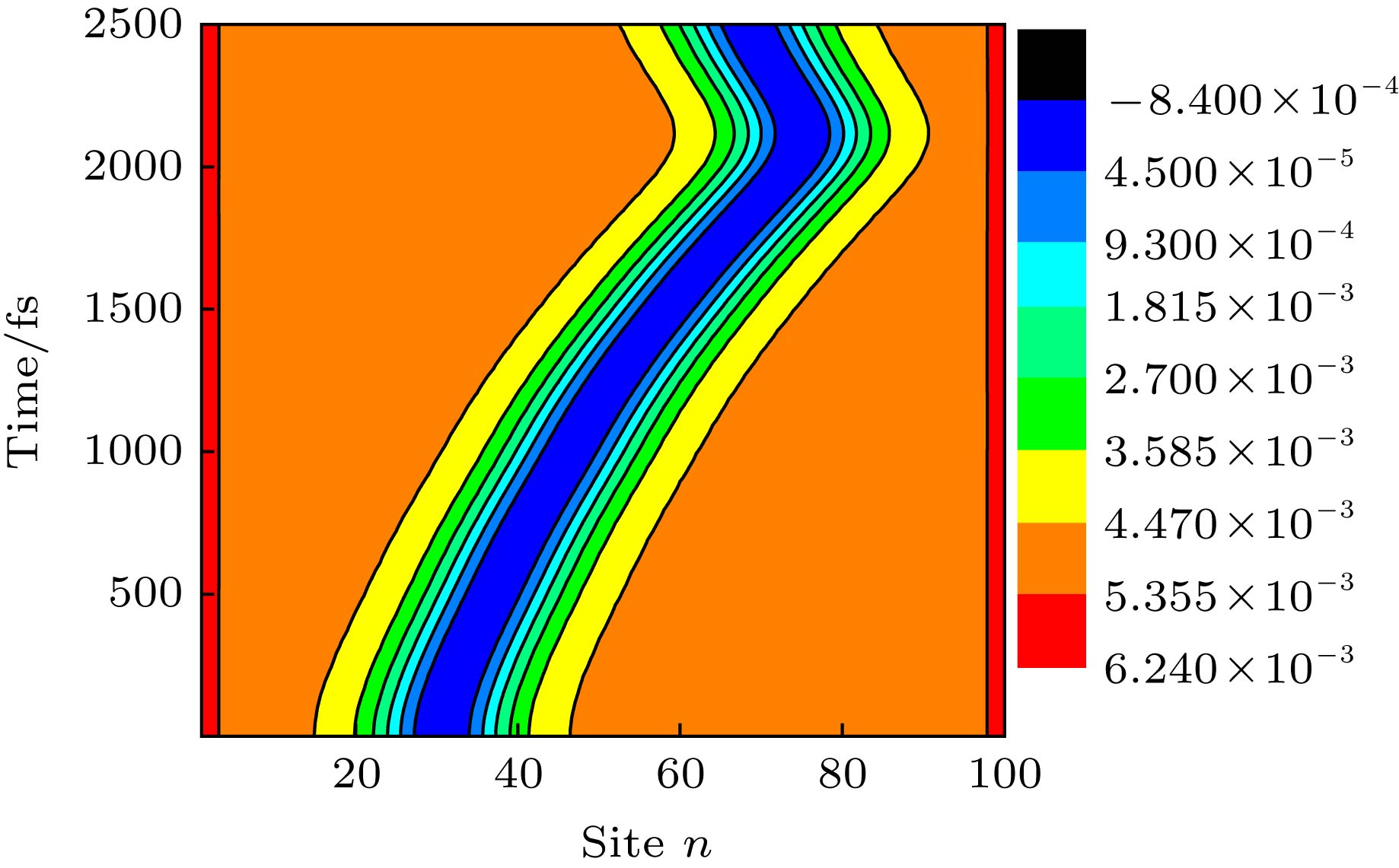 图 2 激子在非均匀电场(d = 3 nm)驱动下沿聚合物链超快输运对应的晶格动力学演化, 其初始产生时的中心位置为nc = 30
图 2 激子在非均匀电场(d = 3 nm)驱动下沿聚合物链超快输运对应的晶格动力学演化, 其初始产生时的中心位置为nc = 30

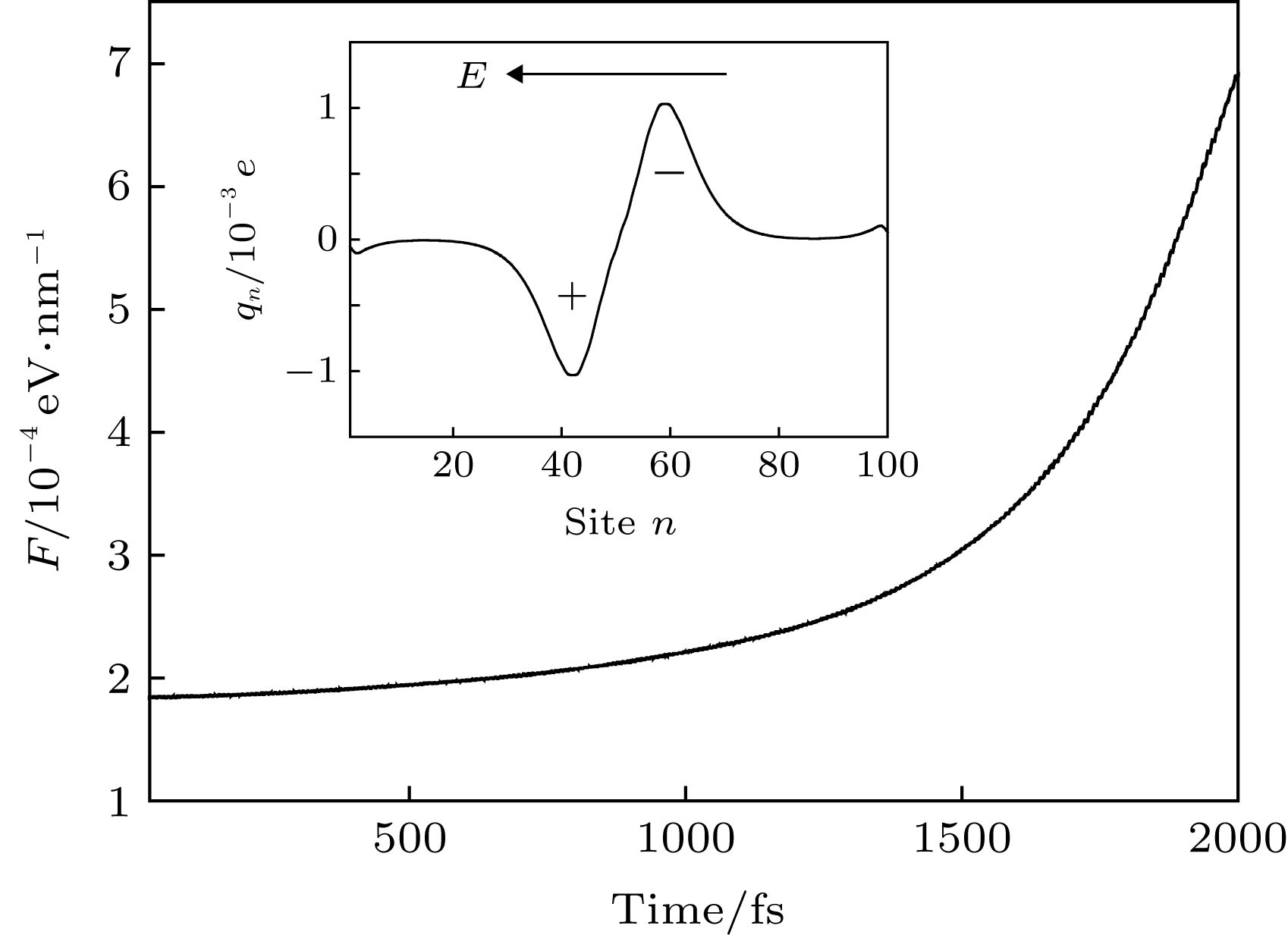 图 3 非均匀电场诱导激子输运的驱动力F随时间的变化; 插图为t = 1000 fs时激子内极化的正电荷与负电荷的分布
图 3 非均匀电场诱导激子输运的驱动力F随时间的变化; 插图为t = 1000 fs时激子内极化的正电荷与负电荷的分布 图 4 不同非均匀电场(通过改变d调控)驱动下激子中心位置nc随时间的演化
图 4 不同非均匀电场(通过改变d调控)驱动下激子中心位置nc随时间的演化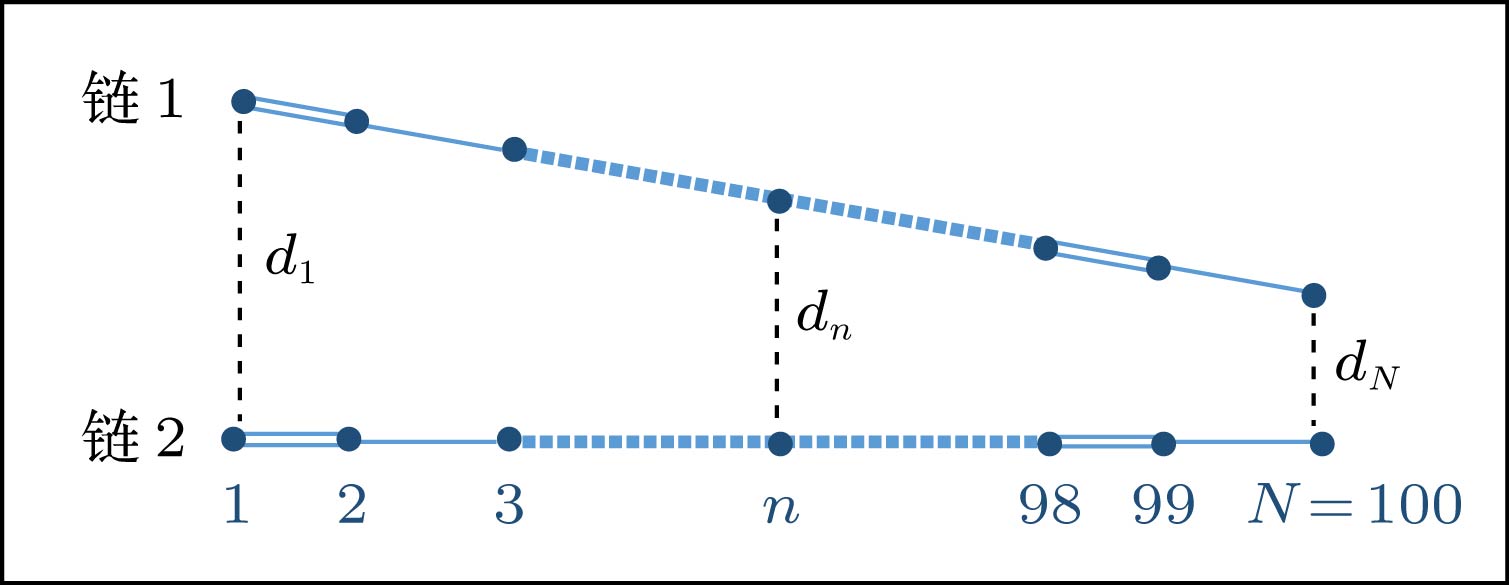 图 5 线性排列构型的耦合双分子链, dn为分子链垂直最近邻格点间的距离
图 5 线性排列构型的耦合双分子链, dn为分子链垂直最近邻格点间的距离


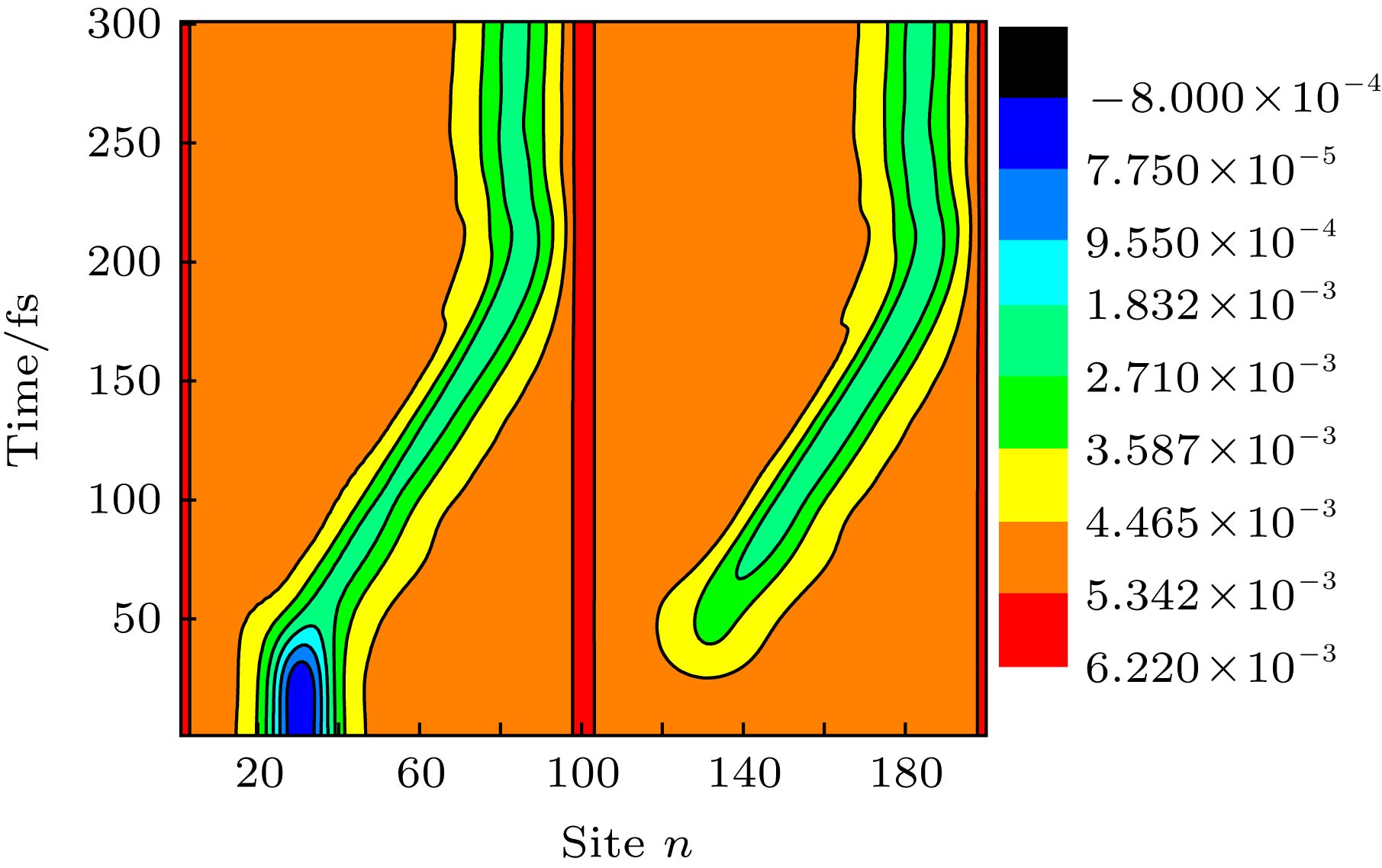 图 6 激子在线性链间构型场 (k = 0.03)驱动下在分子间扩展及沿分子链输运的晶格动力学演化, 激子初始产生在第1条分子链上, 中心位置为nc = 30
图 6 激子在线性链间构型场 (k = 0.03)驱动下在分子间扩展及沿分子链输运的晶格动力学演化, 激子初始产生在第1条分子链上, 中心位置为nc = 30


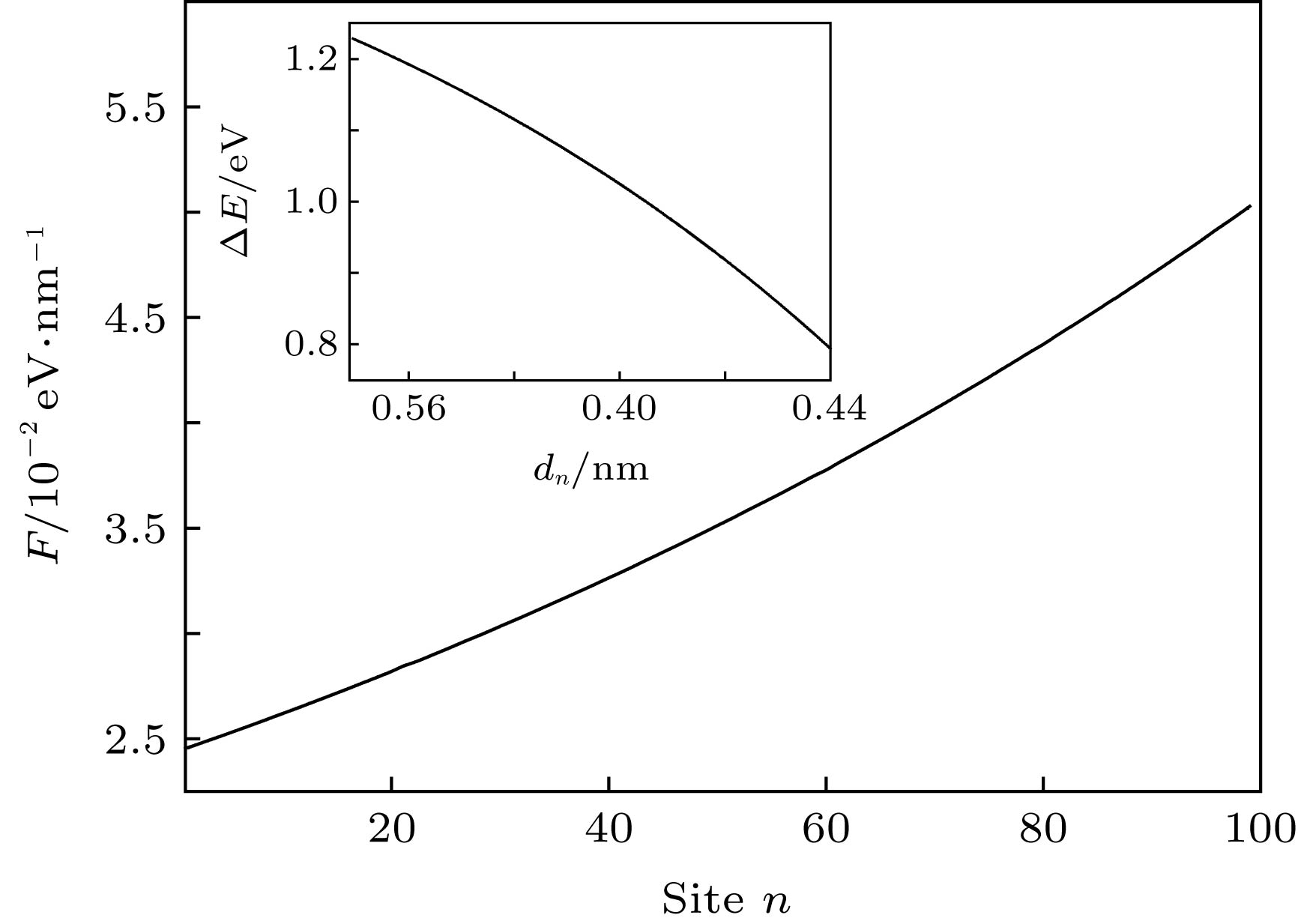 图 7 线性构型场(k = 0.03)诱导的激子驱动力F沿分子链的分布, 插图为激子产生能
图 7 线性构型场(k = 0.03)诱导的激子驱动力F沿分子链的分布, 插图为激子产生能


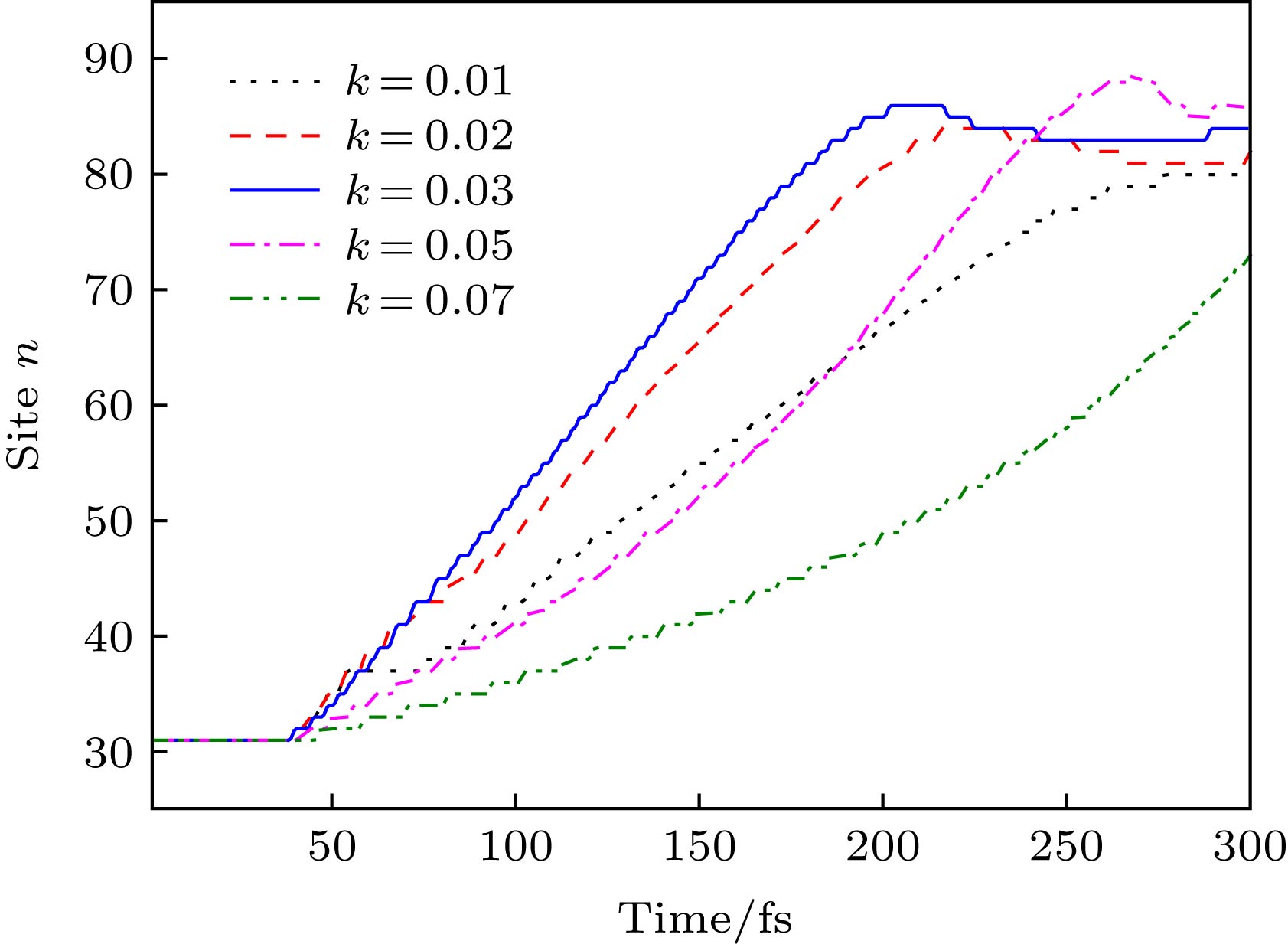 图 8 不同线性分子排列构型场(通过改变k调控)下激子中心位置nc随时间的演化
图 8 不同线性分子排列构型场(通过改变k调控)下激子中心位置nc随时间的演化