 ,首都儿科研究所遗传室,北京 100020
,首都儿科研究所遗传室,北京 100020The NMD escape mechanism and its application in disease therapy
Miaomiao Cheng, Yanyan Cao,Department of Medical Genetics, Capital Institute of Pediatrics, Beijing 100020, China通讯作者: 曹延延,博士,副研究员,研究方向:脊髓性肌萎缩症的分子机制。E-mail:caoyanyan@bjmu.deu.cn
编委: 卢大儒
收稿日期:2019-12-20修回日期:2020-02-25网络出版日期:2020-04-20
| 基金资助: |
Editorial board:
Received:2019-12-20Revised:2020-02-25Online:2020-04-20
| Fund supported: |
作者简介 About authors
程苗苗,硕士研究生,专业方向:儿科学。E-mail:

摘要
无义介导的mRNA降解(nonsense-mediated mRNA decay, NMD)是指在病理或正常生理情况下mRNA上出现了提前终止密码子(premature termination codon, PTC),从而导致mRNA降解。它是一种广泛存在的mRNA质量监控机制。近年来,在多种疾病中发现某些PTC并未触发NMD,这种现象被称为NMD逃逸(NMD escape),然而其确切机制尚不十分清楚。目前公认的两个学说为:(1) PTC通读,即蛋白的翻译可以顺利通过PTC直至正常的终止密码子,产生全长蛋白;(2)翻译的重新启动,即蛋白翻译在PTC下游的潜在起始点重新开始直至终止密码子,产生N端截短蛋白。目前,通过利用PTC通读,越来越多的药物或小分子已被成功用于无义变异相关疾病的治疗。本文主要综述了NMD逃逸的机制及其在疾病治疗中的应用和进展,以期为进一步了解NMD逃逸及其相关应用概况提供参考。
关键词:
Abstract
Nonsense-mediated mRNA decay (NMD) refers to the degradation of mRNA due to the presence of premature stop codon (PTC) on mRNA under pathological or physiological conditions. NMD is widely considered an mRNA-specific quality control process. Recently it was discovered that some PTCs do not trigger NMD in a variety of diseases - a process known as NMD escape; however, its exact mechanism remains unclear. At present, there are two widely accepted mechanistic hypotheses during NMD escape. The first is PTC read-through, in which protein translation undergoes PTC until the normal stop codon is encountered, producing a full-length protein. The second is translation reinitiation, in which protein translation recommences at the potential start codon downstream of PTC and terminates at the stop codon, producing an N-terminal truncated protein. Currently, an increasing number of drugs or small molecules that use PTC read-through have been successfully applied to treat nonsense variation-associated diseases. In this review, we summarize the NMD mechanism and discuss the application and progress in our understanding of NMD escape in disease therapy. This review should provide a useful framework to advance current understanding of the research and application of NMD escape.
Keywords:
PDF (559KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
程苗苗, 曹延延. NMD逃逸机制及其在疾病治疗中的应用. 遗传[J], 2020, 42(4): 354-362 doi:10.16288/j.yczz.19-335
Miaomiao Cheng.
无义介导的mRNA降解(nonsense-mediated mRNA decay, NMD)是一种广泛存在于真核生物的mRNA质量监控机制,特异性识别并降解含有提前终止密码子(premature termination codon, PTC)的mRNA[1],同时NMD也是机体调控基因表达的重要方式之一。在正常生理情况下,mRNA的可变剪接、转录错误或程序性基因重排可产生PTC,触发NMD途径使其降解,从而阻止有害的截短蛋白的产生。在病理情况下,以无义和移码变异为主的基因突变也可产生PTC,通过触发NMD降解mRNA导致靶蛋白表达下降从而引发疾病[2,3]。约1/3已知病因的人类遗传病是由基因突变产生的PTC所致[4]。
在哺乳动物细胞中,前体mRNA被剪接后在距离外显子与外显子连接点(exon-exon junction, EEJ)上游约20~24个核苷酸的位置处会形成一个外显子-外显子连接点复合物(exon-exon junction complex, EJC)。在正常的翻译过程中,核糖体沿着mRNA移动,替换下游所有的EJC并在正常的终止密码子处停止翻译,产生全长蛋白。若mRNA中存在PTC,翻译核糖体会在PTC处停止,募集UPF1、SMG1和两个翻译释放因子(eRF1和eRF3)形成SURF复合物。该复合物通过对下游EJC的识别,使NMD的核心组分-UPF1、UPF2和UPF3相互作用,最终导致UPF1发生磷酸化。磷酸化的UPF1一方面靶向作用于翻译起始因子eIF3抑制下一轮翻译的开始,另一方面进一步招募具有核酸内切酶和核酸外切酶活性的SMG系列因子介导mRNA的降解[5]。NMD效率在不同的转录本、细胞和组织类型以及个体间均存在广泛的变异性,更有甚者甚至不发生NDM,即产生了所谓的NMD逃逸(NMD escape)现象。本文对NMD逃逸的机制及其在临床中的应用进行了综述。
1 NMD逃逸与机制
通常,当PTC距离最后一个EEJ大于或等于50~55个核苷酸即可触发NMD,称为NMD途径的50边界规则[6]。然而,研究者发现某些PTC虽然符合这个规则,但是却没有发生NMD导致mRNA的降解,****称这种现象为NMD逃逸。通过分析功能基因组学数据库发现有52.7%的人群常见的无义变异发生了NMD逃逸[7]。研究表明这些发生NMD逃逸的转录本,其PTC多位于距离翻译起始位点较近的区域,比如一些位于人β球蛋白基因外显子1的PTC并未触发NMD而导致mRNA的降解,携带这类PTC的转录本与野生型β球蛋白mRNA转录水平相当[8]。这种现象也被称为“AUG邻近效应”[9,10]:即在加帽介导的翻译起始过程中,胞质多聚腺苷酸结合蛋白1 (PABPC1)通过与真核起始因子eIF4G和eIF3之间的相互作用,靠近40S核糖体亚基,形成环状结构。PABPC1也因此邻近AUG起始密码子。随着翻译延伸的开始,这个闭合的环状结构使PABPC1与位于AUG附近PTC上的终止复合物近在咫尺。此时PABPC1竞争性的与终止复合物中的eRF3相互作用,从而削弱了eRF3与NMD关键因子UPF1间的作用,抑制NMD途径。虽然NMD逃逸的现象很早就有报道,但是近些年才开展系统性研究且确切机制尚不十分清楚,目前比较公认的学说有两个:PTC通读和翻译的重新启动。蛋白质的翻译过程大体可以分为翻译的起始、肽链的延伸、翻译的终止以及翻译终止后核糖体复合物的解体即核糖体的循环再利用四个阶段。由于NMD与蛋白质的翻译偶联,因此NMD逃逸也与蛋白质的翻译过程息息相关。
1.1 PTC通读
PTC通读是指在蛋白翻译过程中,由于tRNA错配或核糖体移码或跳跃等原因导致翻译可以顺利通过PTC,直至遇到正常的终止密码子。就通读效率而言,PTC的通读效率约为正常终止密码子的10倍,因而哺乳动物细胞中通读更倾向于发生在PTC,而对于正常终止密码子的通读则极为罕见且低效[11]。在生理情况下,真核生物中蛋白质翻译的起始需要核糖体大小亚基、起始tRNA (Met-tRNAMet)和包括起始因子(eIF1、eIF2和eIF3)在内的几十个蛋白因子的参与,在模板mRNA编码区的5′端形成核糖体-mRNA-起始tRNA复合物,并将甲硫氨酸放入核糖体的P位。
蛋白翻译起始后,随着下一个氨基酸由氨酰- tRNA (AA-tRNA)运送到核糖体的A位,在肽基转移酶的催化下,与位于P位的上一个氨基酸生成肽键,肽链得以不断延长。同时核糖体也向mRNA模板的3′端方向逐步移动一个密码子,mRNA模板上的密码子决定了何种AA-tRNA能被结合到A位上。当终止密码子(UAG、UGA或UAA)出现在核糖体的A位时,没有相应的AA-tRNA能与之结合,取而代之的是释放因子。释放因子能识别终止密码子并与之结合,水解P位上多肽链与tRNA之间的二酯键。因其具有GTP酶活性,通过催化GTP水解,使肽链与核糖体解离。在真核细胞中,释放因子eRF1识别终止密码子,催化新合成的多肽链从P位点的tRNA中水解释放出来;而具有GTP酶活性的释放因子eRF3在多肽链释放后刺激eRF1从核糖体A位解离,至此蛋白质的合成结束。
因此,蛋白质的翻译是终止还是继续取决于释放因子和非同工tRNA之间相互竞争与核糖体A位点的结合[3]。当释放因子识别PTC并与核糖体大亚基的A位结合时,则翻译终止,触发NMD途径,mRNA发生降解。虽然细胞内存在多种校正机制以确保mRNA上的密码子可以被正确识别,但是准确率仍然达不到100%。当携带某种氨基酸的非同工tRNA与核糖体A位偶然结合,使无义密码子重新被解码为有义密码子,PTC被该氨基酸所取代,翻译继续进行,即表现为由于tRNA错配导致的PTC通读(图1A)。此外,通常情况下核糖体对mRNA密码子的识别是由起始密码子开始,沿5′至3′端方向,一个接一个直到终止密码子,但也有一些例外,翻译过程中某些反式作用因子决定核糖体在mRNA特定位点上的读码框可以发生程序性移动一个或者多个核苷酸后继续翻译,这一过程称为程序性核糖体移码或核糖体跳跃(ribosome frameshifting/jumping) (图1B)[12]。当核糖体恰巧跳过了PTC,则引发由于核糖体跳跃导致的PTC通读,但是目前尚缺少相关报道。
图1
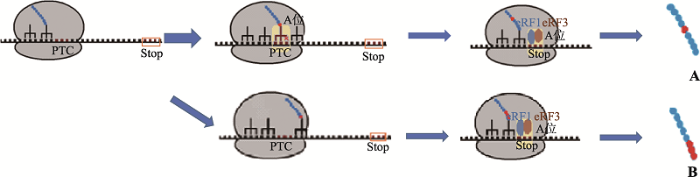 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1PTC通读机制
A:tRNA错配。由于tRNA错配,使PTC被解码为有义密码子,生成氨基酸(红色),翻译继续向下进行,发生PTC通读。B:核糖体跳跃。核糖体在mRNA特定位点上的读码框发生程序性移动,即程序性核糖体移码或核糖体跳跃,当核糖体恰巧跳过了PTC,则引发由于核糖体跳跃导致的PTC通读。PTC:提前终止密码子;Stop:终止密码子;eRF1:翻译释放因子1;eRF3:翻译释放因子3;蓝色为野生型氨基酸序列;红色为非野生型氨基酸序列。
Fig. 1Mechanism of translational read-through
综上所述,因PTC通读引发的NMD逃逸,具有以下两个特点:首先,由于未触发NMD途径,因此携带PTC转录本的转录水平与野生型相当;其次,由于PTC被氨基酸取代,且翻译终止于正常的终止密码子,因而可产生全长的蛋白质。虽然取代PTC的氨基酸可能与正常蛋白相应位置的氨基酸不同,但是如果该位置不是蛋白的关键功能区域,那么新生成的蛋白可发挥正常或部分正常功能。因此,这一机制也是目前主要用于抑制病理性NMD的有害作用、上调功能蛋白的表达从而靶向治疗遗传性疾病的分子基础[13],使用氨基糖甙类抗生素、非氨基糖甙类抗生素、小分子化合物(如PTC124)以及可与PTC结合的tRNA衍生物等均可诱导PTC的通读[14]。
Menkes综合征是一种由ATP7A基因突变导致铜转运障碍而引起的多系统受累的X连锁隐性遗传病,患者主要表现为神经系统变性和结缔组织异常。ATP7A基因位于Xq13.3,包含23个外显子,编码1500个氨基酸的蛋白产物ATP7A。Kaler等[15]报道了1例Menkes综合征患者携带有c.746C>T (p.Arg201Ter)无义变异,该变异在ATP7A基因的第3外显子产生一个PTC。患者cDNA的Sanger测序在相应位置可见明显C>T的碱基替换峰,提示携带PTC的mRNA转录本未发生显著的降解;应用Western blot和免疫组化分析患儿细胞,均可检测到少量全长ATP7A蛋白。以上的检测结果符合由于通读机制导致的NMD逃逸特点。因此,作者推断携带c.746C>T (p.Arg201Ter)无义变异的转录本发生了通读导致的NMD逃逸,为了进一步明确该通读是否源于tRNA错配,作者试图检测氨基酸序列,遗憾的是此例患者NMD抑制效率不是很高,未能足量分离和纯化通读产物。但是,酵母互补实验揭示该突变体表达的蛋白仍保留有部分铜转运功能,提示PTC的通读产物保留了部分功能。加之该患者在出生后不久即被诊断并进行铜注射治疗,因此治疗效果显著,无明显的神经系统症状出现[15]。
1.2 翻译的重新启动
蛋白质翻译终止后,留下一个由mRNA、位于P位的去酰化-tRNA和核糖体组成的翻译终止后核糖体复合物。随后,在ATP酶ABCE1、起始因子eIF3、eIF1、eIF1A以及eIF3j的协同作用下,翻译终止后核糖体复合物解体为mRNA、去酰化-tRNA和核糖体单体或其亚基[3]。解体后的核糖体进入新一轮的蛋白质合成。翻译的重新启动是指在PTC的下游存在着AUG,即潜在的翻译起始点。由于Met的三联密码同为AUG与起始密码子相同,因此可作为PTC下游翻译重新启动的潜在起始点。翻译终止后,核糖体复合物并未完全解离进入再循环,而是沿着mRNA继续滑动,当它遇到潜在的翻译起始点时,蛋白质的翻译重新开始[16]。
已有报道邻近翻译起始密码子的PTC,由于在其下游重新启动了有效的翻译导致携带该PTC的mRNA转录本发生了NMD逃逸,包括神经鞘瘤相关基因SMARCB1[17]、乳腺癌相关基因BRCA1、LQT2基因[18]、α和β珠蛋白基因[10]等,提示有效的翻译重新启动可能存在着边界效应,位于边界内的PTC可通过翻译重新启动而逃离NMD,而边界外的PTC则不能;此外,不同基因转录本的重新启动,其边界也有所不同,即便是密切相关的α和β珠蛋白基因也是如此。
Jagannathan等[7]利用生物信息学方法分析了人群中单核苷酸变异下游50个密码子的序列,研究发现与同义变异相比,Met在无义变异下游的50个密码子内存在显著的富集:57%的罕见和85%的常见无义变异在其下游50个密码子内存在Met,即潜在的翻译起始点;而只有52%的同义变异存在该现象,且这一富集现象仅在编码序列的前10%中成立。以上研究提示,有效的翻译重新启动与正常的起始密码子、PTC和潜在起始密码子3者之间的距离有关。首先,翻译终止后的核糖体与正常的起始密码子足够接近,也就是PTC需要靠近正常的翻译起始密码子;其次PTC距离下游的潜在起始点也较近,使得核糖体可以相对较快的遇到起始位点重启翻译。
尽管翻译的重新启动一直被认为是NMD逃逸的重要原因,但其发生的具体机制尚不十分清楚。具有短uORF转录本的翻译后重新启动是机体应对环境变化调节基因表达的途径之一。研究发现这类转录本在翻译终止后,只有核糖体的大亚基和去酰化tRNA被循环再利用,小亚基仍保留在mRNA上,进入下游从而发生翻译的重新启动。进一步研究显示真核生物起始因子eIF3,在翻译的延伸和终止过程中可与核糖体发生短暂结合[19,20],eIF3可特异性稳定小亚基,使其不被循环,保留eIF3的核糖体能够继续向下游移动,识别PTC下游的潜在起始密码子重新启动翻译(图2)。综上所述,因翻译的重新启动引发的NMD逃逸,同样由于未触发NMD途径,携带PTC转录本的转录水平也与野生型相当;其次,翻译重新启动发生于PTC下游的潜在起始点,因而可产生N端截短蛋白。
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2翻译的重新启动机制
PTC:提前终止密码子;Stop:终止密码子;eRF1:翻译释放因子1;eRF3:翻译释放因子3;AUG:起始密码子;eIF3:真核起始因子3。
Fig. 2The reinitiation mechanism
BRCA1基因与乳腺癌的发生高度相关,该基因发生突变使女性容易罹患乳腺癌和卵巢癌。该基因定位于17q21,含有23个外显子,在DNA修复、细胞周期调控、蛋白质泛素化和染色质重塑中发挥重要作用[21]。Buisson等[22]研究发现BRCA1基因的移码变异185delAG (c.68_69delAG)和188del11 (c.71_81del),分别在第36位(Ter36)和39(Ter39)位氨基酸位置产生PTC,且均位于第3外显子理论上应该触发NMD,但是Northern blot检测显示携带突变的转录本与野生型表达量相当;Western blot检测发现Ter36与Ter39突变质粒可以产生N端截短蛋白。以上检测结果符合由翻译的重新启动所致的NMD逃逸。进一步分析BRCA1基因序列显示在PTC下游22(Met48)和262(Met128)核苷酸的位置分别存在有Met,即潜在的翻译起始位点。为了明确翻译重新启动的具体位置,研究者利用定点诱变技术分别将Ter36与Ter39突变体的Met48和Met128突变为缬氨酸转染细胞,Western blot发现当Met48突变为缬氨酸时,Ter36+Met48Val与Ter39+Met48Val突变体产生的蛋白条带与均与Met48突变之前一致;而当Met128突变为缬氨酸时,Ter36+Met128Val与Ter39+Met128Val突变体则不产生N端截短蛋白。由此推测185delAG(c.68_69delAG)和188del11 (c.71_81del)变异发生NMD逃逸是原于在PTC下游的Met128位置发生了翻译的重新启动。
1.3 其他影响NMD逃逸的因素
除PTC通读和翻译的重新启动外,还存在其他一些因素可能导致PTC的NMD逃逸[23,24],如mRNA的剪接。当mRNA的选择性剪接将包含PTC的外显子从转录本中去除,则不再发生NMD。同样,如果PTC下游的某个单一内含子拼接效率低下,PTC下游没有EJC来发出NMD信号,从而无法触发NMD。其他影响NMD效率的因素还有3′非翻译区(UTR)的序列和结构。一些含有长3′ UTR的mRNA中存在着抑制NMD的顺式作用元件。研究发现位于TRAM1基因mRNA的3′UTR前200个核苷酸的位置存在一个顺式作用元件,当其位于终止密码子下游附近时可以抑制NMD的发生[25]。此外,由于NMD是一个翻译依赖的过程,影响翻译的顺式作用元件和反式作用因子均可以影响NMD的结局。2 NMD逃逸在疾病治疗方面的进展
随着对NMD及其逃逸的研究深入,近几年提出来许多针对PTC相关疾病的治疗策略,研究主要集中在对PTC通读机制的应用,即通过使用药物使致病的PTC发生通读,上调有功能的蛋白表达,从而抑制病理性NMD的有害作用。通读治疗已经被证明是治疗无义变异相关疾病的较成功方案,已在囊性纤维化、杜氏肌营养不良(Duchenne muscular dystrophy, DMD)、血友病、代谢紊乱、眼遗传性疾病和癌症等疾病中取得良好的疗效。PTC通读的治疗首个突破发生在20世纪晚期,研究者发现一些化合物能够在序列中引入一种天然氨基酸取代PTC,从而发生通读产生完整的多肽链,避免因触发NMD导致的mRNA降解[26]。随着研究的深入,越来越多的药物被确定使用(表1)。目前比较成功的治疗方法主要有两类,即药物和分子生物学干预,用于治疗PTC引起的疾病。
Table 1
表1
表1基于促通读机制的药物研究应用现况
Table 1
| 通读药物 | 实验模型 | 临床试验 | 相关疾病 | 特点与问题 |
|---|---|---|---|---|
| 氨基糖苷类 | ||||
| 庆大霉素[27] | 动物和患者 | 有 | 囊性纤维化(CF)、杜氏肌营养不良(DMD)、瑞特综合征(RTT)、共济失调性毛细血管扩张症(AT)、法布里病(Fabry)、尼曼匹克症A/B型(NPA/NPB)、全身性神经节苷脂贮积(Gangliosidosis I )、粘多糖累积症II型(MPS II) 、MPS I-H,MPS IIIB,MPS VI、血友病A和B(HA和HB)、无脉络膜症(Choroideremia)、先天性黑朦(LCA type 2)、眼缺损(Ocular coloboma )和Usher综合征I型(USH1) | 副作用如耳毒性、肾毒性和视网膜毒性 |
| 阿米卡星[28] | 动物和细胞 | 无 | 囊性纤维化(CF)、瑞特综合征(RTT)、粘多糖 累积症I-H(MPS I-H)和脊髓型肌萎缩症I型 (SMA I) | |
| 妥布霉素[29] | 动物 | 无 | 囊性纤维化和脊髓型肌萎缩症I型(SMA I) | |
| 巴龙霉素[30] | 细胞 | 无 | 囊性纤维化(CF)、杜氏肌营养不良(DMD)、 瑞特综合征(RTT)、无脉络膜症(Choroideremia)、粘多糖累积症I-H(MPS I-H)、眼缺损(Ocular coloboma )和Usher综合征I型(USH1) | |
| G418[28] | 动物和细胞 | 无 | 囊性纤维化(CF)、瑞特综合征(RTT)、共济失调性毛细血管扩张症(AT)、粘多糖累积症I-H(MPS I-H)、脊髓型肌萎缩症I型(SMA I)、先天性黑朦(LCA type 2)、色素性视网膜炎(RP)和Usher综合征I型(USH1) | |
| 氨基糖苷类衍生物 | ||||
| 新霉素衍生物 (TC003和TC007)[29] | 动物和细胞 | 无 | 脊髓型肌萎缩症I型(SMA I) | 生物相容性更好 |
| 卡那霉素衍生物 (JL023)[29] | 脊髓型肌萎缩症I型(SMA I) | 无明显副作用 | ||
| 巴龙霉素衍生物 (NB30、NB54、 NB84和NB124)[31] | 动物和细胞 | 无 | 囊性纤维化(CF)、瑞特综合征(RTT) Usher综合征I型(USH1)和粘多糖累积症I-H(MPS I-H) | 较高的生物相容性、剂量依赖性,可通过血脑屏障,无明显副作用 |
| 非氨基糖苷类[32] | ||||
| PTC124 | 动物和患者 | 有 | 囊性纤维化(CF)、杜氏肌营养不良(DMD)、粘多糖累积症VI型(MPS VI)、神经元蜡样脂褐质沉积症(NCLs)、贝敦氏病(Batten disease)、色素性视网膜炎(RP)、Usher综合征I型(USH1)和先天性无虹膜(Congenital aniridia) | 耐受性良好,无严重副作用 |
| RTC13、RTC14和 RTC229 | 细胞 | 无 | 杜氏肌营养不良(DMD)和共济失调性毛细血管扩张症(AT) | 对3种提前终止密码子均有效 |
新窗口打开|下载CSV
2.1 氨基糖苷类
氨基糖苷类是应用较广的一类抗生素,可通过干扰核糖体校正作用减少核糖体翻译过程的提前终止。氨基糖苷类药物选择性地与核糖体的解码中心结合,导致氨基酸的随机引入,从而克服肽链中的PTC,发生通读[33]。例如,庆大霉素是一种常见的氨基糖苷类抗生素,通过与真核核糖体结合发挥无义抑制活性[34],选择性地在PTC中引入氨基酸,但不影响正常终止密码子。在作用过程中,庆大霉素通过促进近同工tRNA与核糖体A位点的结合而抑制校对过程,使翻译的保真度降低,从而导致误读和/或PTC抑制[35]。Hurler综合征被认为是最严重的遗传性溶酶体贮积病,为常染色体隐性遗传病。由于IDUA基因变异导致所编码的α-L-艾杜糖醛酸苷酶缺乏,溶酶体内的粘多糖(GAG)不能正常降解,引发粘多糖贮积病。IDUA基因位于1号染色体,p.Gln70Ter和p.Trp402Ter是欧洲裔Hurler综合征患者中较常见的两种无义变异[36]。2001年,Keeling等[37]使用Hurler综合征成纤维细胞系研究了庆大霉素对IDUA基因p.Gln70Ter和p.Trp402Ter变异的抑制活性,发现经庆大霉素处理后可以显著提升α-L-艾杜糖醛酸苷酶的活性。但是由于长期应用庆大霉素产生副作用,研究者也尝试设计更为有效且毒性更小的氨基糖苷类药物,其中一种氨基糖苷类衍生物-NB84,在疗效上可恢复α-L-艾杜糖醛酸苷酶活性,且几乎没有毒性[31]。
2.2 Ataluren
Ataluren (原称PTC124)是一种非氨基糖苷类的小分子化合物,能够在mRNA转录本的PTC位置随机插入氨基酸,诱导PTC通读,而不影响正常的终止密码子[38]。DMD由编码抗肌萎缩蛋白的基因突变引起,患者主要表现为进行性近端肌肉无力,导致行走困难、依赖轮椅以及最终的呼吸和心脏衰竭恶化。由无义变异产生PTC导致截短蛋白或蛋白质功能丧失约占DMD病例的13%。Ataluren是通过药物的高通量筛选和化学优化而被发现的,它可以诱导核糖体读取PTC,而不是正常的终止密码子。当在无义突变的DMD小鼠模型中测试时,Ataluren可产生全长的功能性抗肌萎缩蛋白。在随后的人DMD实验也成功诱导出了全长功能性抗肌萎缩蛋白[39]。基于PTC124药物的有效性,2017年PTC124通过了欧盟的年度审核。目前,PTC公司正在进行一个随机双盲、安慰剂对照、国际多中心的III期临床试验,以进一步证实PTC124长期治疗无义变异的DMD患者的效果和安全性。该临床试验总目标入组人数250人,中国入组人数50人,预期2021年将公布结果(2.3 抑制子tRNA
无义抑制子是一种tRNA的衍生物,其反密码子能够识别PTC,继而在PTC处掺入氨基酸,产生全长功能蛋白,防止翻译发生终止。抑制子介导的密码子-反密码子的相互作用,可以避免由于核糖体校正而发生的氨酰-tRNA排斥,因此可以有效抑制PTC。研究表明,抑制子tRNA可用于人体由无义变异引起的疾病的体细胞基因治疗。遗传性弥漫性胃癌(HDGC)是一种侵袭性、无法治愈的胃癌,主要由CDH1基因的无义变异引起,导致截短的E-钙粘蛋白产生[40]。最近一项关于HDGC的研究显示,在给予抑制子tRNA诱导后,108个具有HDGC家族史的家庭中,有23个家庭(21.3%)的癌症的发病显著延迟,并使由携带PTC的CDH1基因可以编码产生大约30%功能性E-钙粘蛋白[41]。3 结语与展望
近年来,关于NMD逃逸的研究成果很多,但机制仍未得到完全阐明。随着研究的深入,PTC相关疾病的致病机制和药物治疗越来越成为研究的热点,目前研究成果主要集中在促通读药物的研发,这些药物为遗传病的治疗带来了新希望。然而,促通读药物的临床应用仍存在很多亟待解决的问题,如庆大霉素等氨基糖苷类药物的长期应用有耳毒性、肾毒性等副作用,静脉注射仍是许多氨基糖苷类药物的用药途径,不适于患者长期应用。此外,由于NMD逃逸和新型药物的作用机制尚未完全阐明,使得药物在临床试验中疗效不稳定。针对这些问题,尚需开展更多NMD逃逸机制以及药物作用机制的研究,以提高药物疗效的特异性。综上所述,NMD逃逸使得携带PTC的转录本仍可编码具有部分功能的蛋白质,缓解患者表型。最近,马志鹏等[42]通过引入PTC利用NMD途径构建敲除capn3a基因的斑马鱼突变体,但是却发现该突变体发育正常并未出现预期的异常表型;同时他们还发现该突变体中多个capn家族基因上调弥补capn3a基因的功能,推测发生这种现象是由于遗传补偿效应,并提出假设:无义突变mRNA通过提高同源基因的表达来实现补偿效应。经实验证实无义突变与核酸序列同源性是激活遗传补偿效应的两个必要条件。该研究阐释了PTC不导致表型的另一重要机制,也为PTC相关疾病的致病机制和药物治疗研究提供了新方向。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 3]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}