Carbon density and its spatial distribution in the Potentilla fruticosa dominated alpine shrub in Qinghai, China
YANGLu-Cun1,2, LIChang-Bin1,3, NINGYi1,3, NIEXiu-Qing1,3, XUWen-Hua1,2, ZHOUGuo-Ying1,2,*, 1Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining 810008, China2Key Laboratory of Tibetan Medicine Research, Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining 810008, Chinaand 3University of Chinese Academy of Sciences, Beijing 100049, China 通讯作者:* 通信作者Author for correspondence (E-mail: zhougy@nwipb.ac.cn) 责任编辑:YANGLu-CunLIChang-BinNINGYiNIEXiu-QingXUWen-HuaZHOUGuo-Ying 收稿日期:2016-01-11 接受日期:2016-11-11 网络出版日期:2017-01-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:中国科学院战略性先导科技专项(XDA- 0505030304)、科技基础性工作专项(2015FY1103- 0001)和国家科技支撑计划项目(2014BAC05B01)
关键词:生物碳密度;凋落物碳密度;土壤有机碳密度;总碳密度 Abstract Aims Shrub recovery is recognized as an important cause of the increase of carbon stocks in China, and yet there are great uncertainties in the carbon sink capacities of shrubs. Our objectives were to estimate carbon density and its spatial distribution in alpine shrubs. Methods Eight sites in Potentilla fruticosa dominated shrublands across Qinghai, China were investigated. Plant biomass and carbon content in leaves, branches and stems, and roots were measured to analyze the biomass allocation and carbon density. Important findings Mean carbon densities in biological carbon, litter, soil and whole ecosystem of P. fruticosa shrublands were 5088.54, 542.1, 35903.76 and 41534.4 kg·hm-2, respectively. Carbon density in the shrub layer was more than 68% of the biological carbon density of the whole ecosystem and was mainly distributed in roots (49.5%-56.1%). Carbon density of the herbaceous layer was 22.5% of the biological carbon density of the whole ecosystem and was also mainly distributed in roots (59.6%-75.1%). The biological carbon density of P. fruticosa shrublands (5.08 t·hm-2) was lower than the average carbon density of shrub communities in China (10. 88 t·hm-2). Soil carbon density contributed the largest proportion (85.8%) of total carbon density in P. fruticosa shrublands.
由表2可得, 青海省不同样地金露梅灌丛灌木层的生物量介于5952.05-12023.80 kg·hm-2之间, 平均为8361.92 kg·hm-2。金露梅灌丛灌木层各器官生物量大小排序为: 根>枝>叶。青海省不同样地金露梅灌丛草本层的生物量差异较大, 介于1075.41-11386.98 kg·hm-2之间, 平均为4293.37 kg·hm-2。草本层地下部分生物量明显大于地上部分。青海省不同样地金露梅灌丛凋落物层的生物量介于365.77- 2846.63 kg·hm-2之间, 平均为1246.66 kg·hm-2。 Table 2 表2 表2青海不同样地金露梅灌丛各组分生物量(平均值±标准误差) (kg·hm-2) Table 2Biomass of the Potentilla fruticosa shrub ecosystem in different sampling sites in Qinghai (mean ± SE) (kg·hm-2)
层次/器官 Layer/organ
样地 Sampling site
平均 Average
青海门源 Menyuan, Qinghai
青海祁连 Qilian, Qinghai
青海玉树 Yushu, Qinghai
青海刚察Gangcha, Qinghai
青海天峻Tianjun, Qinghai
青海大通Datong, Qinghai
青海兴海Xinghai, Qinghai
青海贵南Guinan, Qinghai
灌木层 Shrub layer
叶 Leaf
391.81 ± 42.45a
665.50 ± 41.65b
598.030 ± 23.65ab
753.39 ± 90.58b
425.65 ± 68.75a
1 065.54 ± 91.63c
580.96 ± 45.58ab
1 057.52 ± 98.76c
692.30 ± 53.89
枝 Branch
2 677.35 ± 478.95abc
3 083.58 ± 468.43abc
4 527.30 ± 1014.2c
3 548.54 ± 135.69abc
2 313.81 ± 445.49ab
2 055.72 ± 446.22a
2 206.73 ± 119.62a
4 198.33 ± 864.11bc
3 076.43 ± 248.61
根 Root
3 405.82 ± 822.31a
4 745.91 ± 831.42abc
5 538.02 ± 235.86bc
5 715.92 ± 675.68bc
3 212.59 ± 852.82a
4 119.84 ± 605.00ab
3 239.50 ± 178.57a
6 767.94 ± 599.21c
4 593.19 ± 321.39
合计 Total
6 474.98 ± 1231.8a
8 494.99 ± 1 205.1ab
10 663.36 ± 897.24bc
10 017.85 ± 900.44bc
5 952.05 ± 918.99a
7 241.10 ± 1060.2ab
6 027.10 ± 302.16a
12 023.80 ± 1 536.0c
8 361.92 ± 549.46
草本层 Herb layer
叶 Leaf
406.21 ± 64.50a
762.57 ± 115.41ab
1 534.51 ± 292.50b
834.52 ± 48.88ab
1 402.72 ± 684.49b
340.36 ± 119.32a
683.52 ± 128.13ab
727.74 ± 58.50ab
836.52 ± 116.46
根 Root
1 438.43 ± 388.53a
1 761.69 ± 45.60ab
9 852.47 ± 2 780.3c
5 486.23 ± 1 915.2b
5 494.34 ± 1 125.2b
735.05 ± 264.70a
1 274.49 ± 155.68a
1 612.09 ± 253.11ab
3 274.15 ± 753.52
合计 Total
1 844.64 ± 441.05a
2 524.26 ± 160.98ab
11 386.98 ± 2 973.4d
6 320.75 ± 1 964.0bc
6 897.06 ± 1 710.5c
1 075.41 ± 380.29a
1 958.01 ± 277.09ab
2 339.83 ± 285.17ab
4 293.37 ± 838.27
凋落物层 Litter layer
1 352.4 ± 149.86bc
1 653.93 ± 42.58c
2 846.63 ± 504.94d
826.93 ± 19.22ab
1 122.5 ± 143.50bc
897.57 ± 116.48ab
907.57 ± 97.57ab
365.77 ± 65.34a
1 246.66 ± 158.05
生物层 Biological layer
9 672.02 ± 1 635.3ab
12 673.18 ± 1 319.4ab
24 896.97 ± 4 321.5c
17 165.53 ± 2 217.8b
13 971.61 ± 2 643.3ab
9 214.08 ± 1184.5a
9 202.41 ± 647.34a
14 729.4 ± 1 364.9ab
13 940.65 ± 1 222.3
Different lowercase letters indicate significant differences between sampling sites (p < 0.05).不同小写字母表示不同样地间差异显著(p < 0.05)。 新窗口打开 青海省不同样地金露梅灌丛群落平均生物量大小排序为: 灌木层(60.26%) >草本层(30.80%) >凋落物层(8.94%), 且灌木层生物量所占比例明显高于草本层和凋落物层。
3.2 金露梅灌丛植被各组分、凋落物及土壤层有机碳含率
青海省不同样地金露梅灌丛平均含碳率分别为45.12%、44.86%、44.44%、44.02%、44.16%、45.04%、44.53%、45.15%, 不同样地间灌木层平均含碳率差异不显著(p > 0.05)。灌木各器官平均含碳率存在差异, 表现为枝(45.26%) >叶(45.15%) >根(43.69%)草本层、凋落物层含碳率分别为34.6%和43.0%, 均低于灌木层含碳率。青海省不同样地草本各器官的含碳率在31.0 %-39.5%之间。 土壤(0-100 cm)含碳率介于0.79%-12.29%之间(表3)。青海省不同样地表层0-10 cm土壤含碳率均高于其他深层土壤, 且随土层深度增加含碳率逐渐减小, 具有明显的垂直分布特征。 Table 3 表3 表3青海各样地金露梅灌丛土壤含碳率(%) Table 3Soil carbon contents of the Potentilla fruticosa shrublands in different sampling sites in Qinghai (%)
样地 Sampling site
土层 Soil layer (cm)
合计 Total
0-10
10-20
20-30
30-50
50-70
70-100
Q1
9.98
8.02
8.54
3.36
1.60
0.79
32.29
Q2
9.86
9.19
6.63
3.05
3.71
1.03
33.47
Q3
9.78
7.00
5.71
4.38
3.83
3.69
34.39
Q4
9.03
6.81
5.80
3.49
3.19
2.52
30.84
Q5
9.42
6.18
5.66
1.71
1.73
2.44
27.14
Q6
3.55
5.06
6.68
5.53
4.07
-
24.89
Q7
12.29
5.50
4.13
4.43
4.72
4.59
35.66
Q8
8.31
6.70
3.27
2.83
2.70
2.58
26.39
平均 Average
9.03
6.81
5.80
3.60
3.19
2.52
30.63
-, 70-100 cm of soil was not obtained for the reason of gravel. Sampling site see Table 1.-, 在取土样时70-100 cm的土壤由于砾石的原因未取到。样地代码见表1。 新窗口打开
3.3 金露梅灌丛植被、凋落物及土壤层碳密度及分配格局
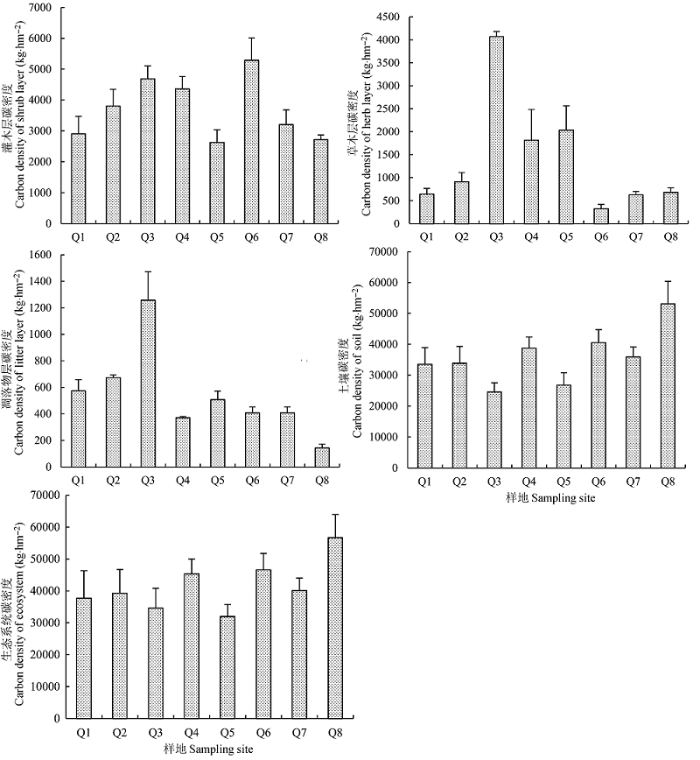
由表4和图1可得, 青海省不同样地金露梅灌丛灌木层碳密度存在显著差异(图1), 碳密度大小依次为: Q6 > Q3 > Q4 > Q2 > Q7 > Q1 > Q8 > Q5, 平均为3698.6 kg·hm-2。青海省不同样地金露梅灌丛灌木层碳主要分配在根部(49.5%-56.1%)。灌木地上部分(叶+枝)碳密度占灌木层碳密度的比例依次为: Q3 > Q1 > Q5 > Q8 > Q6 > Q2 > Q4 > Q7。灌木层碳密度占各生态系统生物层碳密度的68%以上, 占 各生态系统碳密度的比例分别为Q3 (2.54%) > Q6 (1.28%) > Q2 (1.1%) > Q5 (0.96%) > Q1 (0.86%) > Q8 (0.51%)。 Table 4 表4 表4青海各样地金露梅灌丛生态系统各组分碳密度及分配格局(平局值±标准误差) Table 4Density and distribution pattern of catbon in the Potentilla fruticosa shrub ecosystem in different sampling sites in Qinghai(mean±SE)
新窗口打开 显示原图|下载原图ZIP|生成PPT 图1青海各样地金露梅灌丛碳密度(平均值±标准误差)。 -->Fig. 1Carbon density of the Potentilla fruticosa shrub ecosystem in different sampling sites in Qinghai (mean ± SE). -->
DongX (2009). Evaluation of forest resources in Qinghai Province .Journal of Anhui Agricultural Sciences, 37, 5727-5728, 5751. (in Chinese with English abstract)[董旭 (2009). 青海省森林资源评价 . 安徽农业科学, 37, 5727-5728, 5751.] [本文引用: 2]
[2]
FanQS, ShaZJ, CaoGC, CaoSK (2005). Assessment of ecology and environments on climate changing of Qinghai- Tibetan Plateau .Journal of Salt Lake Research, 13(1), 12-18. (in Chinese with English abstract)[樊启顺, 沙占江, 曹广超, 曹生奎 (2005). 气候变化对青藏高原生态环境的影响评价 . 盐湖研究, 13(1), 12-18.] [本文引用: 1]
[3]
GaoQ, YangXC, YinCY, LiuQ (2014). Estimation of biomass allocation and carbon density in alpine dwarf shrubs in Garzê Zangzu Autonomous Prefecture of Sichuan Province, China .Chinese Journal of Plant Ecology, 38, 355-365. (in Chinese with English abstract)[高巧, 阳小成, 尹春英, 刘庆 (2014). 四川省甘孜藏族自治州高寒矮灌丛生物量分配及其碳密度的估算 . 植物生态学报, 38, 355-365.] [本文引用: 1]
[4]
HaoWF, ChenCG, LiangZS, MaL (2008). Research advances in vegetation biomass . Journal of Northwest A & F University (Natural Science Edition), 36, 175-182. (in Chinese with English abstract)[郝文芳, 陈存根, 梁宗锁, 马丽 (2008). 植被生物量的研究进展 . 西北农林科技大学学报(自然科学版), 36, 175-182.] [本文引用: 1]
[5]
HouXY (1982). Vegetation Map of the People’s Republic of China. China Map Press, Beijing. (in Chinese)[侯学煜 (1982). 中华人民共和国植被图. 中国地图出版社, 北京. ] [本文引用: 1]
[6]
HuHF, WangZH, LiuGH, FuBJ (2006). Vegetation carbon storage of major shrublands in China .Journal of Plant Ecology (Chinese Version), 30, 539-544. (in Chinese with English abstract)[胡会峰, 王志恒, 刘国华, 傅伯杰 (2006). 中国主要灌丛植被碳储量 . 植物生态学报, 30, 539-544.] [本文引用: 2]
[7]
JinM, LiY, WangSL, ZhangXL, LeiL (2012). Alpine shrubs biomass and its distribution characteristics in Qilian Mountains .Arid Land Geography, 35, 952-959. (in Chinese with English abstract)[金铭, 李毅, 王顺利, 张学龙, 雷蕾 (2012). 祁连山高山灌丛生物量及其分配特征 . 干旱区地理, 35, 952-959.] [本文引用: 1]
[8]
JobbagyEG, JacksonRB (2000). The vertical distribution of soil organic carbon and its relation to climate and vegetation .Ecological Applications, 10, 423-426. [本文引用: 1]
[9]
KangWX, TianZ, HeJN, CuiSS (2009). Carbon cycle in ten kinds of forest ecosystem in Guangzhou City .Chinese Journal of Applied Ecology, 20, 2917-2924. (in Chinese with English abstract)[康文星, 田徵, 何介南, 崔莎莎 (2009). 广州市十种森林生态系统的碳循环 . 应用生态学报, 20, 2917-2924.] [本文引用: 1]
[10]
LeiL, LiuXD, WangSL, LiY, ZhangXL (2011). Assignment rule of alpine shrubs biomass and its relationships to environmental factors in Qilian Mountains .Ecology and Environmental Sciences, 20, 1602-1607. (in Chinese with English abstract)[雷蕾, 刘贤德, 王顺利, 李毅, 张学龙 (2011). 祁连山高山灌丛生物量分配规律及其与环境因子的关系 . 生态环境学报, 20, 1602-1607.] [本文引用: 1]
[11]
LiJ (2008). Study on Soil Carbon Stock and Its Spatial Distribution, Influence Factors in the Forest of China . Master degree dissertation, Sichuan Agricultural University, Ya’an, Sichuan. 18-21. (in Chinese with English abstract)[李江 (2008). 中国主工森林群落林下土壤有机碳储量格局及其影响因子研究 . 硕士学位论文, 四川农业大学, 四川雅安. 18-21.] [本文引用: 1]
[12]
LiYN, YuXF, XuZQ, LiuLL, YaoWX, WangL (2014). Carbon density characteristics of two typical shrub communities in the northern mountain region of Hebei .Scientia silvae sinicae, 50(6), 28-33. (in Chinese with English abstract)[李亚男, 虞晓凡, 许中旗, 刘乐乐, 姚卫星, 王丽 (2014). 冀北山地2种典型灌丛的碳密度特征 . 林业科学, 50(6), 28-33.] [本文引用: 2]
[13]
LiYN, ZhaoL, WangQX, DuMY, GuS, XuSX, ZhangFQ, ZhaoXQ (2006). Estimation of biomass and annual turnover quantities of Potentilla froticosa shrub . Acta Agrestia Sinica, 14(1), 72-76. (in Chinese with English abstract)[李英年, 赵亮, 王勤学, 杜明远, 古松, 徐世晓, 张发起, 赵新全 (2006). 高寒金露梅灌丛生物量及年周转量 . 草地学报, 14(1), 72-76.] [本文引用: 1]
[14]
LiuH, DongL, AbulaA, LiPJ, JiaRQ, WangWY (2015). Value of carbon sequestration of Tamarix in the midstream of the Tarim River . Acta Agriculturae Universitatis Jiangxiensis, 37, 484-489. (in Chinese with English abstract)[刘华, 董玲, 艾吉尔·阿不拉, 李丕军, 贾瑞琪, 王文月 (2015). 塔里木河中游柽柳灌丛碳储量及其价值评估 . 江西农业大学学报, 37, 484-489.] [本文引用: 1]
[15]
LuRK (1999). Analysis of Soil Agrochemistry. China Agriculture Scientech Press, Beijing. (in Chinese)[鲁如坤 (1999). 土壤农业化学分析方法. 中国农业科技出版社, 北京.] [本文引用: 1]
[16]
PanHL, LiMH, CaiXH, WuJ, DuZ, LiuXL (2009). Responses of growth and ecophysiology of plants to altitude .Ecology and Environmental Sciences, 18, 722-730. (in Chinese with English abstract)[潘红丽, 李迈和, 蔡小虎, 吴杰, 杜忠, 刘兴良 (2009). 海拔梯度上的植物生长与生理生态特性 . 生态环境学报, 18, 722-730.]
[17]
PiaoSL, FangJY, HuangY (2010). The carbon balance of terrestrial ecosystems in China .China Basic Science,12(2), 20-22. (in Chinese with English abstract)[朴世龙, 方精云, 黄耀 (2010). 中国陆地生态系统碳收支 . 中国基础科学, 12(2), 20-22.] [本文引用: 1]
[18]
QiaoCL, LiJM, WangJH, GeSD, ZhaoL, XuSX (2011). Annual carbon dioxide flux variations of alpine shrub ecosystem in the Qinghai-Tibet Plateau .Acta Agrestia Sinica, 19, 910-916. (in Chinese with English abstract)[乔春连, 李婧梅, 王基恒, 葛世栋, 赵亮, 徐世晓 (2011). 青藏高原高寒灌丛生态系统CO2通量年变化特征研究 . 草地学报, 19, 910-916.] [本文引用: 1]
[19]
RenLN, WangHY, DingGD, GaoGL, LiuL (2012). Effects of Larix principis-rupprechtii plantation density control on soil organic carbon and nutrients characteristics . Journal of Arid Land Resources and Environment, 26(4), 138-143. (in Chinese with English abstract)[任丽娜, 王海燕, 丁国栋, 高广磊, 刘玲 (2012). 密度调控对华北落叶松人工林土壤有机碳及养分特征的影响 . 干旱区资源与环境, 26(4), 138-143.]
[20]
SturmM, RacineC, TapeK (2001). Increasing shrub abundance in the Arctic .Nature, 411, 546-547. [本文引用: 1]
[21]
TianYY, QinF, YanH, GuoWH, GuanQW (2011). Carbon content rate in the common woody plants of China .Journal of Anhui Agriculture Science, 39, 16166-16169. (in Chinese with English abstract)[田勇燕, 秦飞, 言华, 郭伟红, 关庆伟 (2011). 我国常见木本植物的含碳率 . 安徽农业科学, 39, 16166-16169.] [本文引用: 1]
[22]
VeldkampE (1994). Organic carbon turnover in three tropical soils under pasture after deforestation .Soil Science Society of American Journal, 58, 175-180. [本文引用: 1]
[23]
WangGR, ChenXR, HanYZ, YangCD, XuCL (2006). Photosynthetic characteristics of three species ofLiriope . Grassland and Turf, (3), 27-34. (in Chinese with English abstract)[王国荣, 陈秀荣, 韩玉竹, 杨成德, 徐长林 (2006). 东祁连山高寒灌丛土壤微生物的分布特征 . 草原与草坪, (3), 27-34.] [本文引用: 1]
[24]
WangQJ, ZhouXM, ZhangYQ, ZhaoXQ (1991). Structure characteristics and biomass of Potentilla fruticose shrub in Qinghai-Tibetan Plateau . Acta Botanic Boreali-Occidentalia Sinica, 11, 333-340. (in Chinese with English abstract)[王启基, 周兴民, 张堰青, 赵兴全 (1991). 青藏高原金露梅灌丛的结构特征及其生物量 . 西北植物学报, 11, 333-340.] [本文引用: 2]
[25]
YangLC, ZhongZB, LiuHC, SongWZ, XuWH, ZhouGY, NieXM (2013). Community characteristics of alpine shrubSalix oritrepha . Bulletin of Botanical Research, 33, 653-658. (in Chinese with English abstract)[杨路存, 钟泽兵, 刘何春, 宋文珠, 徐文华, 周国英, 聂学敏 (2013). 高寒山生柳灌丛的群落特征研究 . 植物研究, 33, 653-658.] [本文引用: 1]
[26]
YangYH, FangJY, TangYH, JiCJ, ZhengCY, HeJS, ZhuB (2008). Storage, patterns and controls of soil organic carbon in the Tibetan grasslands .Global Change Biology, 14, 1592-1599. [本文引用: 1]
[27]
YangYJ, ChenYM, CaoY (2014). Carbon density and distribution of Pinus tabulaeformis plantation ecosystem in Hilly Loess Plateau . Acta Ecologica Sinica, 34, 2128-2136. (in Chinese with English abstract)[杨玉姣, 陈云明, 曹扬 (2014). 黄土丘陵区油松人工林生态系统碳密度及其分配 . 生态学报, 34, 2128-2136.]
[28]
YiLB (2012). Research and Assessment of Forest Carbon Potential in Qinghai .Master degree dissertation, Northwest A & F University, Yangling, Shaanxi. 5-52. (in Chinese with English abstract)[仪律北 (2012). 青海森林碳汇潜力分析与评估 . 硕士学位论文, 西北农林科技大学, 陕西杨凌. 51-52.]
[29]
YuHY, XuJC (2009). Effects of climate change on vegetations on Qinghai-Tibet Plateau: A review .Chinese Journal of Ecology, 28, 747-754. (in Chinese with English abstract)[于海英, 许建初 (2009). 气候变化对青藏高原植被影响研究综述 . 生态学杂志, 28, 747-754.] [本文引用: 1]
[30]
ZhangXH, ZhangYD, GuFX, LiuSR (2011). Dynamics of shrubs normalized difference vegetation index and its correlations with climatic factors in Southwest China .Chinese Journal of Ecology, 30, 2577-2583. (in Chinese with English abstract)[张笑鹤, 张远东, 顾峰雪, 刘世荣 (2011). 西南地区灌丛归一化植被指数动态及其与气候因子的相关性 . 生态学杂志, 30, 2577-2583.] [本文引用: 2]
[31]
ZhaoCC, LiuL, YangJJ, DongX (2013). Study on the biomass of Potentilla fruticosa forest in Qinghai loess hilly region . Guangdong Agricultural Sciences, (5), 179-182. (in Chinese with English abstract)[赵串串, 刘龙, 杨晶晶, 董旭 (2013). 青海黄土丘陵区金露梅灌丛生物量的研究 . 广东农业科学, (5), 179-182.] [本文引用: 1]
[32]
ZhouHK, ZhouL, ZhaoXQ, ShenZX, LiYN, ZhouXM, YanZL, LiuW (2002). Study of formation pattern of belowground biomass in Potentilla fruticosa shrub . Acta Prataculturae Sinica, 11(2), 59-65. (in Chinese with English abstract)[周华坤, 周立, 赵新全, 沈振西, 李英年, 周兴民, 严作良, 刘伟 (2002). 金露梅灌丛地下生物量形成规律的研究 . 草业学报, 11(2), 59-65.] [本文引用: 1]
青海省森林资源评价 2 2009
... 青海省位于青藏高原的东北部(31.65°-39.32° N, 89.58°-103.07° E) , 东西长约1200 km, 南北宽约800 km, 青海省土地总面积7215.24 × 104 hm2.其中林地面积634.00 × 104 hm2, 占8.79%; 森林面积329.56 × 104 hm2, 占林地面积的51.98%, 森林覆盖率4.57%.燕山运动奠定地形复杂多样, 高山、丘陵、河谷、盆地交错分布, 平均海拔3000 m以上, 属典型高原大陆性气候.年平均气温-3.7- 6.0 ℃, 年日照2340-3550 h, 年降水量16.7-776.1 mm (多为400 mm以下), 年蒸发量1118.4-3536.2 mm (董旭, 2009).森林植被分布于96° E以东主要江河及其支流的河谷两岸, 森林分布海拔大多在2500-4200 m, 以寒温带常绿针叶林亚型为主, 其次为落叶林植被型(多为原始林破坏后的次生类型).植被地域上跨青藏高原、温带荒漠和温带草原3个植被区, 具有高寒和旱生的特点, 以寒温性常绿针叶林为主, 常见的针叶树种有云杉(Picea spp.)、圆柏(Juniperus spp.)等, 阔叶树种有杨树(Populus spp.)、桦木(Betula spp.)等, 灌木有金露梅(Potentilla fruticosa)、杜鹃(Rhododendron spp.)、锦鸡儿(Caragana sinica)、杯腺柳(Salix cupularis)等(董旭, 2009). ...

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}