0 引言
【研究意义】水稻是我国三大主要粮食作物之一[1],其产量约占全国粮食总产量的34%,其播种面积约占全国粮食播种面积的27%[2]。氮素是水稻生长发育过程中土壤最易缺乏、需要量最大、损失潜力最大的元素[3]。近年来,为了保障水稻高产稳产,氮肥的投入快速增加,加上氮肥施用不合理等问题,不仅导致氮肥资源的浪费,而且导致氮肥利用率下降,氮素损失严重,大量的氮素通过氨挥发、硝化-反硝化、表面径流和淋溶等途径进入到大气和水环境,导致水体富营养化、地下水污染、土壤酸化、温室效应和大气污染等一系列环境问题[4,5,6,7]。如何根据作物需求进行有效的氮肥管理,充分发挥氮肥的增产潜力,最大限度减少氮素损失和对环境造成的污染,提高氮肥的利用率是我国水稻生产可持续发展面临的一个重大挑战[8,9]。【前人研究进展】尽管氮肥深施和氮肥分次施用等氮肥管理措施能够提高氮肥利用率,增加水稻产量,但因缺乏配套的施肥机械和有效的农村劳动力,往往限制了这些技术的推广应用[10,11,12];而且这些技术不能协调肥料养分释放特征与水稻养分需求特征相匹配[13]。而控释肥料能够根据作物不同生育期的养分需求延缓或者控制养分释放的速率,最大限度的提高作物对养分的吸收,有效的解决了氮肥利用率低和氮素损失严重的问题,且一次性施用简化了施肥技术,能够达到增产增效、省工省时和减少环境污染等目的[14,15,16,17]。目前,国内在水稻控释肥方面的研究报道已较多,郑圣先等[13]田间试验报道,控释肥处理水稻的氮素吸收规律呈三次曲线,控释肥的氮素释放曲线呈二次曲线,其养分释放速率能与水稻各生育期的养分需求相匹配,整个生育期水稻的累积吸收氮量低于控释肥的累积释放氮量,控释肥处理水稻氮肥利用率高达72.3%,比普通尿素高出36.5个百分点。郑圣先等[18]15N示踪试验表明,与尿素相比,施用控释氮肥能明显降低氨挥发损失54.0%、淋溶损失32.5%和硝化-反硝化损失94.1%,增加水稻植株氮素吸收量87.8%,增加氮肥利用率32.2个百分点。张民等[19]和蒋曦龙等[20]研究发现,普通尿素转化为可供水稻吸收利用的氮素需要一段时间,肥料氮素释放应早于水稻氮素吸收。水稻生长前期普通尿素处理水稻的氮素吸收量显著高于控释尿素处理,而水稻生长中后期则相反,控释尿素处理水稻不同时期氮素吸收量与其氮素累积释放量均达到极显著正相关。徐明岗等[21]报道,控释氮肥释放速率的快慢对水稻生长发育有明显影响,肥料释放过程与水稻吸氮过程基本同步的控释肥才能满足水稻生产需求。唐拴虎等[22]认为,水稻一次性施用控释肥增产的主要机理是显著提高了氮素养分供应,明显促进了水稻根系生长发育和根系活力,延缓水稻生长后期的衰老进程,并显著增强了水稻抗倒伏能力。国外研究也表明,控释尿素的氮素释放能够与水稻的氮素需求相匹配。与普通尿素相比,控释尿素显著增加了水稻产量以及秸秆和籽粒的氮素积累量,但受环境因素的影响(如土壤温度和水分),并不是所有控释尿素都能增加产量和氮素积累量[23,24]。控释尿素施入稻田后,肥料氮素释放与水稻氮素需求相匹配时,才能增加氮素吸收利用,进而减小氮素损失。因此,明确水稻生育期对氮素的吸收利用特性和控释尿素施入稻田后氮素的去向,才能更好的发挥控释尿素的增效作用。【本研究切入点】尽管有关控释尿素能够增加水稻产量,提高氮肥利用率和减少氮素损失已有很多报道[22,25-26],但大多采用田间试验的表观指标分析,而应用稳定性同位素15N标记控释尿素的技术并定量化研究其在稻田土壤的行为和去向有利于进一步揭示控释尿素的增产增效机制。【拟解决的关键问题】本研究在前人研究的基础上,通过设置稳定性同位素15N微区试验,研究控释尿素氮在水稻主要生育时期不同器官的积累、分配和转运特征,及其在稻田的去向和肥料氮利用率,挖掘控释尿素氮肥利用潜力和减少氮素损失的作用,定量化研究控释尿素和普通尿素中氮素在土壤-植物系统中的转化与去向。1 材料与方法
1.1 试验地点
试验于2014年在湖北省武穴市梅川镇从政村(30°07′38″ N,115°35′42″ E)进行,供试土壤为花岗片麻岩发育的水稻土。供试田块前茬为早稻,基础土壤(0—20 cm土层)理化性质:pH 6.3、有机质40.2 g·kg-1、全氮2.2 g·kg-1、铵态氮17.3 mg·kg-1、硝态氮3.2 mg·kg-1、速效磷8.12 mg·kg-1、速效钾149.5 mg·kg-1。1.2 试验设计
田间微区试验采用最优氮、磷、钾配合施用,单因素随机区组设计,设3个处理:(1)不施氮肥(CK);(2)15N标记的粉末状普通尿素(购于上海化工研究院,丰度为10%,含氮46%,以U表示);(3)15N标记的控释尿素(购于上海化工研究院的4 mm 15N大粒尿素,经南京土壤研究所研制出的包膜配方,应用流化床包衣设备生产出15N标记的水基聚合物包膜控释尿素,丰度为10%,含氮41.4%,以CRU表示),各处理3次重复。微区由铁皮框制成,面积1 m×1 m,高0.5 m,埋入土壤0.2 m深。(2)和(3)处理氮肥用量均按纯N 16.5 g·m-2,3个处理磷、钾肥的用量分别按P2O5 7.5 g·m-2和K2O 7.5 g·m-2。磷肥为过磷酸钙(含P2O5 12%),钾肥为氯化钾(含K2O 60%)。各处理氮、磷、钾肥均于7月29日水稻移栽前作基肥一次性撒施,并立即用手混合于土中。供试作物为晚稻,品种为丰源优299,属三系杂交中熟晚籼。采用移栽方式种植,于6月27日播种育秧,7月29日移栽,移栽密度为30穴/m2,每穴3株,10月29日收获。1.3 样品的采集与测定
土壤基础养分采用常规方法进行测定[27]。分别于拔节孕穗期(9月2日)、抽穗期(9月15日)、开花期(9月29日)、灌浆期(10月13日)取活体植株(除落叶外)样1穴,齐地收割,并分为茎、鞘、叶、籽粒和穗轴5部分;成熟期(10月29日)取活体植株样3穴,连根拔起,并分为根、茎、鞘、叶、籽粒和穗轴6部分。各处理齐地收割剩余的23穴水稻植株并测产。各部位植物样品65℃烘干,称重,粉碎,过60目筛后测定全氮含量和15N丰度。于收获期取0—20、20—40、40—60和60—80 cm土层土样,烘干,磨碎,过100目筛后测定全氮和15N丰度。土壤和植株全氮含量及15N丰度采用元素分析仪与Isoprime-100稳定同位素质谱仪联用测定。1.4 计算方法[28 ,29 ]
15N原子百分超(%)=样品或15N标记肥料的15N丰度-15N天然丰度;植株氮素来自15N标记肥料的百分比Ndff(%)=样品的15N原子百分超/标记肥料的15N原子百分超×100;
植株氮素来自土壤的百分比Ndfs(%)=1-Ndff%;
某一植株组织或器官的15N积累量=该器官氮积累量×该组织或器官的Ndff%;
15N转运量=开花期茎、鞘、叶和穗轴15N积累量之和-灌浆期(或成熟期)茎、鞘、叶和穗轴15N积累量之和;
15N转运效率= 15N转运量/开花期茎、鞘、叶和穗轴15N积累量之和;
15N转运量对籽粒的贡献率=15N转运量/成熟期籽粒15N积累量;
激发氮量(g·m-2)=施氮处理植株吸收来自土壤的氮-不施氮处理植株吸氮量;
15N利用率(%)=植株地上部15N积累量/投入肥料15N量×100;
15N土壤残留量=土样干重×土壤全氮含量(%)×土壤15N原子百分超×100;
15N土壤残留率(%)= 15N土壤残留量/投入肥料15N量×100;
15N根系吸收利用率=根系15N积累量/投入肥料15N量×100;
15N损失率(%)=100-15N总回收率=100-15N利用率-15N土壤残留率- 15N根系吸收利用率。
1.5 数据统计分析
所有数据均由MS Excel 2016和DPS(v7.05)数据处理系统分析处理,采用LSD法检验差异显著性(P<0.05)。2 结果
2.1 水稻植株各部位干物质积累及分配
不同肥料N处理水稻植株各部位在不同生育期的干物质积累量及分配见表1。随生育期的推进,CRU处理水稻植株地上部干物质量增加迅速,至灌浆期达最大值,较U处理增加5.0%。CRU处理茎和鞘的干物质量在开花期达最大值,与U处理相比,茎的干物质量增加13.8%,鞘的干物质量无显著差异;叶的干物质量在抽穗期达最大值且较U处理无显著差异,随后由于营养器官干物质向籽粒的不断转移而降低;穗(籽粒+穗轴)的干物质量从孕穗期开始不断增加,成熟期达最大值,开花前显著大于U处理,开花后二者差异逐渐缩小,开花期较U处理增加6.2%。成熟期,与U处理相比,CRU处理茎和鞘的干物质量分别增加17.3%和13.2%,穗和地上部干物质量分别增加3.5%和3.7%,但统计上差异不显著,而叶的干物质量降低14.6%。Table 1
表1
表1不同生育期水稻植株各部位干物质积累及分配
Table 1Dry matter accumulation and distribution of different parts of rice plants at different growth stages
| 处理 Treatment | 取样时期 Sampling stage | 干物质量 Dry matter (g·m-2) | ||||||
|---|---|---|---|---|---|---|---|---|
| 茎 Stem | 鞘 Sheath | 叶 Leaf | 籽粒 Grain | 穗轴 Cob | 穗 Panicle | 合计 Total | ||
| U | 孕穗期 Booting | 32.45c (7.1) | 179.52c (39.3) | 219.33b (48.0) | — | — | 25.97e (5.7) | 457.27d |
| 抽穗期 Heading | 128.47b (15.4) | 248.91b (29.8) | 282.95a (33.9) | 127.24d (15.2) | 47.42c (5.7) | 174.66d (20.9) | 834.99c | |
| 开花期 Anthesis | 242.49a (17.6) | 301.50a (21.9) | 265.28a (19.3) | 518.20c (37.6) | 50.13c (3.6) | 568.33c (41.2) | 1377.60b | |
| 灌浆期 Filling | 143.61b (9.7) | 253.03b (17.1) | 196.15bc (13.3) | 827.28b (55.9) | 60.14b (4.1) | 887.42b (60.0) | 1480.21a | |
| 成熟期 Maturity | 130.89b (8.9) | 196.16c (13.3) | 191.73c (13.0) | 892.38a (60.4) | 65.95a (4.5) | 958.33a (64.9) | 1477.11a | |
| CRU | 孕穗期 Booting | 51.22d* (8.3) | 283.40a* (46.2) | 244.82b* (39.9) | — | — | 34.11e* (5.6) | 613.55d* |
| 抽穗期 Heading | 201.80b* (21.1) | 265.89a* (27.8) | 285.74a (29.9) | 156.57d* (16.4) | 45.84c (4.8) | 202.41d* (21.2) | 955.84c* | |
| 开花期 Anthesis | 275.86a* (19.2) | 289.80a (20.2) | 266.67ab (18.6) | 548.43c* (38.2) | 54.98b* (3.8) | 603.41c* (42.0) | 1435.74b | |
| 灌浆期 Filling | 142.73c (9.2) | 282.71a* (18.2) | 209.83c* (13.5) | 858.15b (55.2) | 60.56a (3.9) | 918.71b (59.1) | 1553.98a* | |
| 成熟期 Maturity | 153.57c* (10.0) | 222.08b* (14.5) | 163.72d* (10.7) | 930.62a (60.8) | 61.17a* (4.0) | 991.79a (64.8) | 1531.16a | |
新窗口打开
随生育期的推进,CRU处理水稻植株茎的干物质分配比例从孕穗期开始先逐渐上升,抽穗至开花期达最大值,分别较U处理提高1.6—5.7个百分点,随后逐渐下降;鞘的干物质分配比例在孕穗期达最大值且较U处理提高6.9个百分点,随后逐渐下降;而穗的干物质分配比例从孕穗期开始不断上升,成熟期达最大值,但较U处理无显著差异。成熟期,CRU处理水稻茎、鞘、叶和穗的干物质分配比例分别为10.0%、14.5%、10.7%和64.8%;与U处理相比,CRU处理茎和鞘的干物质分配比例分别增加1.1和1.2个百分点,穗无显著增加,而叶则降低2.3个百分点。
2.2 水稻植株各部位15N积累及分配
w水稻植株各部位在不同生育期的15N积累量与干物质变化趋势一致,见表2。随生育期的推进,CRU处理水稻植株茎和鞘的15N积累量逐渐增加,开花期达最大值,分别较U处理增加62.5%和25.5%,随后由于营养器官的15N向籽粒的不断转运而降低。由于叶的衰老脱落,CRU处理叶的15N积累量在孕穗期达最大值且较U处理增加13.4%,随后逐渐下降,成熟期达最小值。而CRU处理穗的15N积累量从孕穗期开始不断增加,成熟期达最大值,各生育期均较U处理显著增加。成熟期,CRU处理水稻茎、鞘、叶、穗和地上部15N积累量分别为0.35、0.48、0.39、3.30和4.52 g·m-2,其中水稻茎、鞘、穗和地上部15N积累量分别比U处理增加25.0%、20.0%、15.8%和13.3%,而水稻叶的15N积累量则比U处理降低15.2%。Table 2
表2
表2不同生育期水稻植株各部位15N积累及分配
Table 215N accumulation and distribution of different parts of rice plants at different growth stages
| 处理 Treatment | 取样时期 Sampling stage | 15N积累量 15N accumulation (g·m-2) | ||||||
|---|---|---|---|---|---|---|---|---|
| 茎Stem | 鞘Sheath | 叶Leaf | 籽粒Grain | 穗轴Cob | 穗Panicle | 合计Total | ||
| U | 孕穗期 Booting | 0.08c (2.4) | 0.71b (21.1) | 2.46a (73.2) | — | — | 0.11e (3.3) | 3.36c |
| 抽穗期 Heading | 0.22b (5.5) | 0.83b (20.9) | 2.47a (62.2) | 0.34d (8.6) | 0.11a (2.8) | 0.45d (11.4) | 3.97b | |
| 开花期 Anthesis | 0.32a (6.2) | 0.98a (19.1) | 2.10b (40.9) | 1.61c (31.4) | 0.12a (2.3) | 1.73c (33.7) | 5.13a | |
| 灌浆期 Filling | 0.12c (3.5) | 0.54c (15.7) | 0.62c (18.1) | 2.07b (60.3) | 0.08b (2.3) | 2.15b (62.6) | 3.43c | |
| 成熟期 Maturity | 0.28a (7.0) | 0.40d (10.0) | 0.46d (11.5) | 2.77a (69.4) | 0.08b (2.0) | 2.85a (71.4) | 3.99b | |
| CRU | 孕穗期 Booting | 0.14c* (3.3) | 1.16ab* (27.4) | 2.79a* (65.8) | — | — | 0.15e* (3.5) | 4.24b* |
| 抽穗期 Heading | 0.37b* (7.8) | 1.12b* (23.7) | 2.51b (53.2) | 0.54d* (11.4) | 0.18b* (3.8) | 0.72d* (15.2) | 4.72b* | |
| 开花期 Anthesis | 0.52a* (8.3) | 1.23a* (19.7) | 2.04c (32.7) | 2.23c* (35.7) | 0.22a* (3.5) | 2.45c* (39.2) | 6.24a* | |
| 灌浆期 Filling | 0.18c* (3.8) | 0.80c* (16.8) | 0.93d* (19.5) | 2.74b* (57.4) | 0.12c* (2.5) | 2.86b* (59.9) | 4.77b* | |
| 成熟期 Maturity | 0.35b* (7.7) | 0.48d* (10.6) | 0.39e* (8.6) | 3.21a* (71.0) | 0.09d* (2.0) | 3.30a* (73.0) | 4.52b* | |
新窗口打开
水稻开花期之前吸收的15N主要分配在叶中,其次是鞘,再次是茎。开花期后,随着15N从营养器官向穗中转移,茎、鞘和叶中的15N分配比例逐渐降低,而穗中的15N分配比例不断增加。开花期,与U处理相比,CRU处理水稻茎、鞘和穗中15N的分配比例分别提高2.1、0.6和5.5个百分点,叶则降低8.2个百分点。成熟期,CRU处理水稻茎、鞘、叶和穗中15N的分配比例分别为7.7%、10.6%、8.6%和73.0%,其中茎、鞘和穗中15N的分配比例分别比U处理提高0.7、0.6和1.6个百分点,叶则降低2.9个百分点。
2.3 水稻花后各阶段干物质及15N的转运
从表3可知,开花期到灌浆期,水稻干物质和15N的转运量、转运效率和对籽粒的贡献率表现为CRU处理小于U处理,CRU处理水稻干物质和15N转运量分别为191.50和1.97 g·m-2,分别比U处理降低了7.3%和8.4%;干物质和15N转运率分别为21.6%和49.3%,分别比U处理减少了2.4和11.8个百分点;干物质和15N转运对籽粒的贡献率分别为20.6%和61.5%,分别比U处理降低了2.5和16.2个百分点;这显然是由于U处理水稻生育中、后期养分供应不足,植株加速衰老,营养物质迅速从营养器官向生殖器官中转移所致,而CRU处理延缓了水稻生育中、后期植株的衰老进程。而开花期到成熟期,CRU处理水稻干物质转运量、转运率和对籽粒的贡献率分别为286.78 g·m-2、32.3%和30.8%,水稻15N转运量、转运率和对籽粒的贡献率分别为2.69 g·m-2、67.2%和83.8%,与U处理相比略有增加,但统计上差异不显著。可见,开花期到灌浆期,CRU处理水稻干物质及氮素转运虽然较U处理缓慢,但灌浆期到成熟期CRU处理养分供应充足,促进了水稻籽粒的灌浆速率,促进了籽粒中干物质的积累及氮素的同化和营养器官中养分向籽粒的迅速转运,而U处理水稻灌浆期后植株已衰老,存在老叶脱落或水稻植株自身氨挥发损失。Table 3
表3
表3水稻花后各阶段干物质和15N的转运量、转运效率和对籽粒的贡献率
Table 3Dry matter and 15N translocation, translocation efficiency and contribution efficiency to grain of rice at different growth stages after anthesis
| 处理 Treatment | 时期 Stages | 干物质转运量 Day matter translocation (g·m-2) | 转运效率 Translocation efficiency (%) | 对籽粒的贡献率 Contribution efficiency to grain (%) | 15N转运量 15N translocation (g·m-2) | 15N转运效率 15N translocation efficiency (%) | 对籽粒的贡献率 Contribution efficiency to grain (%) |
|---|---|---|---|---|---|---|---|
| U | 开花期—灌浆期 Anthesis-Filling | 206.47 | 24.0 | 23.1 | 2.15 | 61.1 | 77.7 |
| 开花期—成熟期 Anthesis-Maturity | 274.66 | 32.0 | 30.8 | 2.30 | 65.3 | 83.0 | |
| CRU | 开花期—灌浆期 Anthesis-Filling | 191.50 | 21.6 | 20.6 | 1.97 | 49.3 | 61.5 |
| 开花期—成熟期 Anthesis-Maturity | 286.78 | 32.3 | 30.8 | 2.69 | 67.2 | 83.8 |
新窗口打开
2.4 水稻氮素吸收来源
水稻生育期内,不考虑干湿沉降和灌溉所带入的氮素,水稻吸收的氮素主要来自于肥料的氮素和土壤的氮素。表4结果表明,水稻产量和植株吸氮量因施用氮肥而显著增加。等氮量下,与U处理相比,CRU处理水稻产量和植株吸氮量略有增加,但统计上差异不显著。由于土壤中存在一定量的氮素,CK处理在不施氮条件下,仍可吸收土壤中的氮素,整个水稻季共吸收土壤氮素9.09 g·m-2。无论施用控释尿素还是普通尿素,土壤氮素均是水稻生长发育所需氮素的主要来源,水稻生育期吸收土壤中的氮素占植株吸氮量的70%以上。与U处理相比,CRU处理增加了水稻吸收来自肥料的氮素达13.3%,Ndff提高2.9个百分点;而与CRU处理相比,U处理促进了水稻吸收来自土壤的氮素达3.1%,Ndfs提高2.9个百分点,其激发氮量也提高了12.5%。Table 4
表4
表4水稻吸收肥料氮和土壤氮的比例
Table 4The percentage of 15N derived from fertilizer (Ndff) and soil (Ndfs) of rice
| 处理 Treatment | 产量 Yield (g·m-2) | 植株吸氮量 Total N uptake (g·m-2) | 来自肥料的氮 N from fertilizer (g·m-2) | 较U处理增加Increase(%) | Ndff (%) | 较U处理增加百分点Increase percentage points | 来自土壤的氮 N from soil (g·m-2) | 较CRU处理增加Increase(%) | Ndfs (%) | 较CRU处理增加百分点Increase percentage points | 激发氮量 Stimulated N (g·m-2) | 较CRU处理增加Increase(%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CK | 768.35b | 9.09b | — | — | — | — | 9.09b | — | 100.0a | — | — | — |
| U | 892.38a | 16.50a | 3.99 | — | 24.2 | — | 12.51a | 3.1 | 75.8b | 2.9 | 3.42 | 12.5 |
| CRU | 930.62a | 16.65a | 4.52 | 13.3 | 27.1 | 2.9 | 12.13a | — | 72.9b | — | 3.04 | — |
新窗口打开
2.5 15N平衡与去向
施入稻田土壤的肥料氮主要有3种去向:植物吸收、土壤残留和各种途径的氮素损失(如氨挥发、淋溶和硝化反硝化)。另外,有少量残存于收获后的根茬中。表5为水稻收获后,施入稻田土壤的肥料氮在土壤—植物系统中的去向。从氮肥的吸收利用来看,CRU处理水稻15N吸收量为4.52 g·m-2,比U处理增加了13.3%;CRU处理水稻差减法利用率和15N示踪法利用率分别为45.8%和27.4%,分别比U处理提高了0.9和3.2个百分点。从土壤残留来看,CRU处理土壤15N残留量为1.65 g·m-2,比U处理提高10.0%;CRU处理土壤15N残留率为10.0%,比U处理提高0.9个百分点。从氮素的总回收来看,CRU处理15N总回收量为6.43 g·m-2,比U处理提高11.4%;15N总回收率为39.0%,比U处理提高4.0个百分点。从表5还可见,U处理的15N损失量为10.7 g·m-2,15N损失率高达65.0%,CRU处理与其相比损失量降低5.6%,损失率降低4.0个百分点。Table 5
表5
表5稻田15N平衡与去向
Table 515N balance and fate in paddy rice
| 处理 Treatment | 施氮量 N applied (g·m-2) | 15N吸收量 15N uptake (g·m-2) | 氮肥利用率 N recovery efficiency (%) | 土壤残留 Residual in soil | 根系吸收 Uptake by root | 总回收 Total recovery | 氮损失 N loss | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 差减法 Difference method | 15N示踪法 15N tracer method | 15N 残留量 15N residue | 15N残留率 15N residue rate (%) | 15N吸收量 15N uptake (g·m-2) | 15N利用率 15N use efficiency (%) | 15N总 回收量 15N recovery | 15N总 回收率 15N recovery rate (%) | 15N 损失量 15N loss (g·m-2) | 15N损失率15N loss rate (%) | |||
| U | 16.5 | 3.99b | 44.9a | 24.2b | 1.50b | 9.1b | 0.28a | 1.7a | 5.77b | 35.0b | 10.7a | 65.0a |
| CRU | 16.5 | 4.52a | 45.8a | 27.4a | 1.65a | 10.0a | 0.26a | 1.6a | 6.43a | 39.0a | 10.1a | 61.0a |
新窗口打开
2.6 肥料15N在土壤剖面的残留分布
由图1可知,肥料氮在土壤中的残留量随土层深度的增加而显著下降。在等氮量下,普通尿素和控释尿素在0—80 cm土层中(表5)的15N残留量分别为1.50和1.65 g·m-2,残留率分别为9.1%和10.0%,可见施用控释尿素显著增加了0—80 cm土层中15N总残留量和总残留率。但在不同土层中两种肥料15N的残留存在一定的差异。水稻收获后,肥料15N主要残留在0—20 cm土层中,约占总残留量的78%,可为下季作物吸收再利用,提高氮素的利用率。其次是20—40 cm和40—60 cm土层,这两层土壤中肥料15N残留量相近,约占总残留量的19%左右。而在60 cm以下土层,仍有微量的肥料15N残留,占总残留量的4%以下。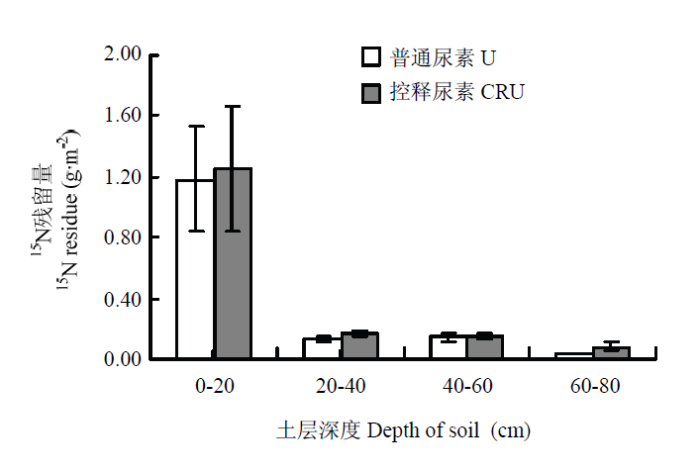 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1肥料15N在土壤剖面上的残留
-->Fig. 1Residual of fertilizer 15N in soil profile
-->
3 讨论
近年来,水基聚合物包衣控释肥料因其包膜材料来源广泛、价格低廉和环境友好等优点,已成为广泛研究的商品化控释肥料之一[30,31]。但由于其膜强度不足、亲水性较高而耐水性较差等,导致了养分的控释效果不够理想,限制了其大规模应用[32]。本研究应用南京土壤研究所基于生物炭改性研制成的水基聚合物聚丙烯酸酯包膜尿素,以期提高膜材料的疏水性和改善其力学性质,同时优化其包膜肥料的控释性能。结果显示,稻田施用水基聚合物包膜控释尿素与普通尿素后,虽然产量差异不显著(表4),但水稻植株干物质和15N积累、分配及转运均存在本质区别,控释尿素在水稻各生育期养分供应充足,为水稻营养生长和生殖生长提供了良好的营养基础,并在提高水稻15N利用率(提高3.2个百分点)和减少15N损失率(降低4.0个百分点)方面有一定效果(表5)。刘德林等[33]研究表明,施用控释尿素可显著增加水稻产量25.5%,使15N利用率高达73.8%,比普通尿素高34.8个百分点,同时减少15N损失率34.3个百分点。本研究在增加水稻产量,提高水稻15N利用率和减少15N损失效果方面与前人结果有一定差异,分析原因可能有两点:①氮肥去向和损失途径受气候、品种和施肥量等因素的影响,存在着很大的变异性[29]。包膜型控释肥养分释放受土壤温度和水分影响很大[24,34],由于淹水稻田,水分充足,故决定其养分释放的主要因素是温度[35]。本研究为晚稻季,水稻生长于7月底—10月底,前期(7—8月)气温较高,控释尿素施入稻田后,高温促进了其养分的快速释放,水稻移栽后到返青期吸氮量少,田面水中有大量NH4+存在,导致氨挥发损失严重[13,16,36]。②15N标记控释尿素包膜材料、释放性能和制备方法不同。15N示踪技术在聚合物包膜控释肥料的研究非常有限,缺乏低成本且成熟完善的15N标记包膜控释肥料技术是主要原因[35,37]。本研究所用15N标记控释尿素为南京土壤研究所基于生物炭改性水基聚合物聚丙烯酸酯包膜控释尿素,由于水基包膜具有较高的亲水性,其控释性能可能还不够理想,田间实际释放期太短[16-17,31];而刘德林等[33]和郑圣先等[18]所用15N标记控释尿素为湖南省农业科学院通过添加魔芋淀粉等粘合剂将粉状15N标记尿素用模具挤压成型,用郑圣先等研制出的包膜配方,在流化床内用单管喷嘴细微雾状喷涂大粒尿素生产出的15N标记控释尿素。有研究表明,添加了粘合剂的肥料颗粒与普通尿素相比,物理结构发生了变化,肥料水解性能大大降低,从而影响了肥料养分的释放[37]。尽管15N同位素示踪控释肥的制备还有待改进,但是在研究水稻生育期对氮素的吸收利用特性和控释尿素施入稻田后氮素的去向等问题,以及解释控释肥增产增效、减少氮素损失的机理上仍是一种有效的方法,为控释肥料的研发、生产及推广应用等方面提供了理论依据。15N示踪技术是目前公认的研究氮素行为和其环境行为最准确的方法之一,它可以准确的追踪肥料氮施入土壤后的转化与去向,区分作物吸收的氮素是来自于肥料还是来自于土壤,但其所测定的氮肥利用率数值不尽相同。一般认为,标记15N示踪法测得的利用率低于差减法计算的表观利用率[29]。本研究结果表明,差值法计算的氮肥表观利用率比15N示踪法测得的利用率平均高出19.6个百分点,这与郑圣先等[13]的报道一致。有研究表明,“激发效应”的存在,是示踪法利用率低于差减法计算的表观利用率的原因之一[38]。在15N标记肥料试验中,经常出现施氮处理的植株比不施氮处理吸收更多的土壤氮,这种土壤释放氮随施肥量的增加而增加的现象,被称作“激发效应”。本研究结果表明,施用普通尿素后,水稻激发氮量比控释尿素高12.5%。刘德林等[33]报道,水稻控释氮肥的激发效应比普通尿素强,但在第四纪红土发育的水稻土上的激发作用则表现得比较迟,这可能与控释肥料的缓慢释放有关。土壤氮素是作物吸收氮的主要来源,本试验中,成熟期水稻吸收土壤氮的比例为72.8%—75.8%,吸收肥料氮的比例为24.2%—27.2%,这与晏娟等[28]的报道基本一致。施用控释尿素后,土壤15N残留率为10.0%,比普通尿素处理提高了0.9个百分点。水稻收获后,肥料15N主要残留在0—20 cm土层中,约占总残留量的78%,下层残留量逐渐减少,但0—80 cm土层中均有分布。朱培立等[39]报道,氮肥施入土壤后,有相当大一部分残留在土壤中,其利用率仅2.5%。而残留肥料氮本身的矿化势与激发效应呈正相关,残留在土壤中的肥料氮,在追加无机氮源后,均产生正激发效应。土壤氮素残留能够补偿土壤氮库,并为下季作物吸收再利用,提高氮素的利用率[29]。因此,在水稻生产中,施用控释尿素应充分考虑上季作物残留在土壤中的氮以及由施肥而产生的激发效应,适当降低控释尿素的施用量,增加对土壤残留氮素的吸收利用,以降低生产成本,达到增产增效,提高肥料的利用率和减少环境污染的目的,助力“化肥零增长”的实现。
4 结论
4.1 成熟期,控释尿素(CRU)处理水稻茎和鞘的干物质量比普通尿素(U)处理分别增加了17.3%和13.2%,其干物质分配比例分别增加1.1和1.2个百分点;CRU处理水稻茎、鞘、穗和地上部15N积累量分别比U处理增加25.0%、20.0%、15.8%和13.3%;施用CRU促进了水稻籽粒的灌浆速率,促进了籽粒中干物质的积累及氮素的同化和营养器官中养分向籽粒的迅速转运。4.2 土壤氮素是水稻生长发育所需氮素的主要来源,水稻生育期吸收氮素来自土壤的氮素约占70%以上。与U处理相比,CRU处理增加了水稻吸收来自肥料的氮素达13.3%,Ndff提高2.9个百分点。
4.3 等氮量下,与U处理相比,CRU处理水稻产量和植株吸氮量略有增加,但统计上差异不显著。CRU处理使水稻15N利用率提高了3.2个百分点,15N土壤残留率增加了0.9个百分点,15N总回收率提高了4个百分点,15N损失率降低了4个百分点。
4.4 肥料氮在土壤中的残留量随土层深度的增加而显著下降。水稻收获后,肥料15N主要残留在0—20 cm土层中,约占总残留量的78%,可为下季作物吸收再利用,提高氮素的利用率。
The authors have declared that no competing interests exist.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | |
| [2] | |
| [3] | . |
| [4] | . . |
| [5] | . |
| [6] | . |
| [7] | . |
| [8] | . . |
| [9] | . . |
| [10] | . |
| [11] | . |
| [12] | |
| [13] | . . |
| [14] | . |
| [15] | . . |
| [16] | . |
| [17] | . |
| [18] | . . |
| [19] | . . |
| [20] | . . |
| [21] | . . |
| [22] | . . |
| [23] | . |
| [24] | . |
| [25] | . |
| [26] | . . |
| [27] | |
| [28] | . . |
| [29] | . . |
| [30] | . . |
| [31] | . |
| [32] | . . |
| [33] | . . |
| [34] | . |
| [35] | . . |
| [36] | . |
| [37] | |
| [38] | . . |
| [39] | . . |

{kind=link}
{kind=link}