0 引言
【研究意义】水稻是世界上最重要的粮食作物之一,也是基础研究的模式生物[1]。胁迫可分为生物胁迫和非生物胁迫,常见的非生物胁迫包括冷、热、淹、光照、旱、盐等,水稻在胁迫条件下的生长发育和产量都会受到影响,了解水稻胁迫反应应答机理具有重要意义。【前人研究进展】水稻是最早完成基因组测序的作物,中国科学家最早完成了超级杂交稻父本9311的基因组测序[2],在水稻基因组以及多种重要植物的基因组测序工作中发挥了引领作用。目前,在建立水稻全基因组基因芯片平台[3]及二代测序技术的基础上,已经积累了丰富的不同时空条件下的转录组信息[4,5,6]。在水稻基础研究的不同阶段,支撑性平台技术都发挥了重要的作用,如20世纪末的分子标记技术、遗传转化技术、基因组文库和cDNA文库构建技术、全长cDNA文库,以及突变体库、基因编辑技术等[7],正是在这些平台技术的支持下,水稻基础研究得以飞速发展,为解析抗病、高产、光合、株型等机理做出了重要贡献[8]。蛋白质是生命活动的功能执行分子,是基因功能的主要体现者,了解每个蛋白质的状态是生物学家今后面临的挑战。借助高灵敏度的质谱技术,现在已经可以从人的多个组织样品中累积鉴定到17 000个以上的蛋白质[9]。为了特异性地检测每个蛋白质,瑞典科学家UHLÉN实验室制备了3万多个针对人的蛋白质的抗体资源库[10]。可以说,质谱和抗体是蛋白质组学研究的二大利器[11,12]。在水稻中,提出了基于抗体的蛋白质组学策略,目前已经制备了2 000多个水稻蛋白质特异的抗体[13]。就像在人的蛋白质研究中一样,抗体也是水稻基础研究的重要资源,可广泛应用于蛋白质定位、蛋白质-蛋白质相互作用、蛋白质-核酸相互作用、以及蛋白质的修饰、降解等。与DNA的分析不同,RNA和蛋白质的表达具有组织、时空特异性,样品的稳定性较低,所以对样品的要求更高。只有可靠的样品,试验结果才有意义。在人的组织病理研究中,样品资源库是专门的课题,如人的前列腺癌样品资源库[14],研究儿童发热与近视所建立的UK biobank[15],研究人类衰老所建立的RPGEH样品库[16],以及鼻咽癌资源库[17]等。2017年,美国及多国科学家联合启动的“人类细胞图谱计划”(Human Cell Atlas,HCA),设想对人体中所有细胞进行分类和测序,系统地描绘人体细胞图谱,并通过这把钥匙来加深对疾病诊断、监测、治疗的了解,可靠的样品是保证项目质量的前提[18]。Facebook创始人马克·扎克伯格出资支持这项计划,并将其与“人类基因组计划”媲美[19]。2018年,浙江大学医学院郭国骥教授团队在Cell发表论文,报道了对小鼠不同生命阶段的近50种器官组织的40余万个细胞进行了系统性的单细胞转录组分析,构建了首个哺乳动物细胞图谱[20]。在这项研究中,最关键的过程之一是如何分离单细胞并保证样品的可靠性。另外,由于蛋白质分子不能在体外进行扩增,所以蛋白质分析中需要的样品量要多于核酸分析。在植物研究中,样品资源库也将是一个重要的基础平台。免疫印迹(western blot,WB)是利用抗体的特异性对混合样品中的目标蛋白质进行检测的技术,同酶联免疫(ELISA)相比,WB具有更好的特异性,结果也更为直观,WB是生物科学领域的基本检测技术,有极为广泛的应用。近年来,在植物基因功能的研究中,引入或敲除特定基因后都需要借助WB检测目标蛋白质的表达,从而确认引入或敲除的效果[21]。WB也可用于病原物的有无及丰度的检测[22]。当然,目前常规的WB分析还涉及比较繁琐的人工操作的过程,但全自动的WB仪器近年来已经开始应用[23]。非生物胁迫是影响水稻生长和产量的重要因素,冷、热、旱、淹、盐等都是常见的逆境,逆境胁迫会引起植物对逆境的反应,包括一系列基因在转录和蛋白质水平的表达变化。河北农业大学生命科学学院分子生物学与生物信息学实验室(Molecular Biology & Bioinformatics Lab,MBB)曾利用WB技术调查了水稻类钙调磷酸酶亚基B蛋白质[24]、PP2Ac类磷酸酶蛋白质[25]、水稻病程相关蛋白质[26]、水稻核糖核酸酶T2蛋白质[27]等在逆境胁迫下的表达,靶向性地揭示了这些蛋白质在逆境胁迫条件下的表达特征。【本研究切入点】水稻是植物研究的模式生物,但系统性构建蛋白质样品资源库目前还属空白,本研究结合水稻乃至植物研究的特点,开创性的提出蛋白质样品资源库的概念,并提出了具体的建设流程和评价体系。【拟解决的关键问题】本研究以水稻幼苗为材料,进行冷、热、旱、盐、淹、恒光、恒暗等胁迫处理,在不同时间点取样,提取近300份蛋白质样品,对样品质量进行评价,建立胁迫条件下的水稻蛋白质资源库(RiceS-A300),并应用这个资源库检测了MBB实验室前期报道的一个水稻内参蛋白质HSP82的表达特征。1 材料与方法
1.1 水稻材料
水稻品种:粳稻品种TP309。试验于2017年在河北农业大学稻竹园及MBB实验室进行。1.2 苗期培养
种子30℃浸泡3 d露白,土培用蛭石土(土壤与蛭石1:1),播种后30℃、光周期L12 h/D12 h培养5 d,进行冷、热、淹和恒光、恒暗处理。水培是将露白后的种子播放在纱网上,30℃、光周期L12 h/D12 h培养5 d,进行旱和盐处理。以30℃、光周期L12 h/D12 h的非胁迫处理材料为对照。1.3 胁迫处理
冷:冷处理为4℃,取材时间点为0、2、4、8、12、18、24、36和48 h。热:热处理为44℃和48℃,取材时间点为0、2、4、8、12、18、24、36和48 h。淹:将水稻幼苗连同花盆浸没于水中,叶尖离水面约30—40 cm,取材时间点为0 h、4 h、8 h、12 h、24 h、2 d、3 d、5 d和7 d。光照:恒光处理为24 h光照,恒暗处理为24 h黑暗,取材时间点为0、1、2、3、4、5、6、7和8 d,以L12 h/D12 h平行培养的材料为对照。旱:对水培培养5 d的水稻幼苗用PEG6000(20%)处理,取材时间点为0 h、4 h、8 h、12 h、24 h、2 d、3 d、5 d和7 d,以不加PEG的幼苗材料为对照。盐:对水培培养5 d的水稻幼苗用NaCl(0.2 mol·L-1)处理,取材时间点为0 h、4 h、8 h、12 h、24 h、2 d、3 d、5 d和7 d,以不加NaCl的幼苗材料为对照。1.4 株高和鲜重测量
用普通直尺测量地上部长度,每个样品点测量5株以上,计算平均值和方差。鲜重测量:用千分之一天平称量水稻整株幼苗的鲜重(包括地上部和地下部),每个样品点测量5株以上,计算平均值和方差。1.5 主要试剂
NaCl、PEG6000、甘氨酸和溴酚蓝购自上海生工;Tris购自Sigma;甘油和NaF购自天津福晨;SDS、β-巯基乙醇、Arc、Bis和AP购自Biotopped;奶粉、HCl、吐温20、TEMED、考马斯亮蓝R250、PMSF、EDTANa2和DTT购自索莱宝;甲醇购自保定天成;冰醋酸购自天津天大化工;HSP82抗体和羊抗鼠二抗购自华大蛋白;ECL化学发光液购自百智生物;PVDF膜购自Millipore;BCA蛋白质检测试剂盒购自宝生物。
1.6 主要仪器
低温离心机(Thermo,D-37520);电泳槽Mini PROTEAN Tetra cell(伯乐,552BR),电泳仪(伯乐,Power Pac Basic)、转膜芯(伯乐)、pH计(Mettler-toledo,DELTA 320);化学发光成像仪(赛智,MiniChemi 610);天平(OHAUS,AR3130);微量分光光度计(Eppendorf,BioSpectrometer fluorescence);光照培养箱(宁波赛福,PAX-350B)。1.7 取材
地上部和地下部分别取材,统一按0.200 g装入锡箔纸中,置于冰上,液氮速冻后-80℃保存备用。1.8 蛋白质提取
取出冻存的水稻样品,液氮冷冻后用研钵研磨,加入2 mL的蛋白质提取液,具体步骤参见文献[28]。1.9 用BCA法测定蛋白质浓度
将提取的蛋白质用BCA法测定蛋白质浓度,具体方法按试剂盒(TaKaRa BCA Protein Assay Kit)的说明书进行。1.10 蛋白质的SDS-PAGE分离和考染
用10%的SDS-PAGE分离蛋白质,电压160 V,电泳时间60 min,用考马斯亮蓝染色(染色液:甲醇45 mL、H2O 45 mL、冰乙酸10 mL和考马斯亮蓝R-250 0.25 g),脱色(脱色液:乙醇50 mL、冰乙酸100 mL和H2O 850 mL)后拍照。1.11 WB分析
SDS-PAGE分离时,蛋白质上样体积为10 μL,电泳完成后蛋白质转移到PVDF膜上,按文献[28]描述的过程进行WB检测,所用一抗为抗HSP82抗体[29],二抗为HRP标记的羊抗鼠二抗,检测液为ECL,WB信号用化学发光成像仪检测。为了对不同WB分析的结果进行比较,以苗期5 d的幼苗(地上部+地下部)样品作为共同参照(common reference,CR),点在每个WB的第一个泳道。2 结果
2.1 不同胁迫处理对水稻幼苗表型的影响
在水稻幼苗胁迫的不同时间点拍照,以未经胁迫处理的幼苗材料为对照(图1)。从图1-A可见,冷胁迫18 h,肉眼可见幼苗有失水、萎缩现象,处理1 d后,植株明显矮于对照。从图1-B可见,44℃胁迫18 h后,肉眼可见幼苗发黄、失水、萎缩,植株也明显矮于对照。从图1-C可见,48℃胁迫12 h后,肉眼即可见幼苗发黄、失水、萎缩,胁迫症状比植株44℃胁迫处理更重。从图1-D可见,淹胁迫1—2 d后,肉眼即可见幼苗较对照更高,植株仍能维持直立,胁迫处理3 d后,植株倒伏,5 d后叶片组织明显软化。从图1-E可见,恒暗胁迫1—2 d,即可肉眼看出叶片发黄,3 d以后叶片明显整体变黄,7 d后叶片发生灰暗、萎蔫,植株较对照矮小。从图1-F可见,恒光胁迫3 d内表型变化不明显,4 d后植株较对照矮小,叶片颜色更深。从图1-G可见,PEG胁迫2 d后植株矮于对照,植株也更为纤细、瘦弱。从图1-H可见,盐胁迫处理后植株基本停止生长,至5 d时可见叶尖枯萎。从植株表型可见,在所设立的时间点范围内,不同胁迫对水稻幼苗均有肉眼可见的影响,表型的变化为保证取材的质量奠定了基础。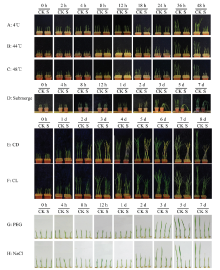 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1水稻幼苗经胁迫处理后的表型图A:4℃冷胁迫,B:44℃热胁迫,C:48℃热胁迫,D:淹胁迫,E:持续黑暗胁迫,F:持续光照胁迫,G:旱(20% PEG6000)胁迫,H:0.2 mol·L-1 NaCl胁迫。A—C取材时间点为0、2、4、8、12、24、36和48 h。D取材时间点为0 h、4 h、8 h、12 h、1 d、2 d、3 d、5 d和7 d。E、F取材时间点为:0 h、1 d、2 d、3 d、4 d、5 d、6 d、7 d和8 d。G、H取材时间点为:0 h、4 h、8 h、12 h、1 d、2 d、3 d、5 d和7 d。CK:正常培养条件,S:胁迫,CD:持续黑暗,CL:持续光照。下同
-->Fig. 1Phenotypic photos of rice seedlings cultivated under stressed conditionsA: Cold stress 4℃, B: Heat stress 44℃, C: Heat stress 48℃, D: Submerge stress, E: Constant dark, F: Constant light, G: 20% PEG6000, H: 0.2 mol·L-1 NaCl. Time points for A-C: 0, 2, 4, 8, 12, 24, 36 and 48 h. Time points for D:0 h, 4 h, 8 h, 12 h, 1 d, 2 d, 3 d, 5 d and 7 d. Time points for E and F: 0 h, 1 d, 2 d, 3 d, 4 d, 5 d, 6 d, 7 d and 8 d. Time points for G and H: 0 h, 4 h, 8 h, 12 h, 1 d, 2 d, 3 d, 5 d and 7 d. CK: Normal culture condition, S: Stressed, CD: Constant dark, CL: Constant light. The same as below
-->
2.2 不同胁迫处理对水稻幼苗株高的影响
株高是评价胁迫应答反应的一个重要指标,调查不同处理在不同时间点的株高(图2)。由图2-A可见,在4℃冷胁迫条件下,水稻幼苗的株高伸长基本停止,二天内即可与对照有显著的区别。由图2-B可见,44℃热胁迫条件下,水稻幼苗的株高仍略有伸长,但在2 d内与对照有显著的区别。由图2-C可见,在48℃热胁迫条件下,水稻幼苗的株高生长停止,与对照有显著的区别。由图2-D可见,淹胁迫能促进株高的伸长,胁迫7 d株高是对照的169%。由图2-E可见,恒暗处理会抑制株高,在第4天株高达到峰值,之后株高持续下降。由图2-F可见,恒光处理对株高也是抑制,但株高可以持续伸长,比较而言,恒光对水稻幼苗的影响小于恒暗。由图2-G可见,PEG处理形成的干旱胁迫对株高有一定的抑制作用。由图2-H可见,盐胁迫对幼苗株高也有比较明显的影响。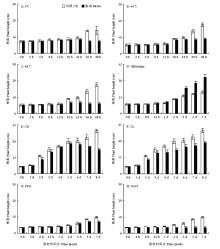 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2不同胁迫处理对水稻幼苗株高的影响
-->Fig. 2Effects on the plant height of rice seedlings under different stressed conditions
-->
2.3 不同胁迫处理对水稻幼苗鲜重的影响
鲜重也是反映水稻植株生长状况的重要指标,调查不同处理在不同时间点的鲜重(图3)。由图3-A可见,4℃冷胁迫,水稻幼苗生长减慢,鲜重下降。由图3-B—图3-C可见,44℃热胁迫,水稻幼苗的鲜重也有下降,但下降的幅度低于48℃热胁迫处理。由图3-D可见,淹胁迫对鲜重的影响不大。由图3-E可见,恒暗处理会显著降低鲜重,与株高类似,鲜重在第4天达到峰值,之后鲜重持续下降。由图3-F可见,恒光处理对鲜重影响不大。由图3-G可见,PEG处理对鲜重有一定影响。由图3-H可见,盐胁迫对幼苗鲜重也有比较明显的影响。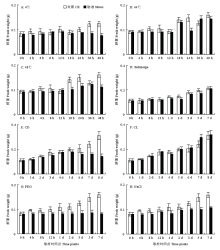 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3不同胁迫处理对水稻幼苗鲜重的影响
-->Fig. 3Effects on the fresh weight of rice seedlings under stressed conditions
-->
此外,曾尝试对根长进行测量,但胁迫处理总体来讲对根长影响不大,且根长的变化幅度较大,不太适合作为胁迫处理的标志性指标。
2.4 不同胁迫处理对水稻幼苗蛋白质含量的影响
为了考察不同胁迫对总蛋白质的影响,提取总蛋白质后进行了SDS-PAGE分离(图4)。不同处理不同时间点的样品均能看到比较典型的考染条带。为了对总蛋白质含量进行定量比较,采集了考染信号(图5)。在对照样品中,由于水稻幼苗的快速生长,单位鲜重样品中的蛋白质含量略呈下降,但3种温度胁迫处理均能提高总蛋白质含量,且蛋白质含量的提高在较短的时间内(2 h)即可检测到,说明总蛋白质含量提高是水稻抵抗温度逆境的一个重要方式。由图5-D可见,淹胁迫对总蛋白质的影响不大。恒暗处理(图5-E)会明显降低总蛋白质的含量,说明光合作用的下降对蛋白质的合成有较为重要的影响。另外,恒光处理(图5-F)、干旱(图5-G)和盐胁迫(图5-H)对总蛋白质含量影响不大。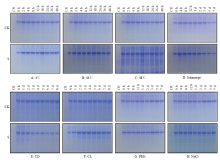 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4水稻幼苗地上部总蛋白质的SDS-PAGE分离及考染检测 SDS-PAGE条件:上样量10 μL,电泳时间60 min,电压160 V,胶浓度10%,电泳槽:Mini PROTEAN Tetra cell
-->Fig. 4Coomassie blue detection of SDS-PAGE separated total protein derived from the shoot of rice seedlings SDS-PAGE conditions: Sample volume 10 μL, Electrophoresis time 60 min, Electrophoretic voltage 160 V, Gel concentration 10%, Electrophoresis apparatus Mini PROTEAN Tetra cell
-->
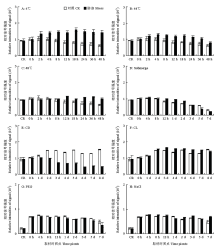 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5胁迫处理后水稻幼苗地上部总蛋白质的定量比较
-->Fig. 5Quantitative comparison of isolated total protein from rice shoots treated by different stress conditions
-->
2.5 蛋白质样品资源库RiceS-A300的建立
在对苗期水稻进行多种胁迫并对表型进行详细调查的基础上,提取了不同时间点样品的总蛋白质,通过考染比较了总蛋白质含量的变化,目前所获得的蛋白质总体积为2 mL,每次WB的用量为10 µL,样品可供200次WB分析,统一编号标注后用9×9的塑料样品盒存放于-20℃冰箱,成为第一个版本的水稻幼苗非生物胁迫处理的蛋白质样品资源库RiceS-A300。由于试验所采用的条件比较规范且容易控制,可根据需要重复扩大样品的规模。预期不同的实验室采用相同的条件获得的蛋白质样品也能获得可重复的试验结果。2.6 基于RiceS-A300调查HSP82蛋白质的表达特征
为了验证RiceS-A300资源库的质量并开展资源库的应用。用WB分析了HSP82的表达特征(图6),采集了WB的信号,绘制成柱状图进行定量分析(图7)。为了比较不同WB之间的结果,以CR的信号为参照,对不同WB进行归一化处理。由图7-A可见,4℃冷胁迫处理对HSP82蛋白质表达的影响不大。由图7-B和图7-C可见,44℃和48℃热胁迫处理,可明显提高HSP82蛋白质的表达,表达的提高在处理2小时即可观察到。由图7-D和5B-E可见,淹胁迫和恒暗处理对HSP82蛋白质的表达影响不大。由图7-F—图7-H可见,恒光、PEG和盐胁迫处理均能提升HSP82蛋白质的表达,但其幅度远低于热胁迫。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6HSP82蛋白质在不同胁迫处理的水稻幼苗地上部的表达特征 WB条件:上样量10 μL,电泳时间60 min,电压160 V,胶浓度10%,电泳槽Mini PROTEAN Tetra cell,转膜时间60 min,转膜电压100 V,膜PVDF,一抗:抗HSP82抗体,二抗:羊抗鼠二抗
-->Fig. 6Expression profiling of HSP82 protein in rice shoot stressed by abiotic conditions WB conditions: Sample volume 10 μL,Electrophoresis time 60 min,Electrophoretic voltage 160 V,Gel concentration 10%,Electrophoresis apparatus Mini PROTEAN Tetra cell. Time of membrane (PVDF) transfer 60 min, voltage 100 V, primary Antibody: anti-HSP82 monoclonal antibody (BPI, AbM51099-31-PU), secondary antibody: goat anti-mouse secondary antibody (BPI,AbP-71003-D-HRP)
-->
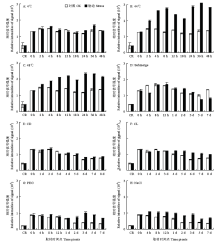 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图7不同胁迫处理条件下水稻幼苗地上部HSP82蛋白质的定量比较
-->Fig. 7Quantitative comparison of HSP82 protein in rice shoots treated by abiotic stress conditions
-->
HSP82是MBB实验室鉴定的水稻蛋白质内参,该蛋白质在多种水稻组织,甚至在其他植物样品中都表达稳定,目前该蛋白质内参已经广为应用。本研究一方面检测了RiceS-A300样品资源库的蛋白质质量,另一方面也在更大范围内检测了HSP82蛋白质的表达特征,是对早期工作的补充和验证[29]。早期试验中由于热处理的时间点不多,也没有采集信号进行定量处理,未能获得热胁迫提高HSP82表达量的结论,本研究中由于有不同温度、不同时间点和定量信号的分析,可以清楚地看到该蛋白质的表达受热胁迫的诱导。
3 讨论
以水稻种子为起点,以水稻幼苗为材料,通过严格控制的条件进行多种胁迫处理,包括冷、热、旱、淹、盐、恒光、恒暗等,调查了胁迫处理对表型、株高和鲜重的影响,在不同的时间点取材,共采集了约300个组织样品,提取总蛋白质,SDS-PAGE分离后进行定量分析和评价后,形成了第一个版本的水稻蛋白质样品资源库RiceS-A300,进而用WB开展了内参蛋白质HSP82的表达特征分析。在生命科学领域,随着高通量分析测定技术的发展,产生的信息资源也呈指数级增长,分析测定技术有时已经不再是主要的研究瓶颈,生物资源却是重要的限制因素。在人类组织样品方面,科研人员对样品的要求也在逐步提高,规范的、相关信息齐备的资源库是重要项目的基本前提。能够为研究群体共享的数据和实物资源具有重要的意义。在水稻中已经积累了全基因组序列和多种时空条件下的转录谱数据,也有一些基于质谱的蛋白质组学等数据资源,在实物资源库方面,也有全基因组基因芯片、分子标记探针、全长cDNA文库、突变体库等。蛋白质样品资源库的建立将有助于推动蛋白质组学的发展,今后这方面的工作一定会得到加强。
结合本研究中所建立的水稻幼苗胁迫处理的蛋白质样品资源库,认为这一类资源库应满足几个条件:一是样品来源要有保障,蛋白质的量总是有限的,样品是需要补充的,所以样品制备条件要能重复,即使在不同实验室间也能重复。本研究从水稻种子起始,在严格控制的生长条件下进行培养和胁迫处理,以便尽可能保证样品的来源可重复。二是资源库样品的相对稳定性,与DNA样品不同,蛋白质样品的贮存要困难的多,但经过100℃处理后的样品在-20℃冰箱贮存1—2年的时间,仍然能够获得可以重复的WB结果。三是样品质量可以跟踪和核查,通过对内在的标志物蛋白质的检测,可检测蛋白质样品的质量。四是资源库可以共享给研究同行,蛋白质资源库的建立对试验的要求很高,经过鉴定的资源库可作为公共平台,共享给研究同行使用,这样可节约社会成本,增加不同实验室间试验结果的可比性。
在建立蛋白质资源库的基础上,检测了HSP82蛋白质在胁迫处理条件下的表达特征,HSP82是MBB实验室在2012年鉴定的一个水稻内参蛋白质[29],该蛋白质在WB检测的几十种水稻样品中都呈现组成型表达,且信号稳定,目前,该内参蛋白质已经被许多水稻乃至其他植物研究同行采用[30,31,32]。本研究证明了该蛋白质在冷、旱、盐、淹、恒光、恒暗等胁迫条件下的幼苗中也相对表达稳定,这一结果扩展了该内参蛋白质的应用范围,而在热胁迫处理过程中其表达量上调,在WB水平证明它确实也是一个热激蛋白。HSP82蛋白质的WB分析结果也可作为检测资源库质量的一个指标。
只要具备了抗体,可以采用本研究建立的蛋白质样品资源库调查目标蛋白质的表达特征,MBB实验室已经进行了多个蛋白激酶、WRKY转录因子、PR等蛋白质的WB分析,发现了一些特定诱导表达的信息(未发表数据)。
为了对不同WB试验间的数据进行比较,本研究中在所有的WB试验中都加入了共同参照样品CR(common reference),通过CR信号调平不同WB间的信号强度就可以进行不同样品信号强度的比较。
在胁迫处理时间的选择上,基本的标准是能够肉眼观察到明显的表型变化,在这样的时间范围内再设置多个取样时间点,以便持续地观察表型,了解表型,了解蛋白质分子表达特征的变化过程。一般来说,在水稻幼苗发生可见的形态变化之前,分子特征会先出现,分子特征出现的时间点、强度变化的幅度都是重要的胁迫应答指标。
本研究所建立的RiceS-A300资源库可为研究同行提供便利,所提出的蛋白质样品资源库建设流程和评价体系也可供植物同行参考。
4 结论
4.1
对水稻幼苗进行了多种非生物逆境处理,调查了胁迫处理过程中9个时间点的幼苗表型、株高、鲜重和总蛋白质含量,发现:淹胁迫促进株高伸长,其他胁迫均对株高伸长有抑制;4℃和恒暗处理降低鲜重,其他胁迫影响不大;3种温度胁迫都能提高总蛋白质含量,恒暗胁迫会降低总蛋白质含量,其他胁迫对总蛋白质含量影响不大。4.2
累计收集了约300份蛋白质样品,建立了第一版的胁迫处理条件下的水稻幼苗蛋白质样品资源库 (RiceS-A300)。4.3
利用RiceS-A300,通过WB调查了HSP82蛋白质的表达特征,发现其在热胁迫下特异诱导表达。4.4
提出并建立了蛋白质样品资源库建设和评价的流程。(责任编辑 李莉)
The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}