0 引言
【研究意义】桃(Prunus persica [L.] Batsch)起源于中国,属于蔷薇科(Rosaceae)李属(Prunus)植物,是世界重要果树树种之一,分布广泛,栽培历史悠久。中国是世界桃的第一生产大国,面积和产量居世界首位。在桃的种植中,存在病虫害严重、着色不良、外观品质差等问题,降低了果实的商品性。果实套袋已成为生产无公害、绿色果品的主要途径和技术措施之一,但套袋对桃果实生长发育及其内在品质也造成一定程度的负面影响[1-4]。【前人研究进展】郭宝林等[5]和陈海江等[6]对燕红、早露蟠桃、北京早艳等桃品种进行套袋试验,结果表明,套袋处理使果实单果重明显减轻。陈建军[7]对9个桃品种套袋,套袋果的可溶性固形物含量均低于对照,而且套袋处理后油桃的固形物含量比普通桃降低幅度更大。沈玉英等[8]和LI等[9]在桃果实套袋上也得到了类似的结果。WANG等[10]发现套袋导致桃果实风味降低,不套袋桃果实的总挥发性成分的含量及C6化合物和酯类要显著高于套袋果实,γ-癸内酯、δ-癸内酯的质量浓度要显著低于不套袋果实。套袋在改善果实外观的同时,对果实内在品质产生一定影响,因此,光照可直接和间接影响果实品质[11-12]。套袋减少了果实着色,降低了果实可溶性固形物含量,使得风味变淡[13]。【本研究切入点】目前,关于桃果实套袋对果实品质和色泽发育的影响,多数是从生理生化角度进行研究,从全基因组表达方面的研究仍鲜见报道。【拟解决的关键问题】本研究以易着色的晚熟桃品种‘秋雪’为材料,研究遮光性套袋与无袋栽培在桃果实转录组中基因表达上的差异,以期探讨遮光性套袋影响桃果实着色和品质的机制,为深入研究桃果实着色的基因调控机制提供理论依据,进一步为优质桃果品的生产提供参考。1 材料与方法
1.1 试验材料
供试桃品种为山东省果树研究所选育的桃新品种‘秋雪’,定植于山东省果树研究所桃种质资源圃内,5年生树,株距为2 m,行距为4 m,主干形整枝,选择干周、冠径及树势等基本一致的作为试验材料。供试果袋(双层外黄内红纸袋,规格为14 cm×18 cm,外层为全木浆纸,内层为石蜡纸)选自青岛小林制袋有限公司。
试验设不套袋(对照)和盛花后50 d套袋 (2014年5月10日),单株小区,随机排列,8月18日沿树冠外围距地面1.5—2 m处随机采收30个果实,3次重复,用液氮迅速冷冻,-80℃保存备用(图1)。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1桃果实的形态特征左:套袋样品;右:对照样品
-->Fig. 1Morphological characteristics of peach fruitsLeft: Bagging peach fruit; Right: CK peach fruit
-->
1.2 样品的RNA提取和检测
采用天根TRNzol Universal总RNA提取试剂(DP424)提取桃果实RNA,步骤参考其说明书。采用Nanodrop、Qubit 2.0、Agilent 2100分别检测RNA样品的纯度、浓度和完整性。1.3 cDNA文库构建和质控文库制备
样品检测合格后,进行文库构建,用带有Oligo(dT)的磁珠富集真核生物mRNA;加入Fragmentation Buffer将mRNA进行随机打断;以mRNA为模板,用六碱基随机引物(random hexamers)合成第一条cDNA链,然后加入缓冲液、dNTPs、RNase H和DNA polymerase I合成第二条cDNA链,利用AMPure XP beads纯化cDNA;纯化的双链cDNA再进行末端修复、加A尾并连接测序接头,然后用AMPure XP beads进行片段大小选择;最后通过PCR富集得到cDNA文库。文库构建完成后,分别使用Qubit2.0和Agilent 2100对文库的浓度和插入片段大小(Insert Size)进行检测,使用Q-PCR方法对文库的有效浓度进行准确定量,以保证文库质量。
1.4 测序和数据处理
利用Illumina HiSeqTM 2500平台进行高通量测序,测序读长为PE125。通过对原始序列(Raw reads)去除接头、重复冗余、低质量序列来获得高质量序列(Clean reads),并采用TopHat2[14]软件将clean reads与桃参考基因组(ftp://ftp.bioinfo.wsu.edu/species/Prunus_persica/Prunus_persica-genome. v1.0/)进行序列比对(允许有2个碱基的错配)。
1.5 差异基因的筛选
采用FPKM(Fragments Per Kilobase of transcript per Million fragments mapped)值来反映基因的表达量[15],然后将不同样品的基因表达量使用IDEG6软件(http://telethon.bio.unipd.it/ bioinfo/IDEG6)进行卡方检验,通过多重假设检验(false discovery ratio,FDR)对P值进行校正;校正后取P<1%,并且样品间FPKM比值(Fold change≥2)≥2的基因,作为差异表达基因。1.6 差异表达基因的COG分类、GO分类和KEGG富集分析
将差异表达基因和Nr数据库进行比对,获得差异表达基因在Nr数据库中的注释信息;与COG(Clusters of orthologous groups of proteins)数据库进行比对分析,获得差异表达基因的COG功能注释及其分类;利用Blast2GO[16]和WEGO软件[17]对差异表达基因进行GO(Gene ongology)功能注释及分类;与KEGG(Kyoto encyclopedia of genes and genomes)数据库进行比对,获得差异表达基因相对应的Pathway注释信息。1.7 差异表达基因荧光定量PCR分析
采用TransScript II All-in-One First-Strand cDNA Synthesis SuperMix for qPCR(北京全式金)试剂盒对1.2中提取的RNA进行反转录;用SYBRGreen染料进行荧光定量PCR(qRT-PCR)分析,以2-ΔΔCt法计算基因的差异倍数。每个样品3次重复。选用PpN1(ppa009483m)为内参基因,引物序列参考文献[18](表1)。Table 1
表1
表1用于qRT-PCR分析的基因及其引物
Table 1The qRT-PCR primers for the 6 selected genes
| 基因名称 Gene name | 引物序列 Primer sequence (5′-3′) | 登录号 Accession No. (Phytozome/ GenBank) |
|---|---|---|
| CHI | F:TGAAGACCTCAAGGAACTTCTCAATGG | ppa011276m |
| R:ACACAGGTGACAACGATACTGCCACT | ||
| F3H | F:TCCGAGGGCAGAGCGAAGAAC | ppa007636m |
| R:TTGTGGAGGCTTGTGAGGATTGG | ||
| DFR | F:GGTCGTCCAGGTGAACATACTGCC | ppa008069m |
| R:ATTTCTCATGCCATCCATGCCAC | ||
| LDOX | F:AAGTGGGTCACTGCCAAGTGTGTTC | ppa007738m |
| R:GTGGCTCACAGAAAACTGCCCAT | ||
| ANR | F:ACTTCAAGGCTAAGGGGCTGCTG | ppa008295m |
| R:CCAAGCCAGATAAACGCCAATCAC | ||
| PpN1 | F:CCAGGAGAATCGGTGAGCAGAAAA | ppa009483m |
| R:TCGAGGGTGGAGGACTTGAGAATG |
新窗口打开
1.8 果实品质检测
选用1.1部分采收的果实,每个处理随机选取30个果实测定平均单果质量,根据GB/T12295-1990、GB/T12456-2008和GB/T6194-1986标准委托农业部食品质量监督检验测试中心(济南)检测果实可溶性固形物、可溶性总糖和总酸含量的测定。2 结果
2.1 测序数据质量分析
遮光性套袋和对照的测序原始数据碱基组成基本平衡,而且大部分reads的碱基质量值分布于20以上,说明测序所得的原始数据质量较好,可以满足后续的分析。2个样品共得到16.62 Gb Clean Data测序数据,并且碱基百分比(Q30)大于91%。充分说明测序得到的数据质量可靠,满足后续分析。遮光性套袋和对照分别获得65 300 730个reads和66 603 686个reads,与参考基因组匹配的reads分别有55 984 355个和56 344 011个,占总读数的85.73%和84.60%。表明2个样品的reads与参考基因组的比对效率较高。
2.2 差异表达基因的筛选
通过对桃果实转录组FPKM值的测定(表2),发现遮光性套袋处理的桃果实转录组FPKM值<5的基因占70.50%,>100的基因占4.25%;对照组的FPKM值<5的基因占70.59%,>100的基因占4.34%,二者差异不显著。为了检测遮光性套袋对桃果实基因表达的影响,使用IDEG6软件,将差异倍数(fold change)>2,显著水平P<0.05作为显著差异的表达基因。共获得1 963个差异表达基因,其中,下调基因708个,上调基因1 255个,进一步将这些差异基因在Nr数据库中进行注释,得到1 957个注释基因,其中下调基因有705个,上调基因有1 252个。Table 2
表2
表2桃果实FPKM值
Table 2FPKM values of peach fruits
| FPKM值 FPKM value | 0-1 | 1-5 | 5-10 | 10-100 | >100 | 总计 Total |
|---|---|---|---|---|---|---|
| 套袋 Bagged(QX) | 16995 (56.25%) | 4305 (14.25%) | 2217 (7.34%) | 5410 (17.91%) | 1284 (4.25%) | 30211 |
| 不套袋 Non-bagged (QXCK) | 17167 (56.82%) | 4160 (13.77%) | 2136 (7.07%) | 5447 (18.03%) | 1301 (4.34%) | 30211 |
新窗口打开
2.3 差异表达基因COG注释
对Nr数据库中注释到的1 957个差异表达基因在COG数据库中进行比对和功能注释,结果表明,在COG功能分类体系中,共获得853个COG功能注释,涉及23个COG功能类别(图2)。其中,最大群体比例为R(一般功能基因),有242个基因;其次为K类(转录)、L类(复制、重组和修复)和T类(信号传导机制),分别有116、126和105个基因。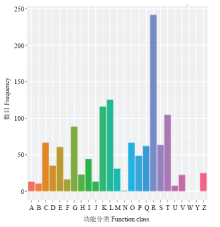 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2差异表达基因COG功能分类图A:RNA加工与修饰;B:染色质结构与变化;C:能量产生与转化;D:细胞周期调控与分裂,染色体重排;E:氨基酸运输与代谢;F:核苷酸运输与代谢;G:碳水化合物运输与代谢;H:辅酶运输与代谢;I:脂类运输与代谢;J:翻译,核糖体结构与生物合成;K:转录;L:自制、重组与修复;M:胞壁/膜生物发生;N:细胞运动;O:蛋白质翻译后修饰与转运,分子伴侣;P:无机离子运输与代谢;Q:次生产物合成,运输及代谢;R:一般功能基因;S:功能未知;T:信号传导机制;U:胞内分泌与膜泡运输;V:防御机制;W:细胞外结构;Y:核酸结构;Z:细胞骨架
-->Fig. 2COG function classification of differentially expressed genes A: RNA processing and modification; B: Chromatin structure and dynamicsl C: Energy production and conversion; D: Cell cycle control, cell division, chromosome partitioning; E: Amino acid transport and metabolism; F: Nucleotide transport and metabolism; G: Carbohydrate transport and metabolism; H: Coenzyme transport and metabolism; I: Lipid transport and metabolism; J: Translation, ribosomal motility; K: Transcription; L: Replocation, recombination and repair; M: Cell wall/memberance/envelope biogenesis; N: Cell motility; O: Posttranslational modification, protein turnover, chaperones; P: Inorganic ion transport and metabolism; Q: Secondary metabolites biosynthesis, transport and catabolixm; R: General function prediction only; S: Functin unknown; T: Signal transduction mechanisms; U: Intracellular trafficking, secretion, and vesicular transport; V: Defense mechanisms; W: Extracellular structures; Y: Nuclear structure; Z: Cytoskeleton
-->
2.4 差异表达基因的GO的分类
利用Blast2GO软件对差异表达基因进行GO分类和功能注释。结果表明,共分为3大类别(细胞组分、分子功能和生物过程),53个小类(图3),1 963个差异表达基因中有1 609个获得功能注释,其中有1 431个差异基因(占总差异基因的88.94%)分类到细胞组分中的16个类别,另外按照分子功能分成17个类别,有1 342个差异基因(占总差异基因的83.41%),按照生物过程分成20个类别,有1 533个差异基因(占总差异基因的95.28%)。在细胞组分分类中,细胞部分(cell part)和细胞(cell)所占比例最多,分别占97.83%和96.09%,其次是细胞器(organelle),占82.50%;而细胞外基质(extracellular matrix)和细胞外基质部分(extracellular matrix part)比例最低,分别仅有2条序列。在分子功能分类中,结合(binding)和催化活性(catalytic activity)所占比例最多,分别占68.18%和68.48%,其次是转动活性(transporter activity)和核酸结合转录因子活性(nucleic acid binding transcription factor activity),分别占9.24%和8.42%;而营养库活性(nutrient reservoir activity)的比例最低,仅有4条序列。在生物学过程中,细胞过程(cellular process)、代谢过程(metabolic process)、单一生物过程(single organism process)和刺激反应(response to stimulus)所占比例最多,分别占91.45%、88.39%、87.21%和74.56%,而细胞杀伤(cell killing)和生物相(biological phase)比例最低,仅有2条和1条序列。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3差异表达基因GO功能分类图C1:细胞部分;C2:细胞;C3:细胞器;C4:膜;C5:细胞器部分;C6:膜部分;C7:细胞连接;C8:高分子复合物;C9:胞外区;C10:膜包围内腔;C11:细胞外区域部分;C12:细胞外基质;C13:细胞核;C14:细胞外基质部分;C15:病毒体;C16:病毒体部分;M1:结合;M2:催化活性;M3:运输活性;M4:核酸结合转录因子活性;M5:分子传感器活性;M6:结构分子活性;M7:受体活性;M8:酶调节活性;M9:电子载体活性;M10:抗氧化活性;M11:蛋白结合转录因子活性;M12:营养库活性;M13:鸟苷酸交换因子活性;M14:转录调节因子活性;M15:成形素活性;M16:蛋白鞭基因;M17:通道调节活性;B1:细胞过程;B2:代谢过程;B3单一生物体过程;B4:刺激反应;B5:生物调节;B6:发育过程;B7:组织或生物起源细胞组件;B8:多细胞生物过程;B9:定位;B10 :繁殖过程;B11:多生物体过程;B12:信号;B13:生长;B14:免疫系统过程; B15:繁殖;B16:有节奏的过程;B17:生物粘附;B18:移动;B19:生物相;B20:细胞杀伤
-->Fig. 3GO function classification of differentially expressed genes C1: Cell part; C2: Cell; C3: Organelle; C4: Membrane; C5: Organelle part; C6 Membrane part; :C7: Cell junction; C8: Macromolecular complex; C9: Extracellular region; C10: Membrane-enclosed lumen; C11: Extracellular region part; C12: Extracellular matrix; C13: Nucleoid; C14: Extracellular matrix part; C15: Virion; C16 Virion part; :M1: Binding; M2: Catalytic activity; M3: Transporter activity; M4: Nucleic acid binding transcription factor activity; M5: Molecular transducer activity; M6: Structural molecule activity; M7: Receptor activity; M8: Enzyme regulator activity; M9: Electron carrier activity; M10: Antioxidant activity; M11: Protein binding transcription factor activity; M12: Nutrient reservoir activity; M13: Guanyl-nucleotide exchange factor activity; M14: Translation regulator activity; M15:Metallochaperone activity; M16: Protein tag; M17: Channel regulator activity; B1: Cellular process; B2: Metabolic process; B3: single-organism process; B4: Response to stimulus; B5: Biological regulation; B6: Developmental process; B7: Cellular component organization or biogenesis; B8: Multicellular organismal process; B9: Localization; B10: Reproductive process; B11: Multi-organism process; B12: Signaling; B13: growth; B14: Immune system process; B15: Reproduction; B16: Rhythmic process; B17: Biological adhesion; B18: Locomotion; B19: Biological phase; B20: Cell kill
-->
2.5 差异表达基因KEGG富集分析
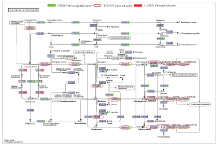
差异表达基因的KEGG分析结果如图4所示,共有421个差异表达基因被注释到了94个通路中,其中显著富集通路(P<0.05)有5个,主要参与到光合作用通路、类黄酮生物合成、核糖体生物合成等通路中(表3)。在各通路中注释到基因多数表达下调,其中光合作用(photosynthesis)注释到25个差异表达基因(22个下调,3个上调),光合作用-天线蛋白通路 (photosynthesis-antenna proteins)注释到9个差异表达基因(8个下调,1个上调),类黄酮生物合成途径(flavonoid biosynthesis),注释到12个差异表达基因(11个下调,1个上调),DNA复制路径(DNA replication)注释到13个差异表达基因(11个下调,2个上调),光合生物碳固定作用通路(carbon fixation in photosynthetic organisms)注释到14个差异表达基因(13个下调,1个上调)。类黄酮生物合成途径KEGG数据库图谱如图5所示,差异表达的基因大多下调(绿色),注释到的12个差异表达基因多数参与花色苷生物合成途径,如F3H(ppa007636m)、LDOX(ppa007738m)、DFR(ppa008069m)、ANR(ppa008295m)、CHI(ppa011276m)等功能基因,且均表现下调。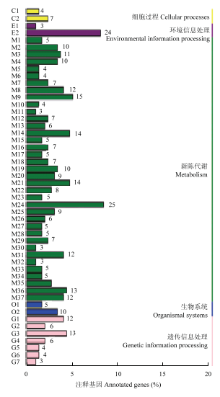 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4差异表达基因KEGG功能分类图C1:内噬作用;C2:吞噬体;E1:ABC转动蛋白;E2:植物激素信号转导;M1:丙氨酸,天门冬氨酸和谷氨酸代谢;M2:丙氨酸和脯氨酸代谢;M3:半胱氨酸和蛋氨酸代谢;M4:苯丙氨酸代谢;M5:色氨酸代谢;M6:酪氨酸代谢;M7:缬氨酸、亮氨酸和异亮氨酸降解;M8:类黄酮生物合成;M9:苯丙素的生物合成;M10:芪类化合物及姜辣素生物合成;M11:氨基糖和核苷酸糖代谢;M12: 维生素C代谢;M13:果糖和甘露糖代谢;M14:糖酵解途径;M15:二羧酸代谢;M16:戊糖、葡萄糖醛酸转换;M17:磷酸戊糖途径;M18:丙酸代谢;M19:丙酮酸盐代谢;M20:淀粉与蔗糖的代谢;M21:光合生物固碳;M22:氮素代谢;M23:氧化磷酸化;M24:光合作用;M25:光合作用-天线蛋白;M26:硫代谢;M27:脂肪酸代谢;M28:不饱和脂肪酸生物合成;M29:卟啉与叶绿素代谢;M30:氰基氨基酸代谢;M31:谷胱甘肽代谢;M32:含硒化合物代谢;M33:β-丙氨酸代谢;M34:类胡萝卜素生物合成;M35:萜类化合物骨架生物合成;M36:嘌呤代谢;M37:嘧啶代谢作用;O1:植物昼夜节律;O2:植物病原菌相互作用;G1:内质网蛋白加工;G2:水解作用;G3:DNA自制;G4:同源重组;G5:错配修复;G6:剪接体;G7:氨酰tRNA生物合成
-->Fig. 4KEGG function classification of differentially expressed genes C1: Endocytosis; C2: Phagosome; E1: ABC transporters; E2: Plant hormone singnal transduction; M1: Alanine, aspartate and glutamate metabolism; M2: Arginine and proline metabolism; M3: Cysteine and methionine metabolism; M4: Phenylalanine metabolism; M5: Tryptophan metabolism; M6: Tyrosine metabolism; M7: Valine, leucine and isoleucine degradation; M8: Flavonoid biosynthesis; M9: Phenylpropanoid biosynthesis; M10: Stilbenoid, diarylheptanoid and gingerol biosynthesis; M11: Amino sugar and nucleotide sugar metabolism; M12:Ascorbate and aldarate metabolism; M13: Fructise and mannose metabolism; M14: Glycolysis/Gluconeogenesis; M15: Glyoxylate and dicarboxylate metabolism; M16: Pentose and glucuronate interconversions; M17: Pentose and glucuronate interconversions; M18: Propanoated metabolism; M19: Pyruvate metabolism; M20: Starch and sucrose metabolism; M21: Carbon fixation in photosynthetic organisms; M22: Nitrogen metabolism; M23: Oxiative phosphorylation; M24: Photosynthesis; M25: Photosynthesis-antenna proteins; M26: Sulfur metabolism; M27: Fatty acid metabolism; M28: Biosynthesis of unsaturated fatty acids; M29: Porphyrin and chlorophyll metabolism; M30: Cyanoamino acid metabolism; M31: Glutathiome metabolism; M32: Selenocompoumd metabolism; M33: Beta-Alanine metabolism; M34: Carotenoid biosynthesis; M35: Terpenoid backbone biosynthesis; M36: Purine metabolism; M37: Pyrimidime metabolism; O1: Circadian rhythm-plant; O2: Plant-pathogen interaction; G1: Protein processing in endiplasmic reticulum; G2: Ubiquitin mediated proteolysis; G3: DNA replication; G4: Homologous recombination; G5: Mismathc repair; G6: Spliceosome; G7: Aminoacyl- tRNA biosynthesis
-->
Table 3
表3
表3差异表达基因KEGG代谢途径分类
Table 3KEGG classification of differentially expressed genes
| 序号 No. | 代谢通路 Metabolism_pathway | DGE数量 DEG number | 通路ID Pathway ID | P值 P value |
|---|---|---|---|---|
| 1 | 光合作用 Photosynthesis | 25 | ko00195 | 1.77E-17 |
| 2 | 光合作用-天线蛋白 Photosynthesis - antenna proteins | 9 | ko00196 | 0.000606704 |
| 3 | 类黄酮生物合成途径 Flavonoid biosynthesis | 12 | ko00941 | 0.001013209 |
| 4 | DNA复制 DNA replication | 13 | ko03030 | 0.001148887 |
| 5 | 光合生物碳固定作用 Carbon fixation in photosynthetic organisms | 14 | ko00710 | 0.032804394 |
新窗口打开
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5KEGG数据库中类黄酮生物合成途径
-->Fig.5Flavonoid biosynthesis pathway in KEGG
-->
2.6 差异表达基因的qRT-PCR荧光定量分析
对类黄酮途径中的关键基因F3H(ppa007636m)、LDOX(ppa007738m)、DFR(ppa008069m)、ANR(ppa008295m)、CHI(ppa011276m)进行qRT-PCR荧光定量分析(表4),发现遮光性套袋处理的桃果实样品各基因的表达均与对照表现下调趋势,只是差异倍数上与转录组的数据有所区别。Table 4
表4
表4差异表达基因的RNA-Seq FPKM值、qRT-PCR相对表达水平及其比率
Table 4RNA-Seq FPKM value, qRT-PCR relative expression level, ratio of CK and QX in the differentially expressed genes
| 方法 Method | 样品 Sample | CHI | F3H | DRF | LDOX | ANR |
|---|---|---|---|---|---|---|
| RNA-Seq | CK | 331.9 | 810.1 | 966.2 | 5122.7 | 80.0 |
| QX | 59.8 | 65.7 | 13.9 | 237.2 | 24.5 | |
| CK/QX | 5.55 | 12.33 | 69.51 | 21.60 | 3.27 | |
| qRT-PCR | CK | 1.00 | 1.04 | 0.62 | 0.28 | 0.26 |
| QX | 0.06 | 0.41 | 0.01 | 0.08 | 0.07 | |
| CK/QX | 16.67 | 2.54 | 62.00 | 4.75 | 3.71 |
新窗口打开
2.7 果实品质的检测
由表5的测评结果可知桃果实遮光性套袋对果实的可溶性固形物及可溶性总糖产生显著性影响,可溶性固形物及可溶性总糖显著降低,而对于果实总酸的影响不大。在果实大小上几乎没有影响。Table 5
表5
表5果实品质检测分析
Table 5Analysis of the fruit quality detection
| 样品 Sample | 可溶性固形物 Soluble solids (%) | 可溶性总糖 Total soluble sugar (%) | 总酸 Total acid (%) | 单果重 Per fruit weight (g) |
|---|---|---|---|---|
| QX | 10.1b | 7.2b | 0.21a | 233.9a |
| CK | 12.4a | 9.9a | 0.23a | 242.5a |
新窗口打开
3 讨论
高通量转录组测序技术在挖掘差异表达基因,揭示生物基因表达及调控机理发挥了重要作用 [19-22]。本研究通过Illumina HiSeqTM 2500高通量测序技术构建了遮光性套袋桃果实及其对照的转录组文库,获得大量的转录组信息。处理与对照共获得16.62 Gb Clean Data 数据量,比对到桃参考基因组上的reads在84%以上,说明本试验文库构建很成功,数据覆盖度较高。通过对文库分析,获得1 963个差异基因,其中1 957个基因在Nr数据库中得到功能注释,未得到注释的差异表达基因有6个;有1 609个基因获得GO功能注释,348个基因未得到GO功能注释;421个基因被注释到了KEGG的94个代谢通路中,153个基因未得到KEGG的注释。这种造成基因注释信息缺失的现象,前人认为转录组序列越短,获得注释信息就越少[23]。COG功能分类表明,853个差异表达基因获得COG功能注释,包括23个功能类别。GO功能注释分类显示,差异表达基因主要涉及细胞相关类别(包括细胞部分、细胞、细胞器等)、分子结合、细胞过程、代谢过程等类型。深入对差异基因进行Pathway分析,进一步对差异表达基因参与的主要代谢途径和信号传导途径进行研究,有利于明确差异表达基因在生物代谢周期中发辉的功能以及与其他基因的相互作用[22-23]。本研究KEGG Pathway富集分析表明,差异表达基因共涉及到94个通路,其中光合作用相关通路3个、类黄酮生物合成1个、核糖体生物合成1个等5个通路显著富集。果实色泽是果实商品品质的重要因素之一,果实成熟着色是由于叶绿素降解,同时形成显现类胡萝卜素或花青苷的结果。花青苷是一类类黄酮化合物,花青苷是果实的次生代谢产物之一,它是经过莽草酸途径合成的[24],花青苷的生物合成是依赖光照进行的反应过程[25]。花青苷是由花色素和糖组成,而花色素又是在糖代谢的基础上由丙酮酸和乙酸缩合而成的[26-27]。花青苷含量和着色程度与果肉中还原糖和可溶性糖含量呈显著相关,可溶性固形物含量与花青苷合成呈正相关[28-29]。套袋虽然在一定程度上降低果实的含糖量,但也能有效地促进果实着色[13,30]。这一事实说明果实着色在一定范围内受限于果实中含糖量,如果越过一定阈值则含糖量不再成为限制因素;再则充分表明光照对果实着色的重要性,果实套袋增加光有效性的同时也促使光照不足,以致有关基因不能活化[31-32]。前人研究认为光照影响花青苷合成和果实着色,首先光照影响光合作用,进而影响糖、苯丙氨酸等有机物合成;其次光照调解花青苷合成相关酶的活性,其中查尔酮合成酶(CHS)、查尔酮异构酶(CHI)、黄烷酮3-羟化酶(F3H)、二羟基黄酮醇还原酶(DFR)、花色素合成酶(ANS)等基因的表达受到光照因子的诱导[33-37]。本研究显著富集的通路中有3个与光合作用相关,充分说明了光照在果实成熟期对着色的重要性,同时又显著富集到类黄酮生物合成通路,进一步说明光照调控花青苷代谢过程相关基因表达的事实。本研究筛选到了参与此通路上的F3H、LDOX、DFR、ANR、CHI等5个差异表达基因,且表达水平较对照低,说明套袋增加了光的有效性,但光照不足,以致有关基因不能活化或表达水平较低,此结果与王惠聪在荔枝[27]上和马瑞娟等[38]在桃上的研究相似。同时检测发现遮光性套袋显著降低了果实的可溶性固形物的含量,进一步证明套袋在一定程度上降低果实的含糖量的论点,说明可溶性固形物含量与花青苷合成呈正相关。此外,核酸生物合成(DNA复制)也得到了显著富集,这类代谢通路在果实着色上的作用尚不明确。差异表达基因KEGG Pathway分析表明光照与类黄酮生物合成通路调控着果实着色,但具体的调控机制还需要对相关通路的基因进行更深入的研究。
4 结论
通过高通量测序技术获得一定数量在遮光性套袋处理状态下桃果实差异表达基因,光合作用通路和类黄酮生物合成通路基因在果实着色中发挥了重要作用,再次印证了遮光性套袋对桃果实的可溶性固形物及可溶性总糖产生显著性影响。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}