0 引言
【研究意义】谷子抗旱耐瘠、适应性广、营养丰富,在中国北方旱作农业中占有重要地位[1-4]。谷子的幼穗分化标志着营养生长向生殖生长的转变,是个体生长发育的重要阶段,具有自身的规律性。叶片是谷子的主要同化器官,其同化能力与叶龄指数紧密关联[5-6]。研究谷子穗分化和叶龄指数的关系,是形态诊断的核心内容,在栽培指标化管理中发挥着重要作用[7]。【前人研究进展】早在1940年EVANS等[8]描述了谷子生长初期的花序发育;朱澂等[9]1957年对春谷农家品种的穗分化进行了详细观察,奠定了谷子穗分化研究的基础;1986年,辛淑芳等[10]通过研究谷子外部形态与幼穗分化各个时期的关系时发现,以叶龄指数作为判断穗分化时期的标准比较理想。1998年,段胜军等[11]通过扫描电镜观察,将谷子穗分化过程划分为5个时期,并就其与栽培育种的关系等问题进行了讨论;2012年,卢海博等[12]以杂交谷为研究对象,指出可以利用回归方程,根据叶龄指数来预测穗分化时期。【本研究切入点】近年来,国内外对幼穗分化及其与外部形态器官的相关性研究主要集中在小麦、水稻、玉米等主要禾谷类作物[13-19],谷子相关研究较为迟缓且报道甚少[20-25]。谷子品种类型繁多,不同叶数的品种在同一叶龄的生育进程及各部器官生长状态不同,播期等栽培管理措施对其影响也较大,而目前的相关研究较少,同时,前人描述谷子幼穗分化过程的观察手绘草图或实物照片受当时试验条件的限制,不够清晰系统,因此,亟需深入研究不同品种谷子幼穗分化进程的差异以及幼穗分化与叶龄指数的关系,探究播期对其的调控效应,从而明确不同品种谷子在不同播期条件下生育进程的简易叶龄诊断方法,并完善谷子幼穗分化进程图。【拟解决的关键问题】本研究通过观察不同品种谷子在不同播期条件下的幼穗分化详细过程,同时记录不同生育期的叶龄,揭示谷子穗分化与叶龄指数的对应关系,旨在为谷子生产中的科学管理提供依据。1 材料与方法
1.1 供试材料
济谷16、济谷18分别为抗拿捕净除草剂和糯性谷子新品种,由山东省农业科学院作物研究所提供。试验于2015年在山东省农业科学院作物研究所济南试验基地进行,土壤质地为壤土,0—20 cm耕层土壤含有机质16.9 g·kg-1、速效氮72.4 mg·kg-1、速效磷18.8 mg·kg-1、速效钾168 mg·kg-1,pH为7.8,每公顷基施氮磷钾(15-15-15)三元素复合肥375 kg,腐熟有机肥750 kg。试验地前茬为冬小麦,栽培管理措施同丰产田。试验从4月29日开始播种,之后每10天播种1期直至7月8日,共播种8期,依次编号为“1—8”。小区行长5 m,6行区,行距0.5 m,密度为60万株/hm2,依品种按播期顺序排列。
1.2 测定方法
在谷子5叶期,每个小区选择120株生长均匀一致的植株进行标记,每隔1—2 d选取有代表性的植株3—5株在解剖镜下观察幼穗分化结果,并详细记载幼穗发育的阶段及其对应的叶龄。幼穗分化过程观察:在OLYMPUS SZX16体视显微镜下,剥去样品苞叶至露出完整幼穗,进行观察并拍照。
幼穗分化时期划分:营养生长期(穗分化前期)、生长锥伸长期(Ⅱ)、枝梗分化期(Ⅲ-1、Ⅲ-2、Ⅲ-3)、小穗和刚毛分化期(Ⅳ)以及雌雄蕊分化期(小花分化期,Ⅴ)共5个时期[11]。
叶龄(leaf number,LN):展开叶片数,以叶片与叶鞘交界处的叶枕露出为准[26],半展开叶按照比例值估算。
总叶片数(total leaf number,TLN):按田间标记叶龄的20个植株主茎总叶片数的平均值计算。
叶龄指数(leaf number index,LNI):已出叶片数占主茎总叶片数的百分比,即LNI(%)=
1.3 数据分析
采用Microsoft Excel 2007进行数据处理,SPSS 20.0数据处理软件进行方差分析、相关分析、回归分析及配合力分析,Origin 8.0绘图软件进行图形分析及绘制[27]。2 结果
2.1 谷子幼穗分化过程
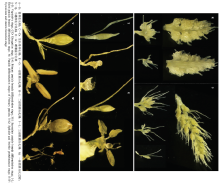
谷穗的分化形成,是一个连续分化发育的过程,根据各阶段生长和分化的形态特征差异,可以将其划分为5个分化时期,而不同品种谷子在不同播期条件下幼穗分化的方式及其形态特征基本一致,因此,用一组济谷16的幼穗发育图片即可描绘谷子幼穗分化的整个过程(图1)。2.1.1 营养生长期 幼苗出土至拔节阶段为营养生长期,生长锥呈光滑的半球体(图1-A),并未开始伸长,主要进行叶、节和节间的分化。生长点随着叶龄的增加逐渐增大,基部一般可见2个明显的半环状互生突起,即为叶原基(图1-B)。叶原基继续生长包围生长锥,继而形成叶片和叶鞘。谷子营养生长期的持续时间随着播期的推迟逐渐缩短。
2.1.2 生长锥伸长期 茎的节和节间进行居间生长,标志着拔节的开始;叶原基分化停止,生长锥开始伸长(图1-C和图1-D),标志着幼穗分化阶段的正式开始,营养生长逐渐转变为生殖生长。较早播种的谷子拔节期比生长锥伸长期开始的早,随着播期的推迟,其间隔时间逐渐缩短直至基本同步,甚至会有生长锥伸长期早于拔节期的情况出现。
2.1.3 枝梗分化期 生长锥的顶端保持光滑并继续延伸形成谷穗中轴,在其基部出现小的突起,即为一级枝梗原基(图1-E),呈向顶式生长,各枝梗原基之间略有凹陷。当每列的一级枝梗原基数达十多个后,生长锥不再伸长,而由于中轴上部一级枝梗的发育时间较晚且速度较慢,故顶部的谷码较小(图1-F)。在幼穗分化的早中期,一级枝梗原基表现为排列整齐的纵列,每个中轴上有6—8列(图1-I),之后随着生长发育的进程逐渐变为旋转式分布(图1-G)。中轴伸长的长短和一级枝梗原基的多少决定了谷穗的谷码数,进而决定了谷穗的大小。
当一级枝梗的发育达到一定体积后,在其基部出现左右互生的突起,即为二级枝梗原基(图1-H)。二级枝梗也呈向顶式分化,从而使得一级枝梗在整体上表现为三角形状;三级枝梗原基在二级枝梗上以同样的方式发生(图1-J)。谷穗中下部的一级枝梗分化出的二、三级枝梗数量多,而上部的一级枝梗的分枝数明显较少(图1-K),在很大程度上决定了谷码的大小,进而影响结实粒数。
2.1.4 小穗和刚毛分化期 三级枝梗分化完成后(图1-L和图1-M),在其上产生众多乳头状突起,即为小穗或刚毛原基(图1-N),顶端多数是光滑的,少部分表现为钝形的凹陷。刚毛原基和小穗原基的初期形态基本一致,难以区分。小穗原基基部继续膨大,顶端钝圆光滑,基部出现同叶原基分化形式相似的外颖片原基和内颖片原基,而刚毛原基不发生增粗变化,只进行伸长,且速度快于小穗,顶端变成梯形或其他不规则形状(图1-O)。当小穗上的小花分化进行到雌雄蕊原基形成时,刚毛上产生许多细小突起,谷穗成熟后这些突起形成刚毛上的刺(图1-P和图1-Q)。穗轴和枝梗上的毛刺也是以这种方式形成的。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1-1济谷16的幼穗分化过程
-->Fig. 1The panicle differentiation stages of Jigu16
-->
2.1.5 雌雄蕊分化期 内外护颖原基形成,标志着小花分化的开始(图1-R和图1-S)。谷子每个小穗有2朵小花,第1朵花为败育花,第2朵花才是正常发育结实的小花。第1朵花的外稃相对于小穗内颖而生,内稃位于外稃对面,其外稃的上方有偏圆形突起,即为第1朵花原基。第1朵花至此基本上完成,不再进行雌雄蕊等其他器官的分化,在以后发育过程中逐渐消失。第2朵花原基位于第1朵花之上,是小穗原基的原生长点转变形成。可育花的外稃相对于第1朵花的生长点而生,其最初形状为偏平半球形突起,但分化后生长较快,内稃位于外稃之上的相对侧,分化后生长相对较慢。可育花的内外稃分化完成之后,在其生长点顶部的侧面出现3个顶部较平坦的雄蕊原基(图1-T),1个位于外稃的正上方,另2个在原基侧面呈相对状(图1-V)。花药原基形成后,可育花原基的顶端部分由平坦转变为突起,即为雌蕊原基。雌蕊分化最早的为子房,形状新月形,然后在其左右两侧形成2个尖角突起,以后形成花柱和柱头(图1-U)。小花分化完成之后,各部分器官进行增大生长和发育,由于护颖的生长,开花前一直将雌雄蕊等器官包被在其内。开花前雌蕊发育成具有子房、花柱和羽毛状二歧柱头的完整器官,花柱侧生子房内侧(图1-W)。雄蕊原基增粗增大,形成具有花粉囊的花药。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1-2 济谷16的幼穗分化过程
-->Fig. 2The panicle differentiation stages of Jigu16
-->
2.2 不同播期夏谷的幼穗发育进程
随着播期的推迟,幼穗分化开始日期逐渐提前(表1),济谷16由出苗后32 d开始穗分化缩短到23 d,变异系数为11.03%,济谷18由31 d缩短到22 d,变异系数为10.86%;幼穗分化持续时间逐渐缩短,济谷16由28 d减少到22 d,变异系数为8.92%,济谷18由28 d减少到19 d,变异系数为13.18%;从抽穗到成熟阶段的持续天数逐渐缩短,济谷16由49 d变为41 d,变异系数为5.39%,济谷18由49 d变为42 d,变异系数分别为4.96%;从出苗到成熟阶段的生育期持续天数逐渐缩短,济谷16由109 d变为86 d,变异系数为7.77%,济谷18由108 d变为83 d,变异系数分别为8.57%。通过比较变异系数可以发现,随着播期的推迟,谷子幼穗分化阶段开始的时间及持续天数的变化幅度较大,而抽穗到成熟阶段持续天数的变化幅度较小,说明播期对谷子生育期的影响主要是由于幼穗分化阶段的开始时间和持续天数的变化引起的。Table 1
表1
表1不同播期条件下夏谷各幼穗分化时期的开始日期及持续天数
Table 1The occurred date and continued days of each panicle differentiation stage in different sowing date
| 品种 Cultivar | 处理 Treatment | 出苗期 Seeding stage | 拔节期 Jointing stage | Ⅱ期 | Ⅲ期 | Ⅳ期 | Ⅴ期 | 抽穗期 Heading stage | 成熟期 Mature stage | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ⅲ-1 | Ⅲ-2 | Ⅲ-3 | |||||||||||||||||||||||
| 日期 Date | 天数 Days(d) | 日期 Date | 天数 Days(d) | 日期 Date | 天数 Days(d) | 日期 Date | 日期 Date | 日期 Date | 天数 Days(d) | 日期 Date | 天数 Days(d) | 日期 Date | 天数 Days(d) | 日期 Date | 天数 Days(d) | 日期 Date | 天数 Days(d) | ||||||||
| 济谷16 Jigu16 | 1 | 5.8 | 25 | 6.2 | 7 | 6.9 | 5 | 6.14 | 6.18 | 6.22 | 12 | 6.26 | 5 | 7.1 | 6 | 7.7 | 49 | 8.25 | 109 | ||||||
| 2 | 5.17 | 24 | 6.10 | 6 | 6.16 | 5 | 6.21 | 6.25 | 6.29 | 12 | 7.3 | 5 | 7.8 | 6 | 7.14 | 47 | 8.30 | 105 | |||||||
| 3 | 5.26 | 23 | 6.18 | 6 | 6.24 | 5 | 6.29 | 7.3 | 7.7 | 12 | 7.11 | 5 | 7.16 | 5 | 7.21 | 46 | 9.5 | 102 | |||||||
| 4 | 6.4 | 22 | 6.26 | 6 | 7.2 | 4 | 7.6 | 7.10 | 7.14 | 12 | 7.18 | 5 | 7.23 | 5 | 7.28 | 46 | 9.12 | 100 | |||||||
| 5 | 6.13 | 22 | 7.5 | 5 | 7.10 | 4 | 7.14 | 7.17 | 7.21 | 11 | 7.25 | 5 | 7.30 | 5 | 8.4 | 46 | 9.19 | 98 | |||||||
| 6 | 6.22 | 21 | 7.13 | 5 | 7.18 | 4 | 7.22 | 7.25 | 7.29 | 11 | 8.2 | 4 | 8.6 | 5 | 8.11 | 45 | 9.25 | 95 | |||||||
| 7 | 7.2 | 20 | 7.22 | 4 | 7.26 | 4 | 7.30 | 8.2 | 8.5 | 10 | 8.9 | 4 | 8.13 | 5 | 8.18 | 43 | 9.30 | 90 | |||||||
| 8 | 7.11 | 19 | 7.30 | 4 | 8.3 | 3 | 8.6 | 8.9 | 8.12 | 10 | 8.16 | 4 | 8.20 | 5 | 8.25 | 41 | 10.5 | 86 | |||||||
| CV | 9.09 | 19.73 | 16.64 | 7.88 | 11.19 | 8.82 | 5.39 | 7.77 | |||||||||||||||||
| 济谷18 Jigu18 | 1 | 5.9 | 25 | 6.3 | 6 | 6.9 | 5 | 6.14 | 6.18 | 6.22 | 13 | 6.27 | 5 | 7.2 | 5 | 7.7 | 49 | 8.25 | 108 | ||||||
| 2 | 5.18 | 24 | 6.11 | 5 | 6.16 | 5 | 6.21 | 6.25 | 6.29 | 13 | 7.4 | 5 | 7.9 | 5 | 7.14 | 47 | 8.30 | 104 | |||||||
| 3 | 5.27 | 23 | 6.19 | 5 | 6.24 | 4 | 6.28 | 7.2 | 7.6 | 12 | 7.10 | 5 | 7.15 | 5 | 7.20 | 47 | 9.5 | 101 | |||||||
| 4 | 6.5 | 23 | 6.28 | 4 | 7.2 | 4 | 7.6 | 7.9 | 7.13 | 11 | 7.17 | 5 | 7.22 | 5 | 7.27 | 46 | 9.11 | 98 | |||||||
| 5 | 6.13 | 23 | 7.6 | 3 | 7.9 | 4 | 7.13 | 7.16 | 7.19 | 10 | 7.23 | 5 | 7.28 | 5 | 8.2 | 46 | 9.17 | 96 | |||||||
| 6 | 6.22 | 22 | 7.14 | 3 | 7.17 | 3 | 7.20 | 7.23 | 7.26 | 10 | 7.30 | 5 | 8.4 | 5 | 8.9 | 45 | 9.23 | 93 | |||||||
| 7 | 7.1 | 22 | 7.23 | 2 | 7.25 | 3 | 7.28 | 7.31 | 8.3 | 9 | 8.6 | 4 | 8.10 | 5 | 8.15 | 43 | 9.27 | 88 | |||||||
| 8 | 7.11 | 21 | 8.1 | 1 | 8.2 | 3 | 8.5 | 8.7 | 8.10 | 8 | 8.13 | 4 | 8.17 | 4 | 8.21 | 42 | 10.2 | 83 | |||||||
| CV | 5.45 | 46.48 | 21.54 | 17.04 | 9.75 | 7.25 | 4.96 | 8.57 | |||||||||||||||||
新窗口打开
谷子幼穗各分化时期的持续时间也随着播期的推迟逐渐缩短,最早播种处理与最晚播种处理的天数相差在2 d左右,其中,以生长锥伸长期(Ⅱ期)的变化幅度最大,济谷16和济谷18的变异系数分别为16.64%和21.54%,其他各时期(Ⅲ期、Ⅳ期、Ⅴ期)的变异系数也均在10%左右,济谷16分别为7.88%、11.19%和8.82%,济谷18分别为17.04%、9.75%和7.25%。拔节期到生长锥伸长期(Ⅱ期)的时间间隔随着播期的推迟明显减少,济谷16由7 d缩短为4 d,变异系数为19.73%,济谷18由6 d缩短为1 d,变异系数为46.48%。
2.3 不同播期夏谷的叶龄及叶龄指数
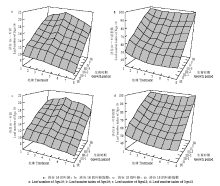
结合表2、图2可以看出,随着播期的推迟,在生长锥伸长期(Ⅱ期),谷子的叶片数逐渐减少,由11.9—12.0片叶变为9.3—9.4片叶,呈“乀”型下降状态;从枝梗分化期到小穗刚毛分化期(Ⅲ-1期到Ⅳ期),谷子叶片数整体变化趋势仍为逐渐降低,但在第4个处理有反超早播和晚播的现象,呈“∽”型降升降状态;雌雄蕊分化期(Ⅴ期)之后,谷子叶片数先增加后减少,呈“∧”型升降状态;到成熟期时,不同播期条件下的谷子总叶片数差异显著,且济谷16和济谷18都是在第4处理最多、第8处理最少,分别为21.8、17.2和21.1、17.3片叶。Table 2
表2
表2不同播期条件下夏谷幼穗各分化时期的叶片数和叶龄指数
Table 2The leaf number and leaf number index of each panicle differentiation stage in different sowing date
| 品种 Cultivar | 处理 Treatment | 拔节期 Jointing stage | Ⅱ期 | Ⅲ-1期 | Ⅲ-2期 | Ⅲ-3期 | Ⅳ期 | Ⅴ期 | 抽穗期 Heading stage | 成熟期 Mature stage | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 叶龄 LN | 叶龄指数 LNI | 叶龄 LN | 叶龄指数 LNI | 叶龄 LN | 叶龄指数 LNI | 叶龄 LN | 叶龄指数 LNI | 叶龄 LN | 叶龄指数 LNI | 叶龄 LN | 叶龄指数 LNI | 叶龄 LN | 叶龄指数 LNI | 叶龄 LN | 叶龄指数 LNI | 总叶片数 TLN | ||
| 济谷16 Jigu 16 | 1 | 9.5 | 49.79 | 12.0 | 62.89 | 12.9 | 67.61 | 14.4 | 75.47 | 15.6 | 81.76 | 16.8 | 88.05 | 17.6 | 92.24 | 18.6 | 97.48 | 19.08 |
| 2 | 9.3 | 45.93 | 11.7 | 57.78 | 12.6 | 62.22 | 13.8 | 68.15 | 15.0 | 74.07 | 16.5 | 81.48 | 17.8 | 87.90 | 19.6 | 96.79 | 20.25 | |
| 3 | 9.2 | 43.81 | 11.5 | 54.76 | 12.5 | 59.52 | 13.2 | 62.86 | 14.6 | 69.52 | 16.5 | 78.57 | 18.0 | 85.71 | 20.0 | 95.24 | 21.00 | |
| 4 | 9.1 | 41.84 | 11.4 | 52.41 | 12.6 | 57.93 | 14.0 | 64.37 | 15.4 | 70.80 | 17.4 | 80.00 | 18.9 | 86.90 | 20.8 | 95.63 | 21.75 | |
| 5 | 8.7 | 41.59 | 10.8 | 51.63 | 11.7 | 55.93 | 12.9 | 61.66 | 14.3 | 68.36 | 16.4 | 78.39 | 17.7 | 84.61 | 20.0 | 95.60 | 20.92 | |
| 6 | 8.4 | 42.36 | 10.3 | 51.94 | 11.4 | 57.49 | 12.5 | 63.04 | 14.1 | 71.10 | 15.8 | 79.68 | 16.8 | 84.72 | 19.1 | 96.32 | 19.83 | |
| 7 | 8.1 | 43.20 | 10.0 | 53.33 | 11.0 | 58.67 | 12.3 | 65.60 | 13.9 | 74.13 | 15.2 | 81.07 | 16.2 | 86.40 | 18.2 | 97.07 | 18.75 | |
| 8 | 7.7 | 44.85 | 9.3 | 54.16 | 10.5 | 61.15 | 11.7 | 68.14 | 13.1 | 76.30 | 14.3 | 83.28 | 15.1 | 87.94 | 17.0 | 99.01 | 17.17 | |
| 均值 A | 8.8 | 44.17 | 10.9 | 54.86 | 11.9 | 60.07 | 13.1 | 66.16 | 14.5 | 73.26 | 16.1 | 81.32 | 17.3 | 87.05 | 19.2 | 96.64 | 19.84 | |
| CV | 7.28 | 6.15 | 8.65 | 6.92 | 7.38 | 6.08 | 7.07 | 6.73 | 5.71 | 5.92 | 6.08 | 3.88 | 6.87 | 2.82 | 6.30 | 1.28 | 7.43 | |
| 济谷 18 Jigu 18 | 1 | 9.5 | 50.67 | 11.9 | 63.47 | 12.8 | 68.27 | 14.3 | 76.27 | 15.5 | 82.67 | 16.4 | 87.47 | 17.4 | 92.80 | 18.2 | 97.07 | 18.75 |
| 2 | 9.4 | 47.40 | 11.6 | 58.50 | 12.6 | 63.54 | 14.0 | 70.60 | 15.2 | 76.65 | 16.5 | 83.21 | 17.6 | 88.75 | 19.2 | 96.82 | 19.83 | |
| 3 | 9.3 | 45.93 | 11.3 | 55.80 | 12.4 | 61.23 | 13.2 | 65.19 | 14.7 | 72.59 | 16.3 | 80.49 | 17.4 | 85.93 | 19.4 | 95.80 | 20.25 | |
| 4 | 9.3 | 44.12 | 11.3 | 53.61 | 12.5 | 59.30 | 14.0 | 66.41 | 15.4 | 73.06 | 17.1 | 81.12 | 18.2 | 86.34 | 20.2 | 95.83 | 21.08 | |
| 5 | 8.8 | 43.63 | 10.7 | 53.05 | 11.7 | 58.01 | 12.8 | 63.46 | 14.4 | 71.39 | 16.2 | 80.32 | 17.1 | 84.78 | 19.2 | 95.19 | 20.17 | |
| 6 | 8.6 | 43.00 | 10.4 | 52.00 | 11.4 | 57.00 | 12.6 | 63.00 | 14.1 | 70.50 | 15.8 | 79.00 | 16.8 | 84.00 | 19.1 | 95.50 | 20.00 | |
| 7 | 8.5 | 44.34 | 10.1 | 52.69 | 11.1 | 57.90 | 12.5 | 65.21 | 14.0 | 73.03 | 15.3 | 79.81 | 16.4 | 85.55 | 18.5 | 96.50 | 19.17 | |
| 8 | 8.0 | 46.16 | 9.4 | 54.24 | 10.6 | 61.17 | 11.8 | 68.09 | 13.2 | 76.17 | 14.4 | 83.09 | 15.3 | 88.29 | 17.1 | 98.67 | 17.33 | |
| 均值A | 8.9 | 45.66 | 10.8 | 55.42 | 11.9 | 60.80 | 13.2 | 67.28 | 14.6 | 74.51 | 16.0 | 81.81 | 17.0 | 87.06 | 18.9 | 96.42 | 19.57 | |
| CV | 5.98 | 5.48 | 7.77 | 6.95 | 6.77 | 6.11 | 6.70 | 6.52 | 5.46 | 5.26 | 5.20 | 3.33 | 5.17 | 3.25 | 4.92 | 1.16 | 5.86 | |
新窗口打开
随着播期的推迟,谷子从出苗到抽穗的生长发育过程中,叶片生长速率逐渐提高,由每天的0.31片叶增加为0.40片叶。其中,出苗到拔节阶段,叶片生长速率并不随着播期的推迟而有较大变化,基本都维持在每天0.40片叶的状态,济谷16和济谷18之间的差异也较小;拔节到抽穗阶段,济谷16的叶片生长速率随着播期的推迟逐渐升高,由每天0.26片叶变为0.37片叶,均比上一个阶段有明显降低,济谷18的叶片生长速率也随着播期的推迟逐渐升高,由每天0.26片叶提高到0.43片叶,与上一个阶段相比,前4个处理明显降低,后4个处理明显提高。
在一级枝梗分化期之前的各阶段,叶龄指数随着播期的推迟先降低后升高,呈“∨”型降升状态,在二级枝梗分化期之后的各阶段,叶龄指数随着播期的推迟先降低后略微升高再降低之后再逐渐升高,呈“W”型降升降升状态,与叶龄的变化趋势相比存在明显差异。可见,播期对谷子成熟期植株总叶片数的影响,造成了幼穗各分化时期内叶龄与叶龄指数变化趋势的不同。通过比较叶龄与叶龄指数在同一幼穗分化时期的变异系数发现,叶龄指数的CV值均较小,表明叶龄指数与幼穗分化阶段在不同播期间保持着更为稳定的对应关系。
济谷16和济谷18叶龄和叶龄指数的变化趋势基本一致,但数值差异显著。通过分析均值可知,进入拔节期时,济谷16约有8.8片叶、LNI为44.17%,济谷18约有8.9片叶、LNI为45.66%;进入生长锥伸长期时,济谷16约有10.9片叶、LNI为54.86%,济谷18约有10.8片叶、LNI为55.42%;进入一级枝梗分化期时,济谷16约有11.9片叶、LNI为60.07%,济谷18约有11.9片叶、LNI为60.80%;进入二级枝梗分化期时,济谷16约有13.1片叶、LNI为66.16%,济谷18约有13.2片叶、LNI为67.28%;进入三级枝梗分化期时,济谷16约有14.5片叶、LNI为73.26%,济谷18约有14.6片叶、LNI为74.51%;进入小穗刚毛分化期时,济谷16约有16.1片叶、LNI为81.32%,济谷18约有16.0片叶、LNI为81.81%;进入雌雄蕊分化期时,济谷16约有17.3片叶、LNI为87.05%,济谷18约有17.0片叶、LNI为87.06%;进入抽穗期时,济谷16约有19.2片叶、LNI为96.64%,济谷18约有18.9片叶、LNI为96.42%。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2叶片数及叶龄指数的三维图形
-->Fig. 23D graphics of leaf number and leaf number index
-->
2.4 不同播期夏谷的幼穗分化与叶龄指数的关联
用自然数1—8分别代表“拔节期、Ⅱ期、Ⅲ-1期、Ⅲ-2期、Ⅲ-3期、Ⅳ期、Ⅴ期、抽穗期”作为统计回归分析的依变量Y,幼穗各分化时期所对应的叶龄指数作为自变量X。通过分析发现,不同品种谷子在不同播期条件下,幼穗分化阶段与叶龄指数均符合直线回归关系,R2值在0.977—0.997(表3),表现为极显著正相关性,幼穗分化期Yi依叶龄指数Xi的直线回归方程通式为:Yi=bXi+a(a、b代表系数)。随着播期的推迟,b值逐渐减小,济谷16由0.150降为0.139,济谷18由0.153降为0.143;|a|值先减小后增大,济谷16变化范围为7.043—4.548,济谷18变化范围为7.356—4.932。对济谷16和济谷18方程的系数b、a进行差异显著性t检验,tb16=62.88,ta16=-18.12,tb18=70.98,ta18=-19.85(t0.01=3.355,n=8),结果表明,各系数间存在极显著性差异,说明不同品种在不同播期条件下对应的直线回归方程之间的差异达到极显著水平,不可以配置出一个共用的直线回归方程来表示济谷16、济谷18在不同播期条件下幼穗分化时期与叶龄指数的直线回归关系。
由直线方程的性质可知,公式X= (2-a)/b代表各个播期条件下谷子幼穗分化开始(Ⅱ期)时叶龄指数的理论值,随着播期的推迟表现为先减小后增大的趋势,与田间结果相一致,表明播期对谷子幼穗分化开始时的叶龄指数有显著影响;公式Y= 6/b代表谷子幼穗分化起止时期叶龄指数之差的理论值,随着播期的推迟表现为逐渐增大的趋势,而幼穗分化的持续时间又是逐渐减小的,表明播期对谷子幼穗分化过程中叶龄指数的变化幅度和变化速率有显著影响。
Table 3
表3
表3幼穗分化与叶龄指数的线性回归分析
Table 3The liner regression analysis of panicle differentiation and leaf number index
| 处理 T | 济谷16 Jigu16 | 济谷18 Jigu18 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| b | X | + a | R2 | F | (2-a)/b | 6/b | b | X | + a | R2 | F | (2-a)/b | 6/b | ||
| Y1= | 0.150 | X1 | -7.043 | 0.979 | 285.4 | 60.29 | 40.00 | 0.153 | X1 | -7.356 | 0.977 | 257.3 | 61.15 | 39.22 | |
| Y2= | 0.146 | X2 | -5.980 | 0.991 | 668.4 | 54.66 | 41.10 | 0.148 | X2 | -6.365 | 0.994 | 947.5 | 56.52 | 40.54 | |
| Y3= | 0.143 | X3 | -5.326 | 0.984 | 379.8 | 51.23 | 41.96 | 0.147 | X3 | -5.873 | 0.991 | 690.7 | 53.56 | 40.82 | |
| Y4= | 0.135 | X4 | -4.748 | 0.996 | 1372.8 | 49.99 | 44.44 | 0.140 | X4 | -5.304 | 0.997 | 2022.8 | 52.17 | 42.86 | |
| Y5= | 0.135 | X5 | -4.548 | 0.989 | 526.0 | 48.50 | 44.44 | 0.140 | X5 | -5.146 | 0.993 | 873.6 | 51.04 | 42.86 | |
| Y6= | 0.135 | X6 | -4.746 | 0.993 | 815.6 | 49.97 | 44.44 | 0.139 | X6 | -4.932 | 0.992 | 782.1 | 49.87 | 43.17 | |
| Y7= | 0.135 | X7 | -4.959 | 0.995 | 1292.2 | 51.55 | 44.44 | 0.139 | X7 | -5.122 | 0.996 | 1340.9 | 51.24 | 43.17 | |
| Y8= | 0.134 | X8 | -5.153 | 0.996 | 1372.2 | 53.38 | 44.78 | 0.137 | X8 | -5.360 | 0.997 | 2313.2 | 53.72 | 43.80 | |
| t-test | 62.80** | -18.12** | 39.78** | 64.87** | 70.98** | -19.85** | 41.51** | 72.81** | |||||||
新窗口打开
3 讨论
3.1 播期对夏谷幼穗分化的影响
谷子的花序为类穗状圆锥花序,单个谷穗上的近万个小穗和刚毛均着生于三级枝梗的末端,其幼穗分化过程远较水稻、小麦等其他禾谷类作物复杂[28-29]。段胜军等[11]研究发现,春、夏谷之间穗分化的方式及形态特征无明显差异,但夏谷营养生长期明显缩短,叶片分化数显著减少,幼穗分化持续时间也较短,并根据小穗、小花的特点将谷子穗分化过程划分为五个阶段沿用至今。本研究与前人研究结果基本一致,但之前有关****的观察手绘草图或实物照片不够全面完整[25,30-31],本研究更为清晰完整的阐述了谷子的整个幼穗分化过程。温度、水分、光照是影响谷子生长发育的重要环境因素[32-34]。播期越早,谷子生育前期的平均日温就越低,生育期也就越长,随着播期的推迟,温度逐渐升高,光照强度增大、降水增多,谷子的生育期也就越短[30]。春播谷子的穗分化开始在拔节之后的现象是较普遍且易观察到的,夏播穗分化多始于拔节之前,晚春播或早夏播能观察到穗分化与拔节同时开始的情况,这可能与光照反应有关[10,35]。本研究中,随着播期的推迟,夏谷的生育期由109 d减少到83 d左右,其中,幼穗分化开始时间由出苗后32 d减少到22 d、由拔节后7 d减少到1 d,幼穗分化阶段的持续时间由28 d减少到20 d,抽穗期到成熟期由49 d减少41 d,表明播期是影响谷子生育期的关键因素,播期越晚,出苗到幼穗分化开始以及幼穗分化过程的持续时间就越短,从而大幅缩短谷子的生育期。
3.2 播期对夏谷叶龄及叶龄指数的影响
随着播期的改变,谷子幼穗分化各个阶段的叶龄和叶龄指数表现出不同的变化趋势,品种之间的差异也较为明显,主要是由于谷子生育期、叶片生长速率、品种特性之间的差异造成的[36-37]。李存东等[7]曾指出,播期间叶龄进程的差异是其阶段发育进程差异的外在表现,而叶龄指数反映了叶片出生的相对进程,具有较好的可比性。本研究揭示,播期对夏谷幼穗分化过程中的叶龄、叶龄指数、叶片增长速率、叶龄指数变化幅度有很大影响,而叶龄指数比叶龄更适合作为衡量幼穗分化进程的指标。3.3 播期对夏谷幼穗分化与叶龄指数关联的影响
谷子幼穗分化与叶龄指数之间的线性回归关系已被广泛研究证实,利用直线回归方程Y=bX+a,通过叶龄指数可以准确判断谷子的幼穗发育阶段,而不同品种对应的回归函数系数略有不同,其中b值的大小与总生育期天数呈正相关[10,38]。本研究的线性回归分析结果显示,在不同播期条件下,夏谷幼穗分化与叶龄指数之间均存在真实的线性关系,而不同播期、不同品种所对应的回归函数差异显著,用叶龄指数推算谷子幼穗分化阶段的方法可行,但是在实际应用过程中,还要进一步研究,获得相匹配的公式。4 结论
2个夏谷品种穗分化发育形态基本一致,但播期对2品种幼穗分化的影响有所不同。播期延迟导致幼穗分化各阶段时间缩短,幼穗分化时间的差异是播期影响谷子生育期的主要原因。受播期的影响,穗分化不同阶段对应的叶龄和叶龄指数也有所差异,随播期的延迟,穗分化各阶段的叶片数逐渐减少,而对应的叶龄指数呈先下降再上升的趋势。致谢:谷子穗分化材料的观察照相利用山东省农业科学院生物技术中心李国卫博士实验室的数显体视显微镜完成,在此表示感谢!
The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}