0 引言
【研究意义】二化螟(Chilo suppressalis)是危害水稻最重要的害虫之一,广泛分布于亚欧大陆。二化螟在苗期危害水稻可造成枯鞘、枯心和虫伤株,在孕穗期和穗期危害可造成枯孕穗和白穗等,严重影响水稻的质量和产量[1-3]。近年来,种植结构和耕作制度调整,杂交稻面积逐年增加[4],使越冬代有效虫源面积扩大,更加剧了二化螟的危害[5-6]。目前,多采用化学药剂对二化螟进行防治[7]。但是,化学农药的不合理使用,导致二化螟抗药性不断提高,加大了二化螟的猖獗危害。同时,化学农药带来的农药残留超标、环境污染等问题也日益突出。这些不利因素促使人们迫切寻求环境友好的作物保护方式[8]。【前人研究进展】香根草(Vetiveria zizanioides)又名岩兰草,是一种多年生草本植物,常用于生态环境治理、水土保持等方面。已有研究报道,香根草可以作为玉米田诱集植物引诱玉米禾螟(Chilo partellus),从而降低其对玉米的危害[9]。室内盆栽试验表明,香根草具有引诱稻螟雌成虫产卵的特性,二化螟在香根草上的产卵量是水稻上的4倍左右[10-11]。田间防控效果调查发现,种植香根草的小区稻螟造成的枯心率比对照区降低了50%以上[10]。此外,田埂种植香根草除了可诱集二化螟成虫在其上产卵外,且孵化后的幼虫在香根草上不能完成生活史,仅有极少数的幼虫能存活至2龄、3龄,从而显著降低了二化螟种群数量,减少了其对水稻的危害[11]。因此,香根草作为诱集植物防治稻螟已成为水稻害虫绿色防控的重要措施之一,具有很大的开发潜力[12-14]。诱集植物根据其不同特征可分为传统型诱集植物、致死型诱集植物和基因工程诱集植物[15]。致死型诱集植物是指该诱集植物对害虫极具吸引力,害虫一旦取食或者在其上产卵,害虫及其后代将无法生存[16]。【本研究切入点】对二化螟而言,香根草属于典型的致死型诱集植物,但香根草对二化螟幼虫的致死作用机制尚未见报道。为更好地建立以香根草为基础的水稻螟虫绿色防控新技术,明确其对二化螟幼虫的致死作用机制尤为重要[8]。【拟解决的关键问题】通过测定香根草提取物组分对二化螟幼虫的生物活性,比较香根草和水稻中的营养物质含量及取食水稻和香根草后二化螟幼虫体内解毒酶和消化酶活性的变化,从毒理学和营养学两方面揭示香根草对二化螟幼虫致死的作用机制,为进一步深入了解其致死作用机制及建立稻螟绿色防控新技术提供重要的理论依据,为化学农药的减施、水稻害虫的可持续治理及水稻的安全生产提供有力保障。1 材料与方法
试验于2015—2016年在浙江省农业科学院完成。1.1 供试植物
水稻品种浙优12号购自浙江省农科种业有限公司。播种于温室塑料槽内,约30 d后移栽至直径为12 cm的塑料盆中,至45 d分蘖期时剪取茎秆以供营养物质测定及二化螟3龄幼虫的饲喂试验。香根草种苗购自深圳市鑫淼森园林草坪有限公司。将香根草种苗种于温室,以分蘖苗作为繁殖体,待香根草分蘖较多时剪取茎秆供营养物质测定及二化螟3龄幼虫的饲喂试验。
1.2 供试昆虫
二化螟种群于2015年5月采自杭州市萧山区义桥镇(30°04′ N,120°12′ E)水稻田,室内建立并维持种群,幼虫采用人工饲料饲养,饲料配方由中国水稻研究所提供[17],置于温度(27±2)℃,相对湿度(70±5)%,光周期16L﹕8D的智能人工气候室内饲养。待二化螟化蛹后将蛹收集起来,放置在干净的养虫笼中,保湿,让其继续发育羽化,对羽化后的成虫以10%的蜂蜜水提供营养。选取第1代二化螟3龄幼虫用于香根草活性物质的生物测定及水稻和香根草的饲喂试验。
1.3 香根草活性物质的制备及其对二化螟幼虫的生物活性测定
香根草活性物质的制备:称取0.5 kg香根草茎秆,阴干后用剪刀剪为长约2 cm的小节,置于多功能提取浓缩罐中,加80%的工业乙醇12 L室温浸泡20 h后,80℃恒温提取8 h,浓缩后得到乙醇提取物。将残渣置于中药煎药机,加蒸馏水6 L煎煮1.5 h,得水提取液。合并提取液,过滤浓缩蒸干得浸膏。试验重复两次,合并得总提取物36.6 g。取24.4 g浸膏制成1 L混悬液,依次用等体积石油醚、乙酸乙酯、正丁醇萃取3次,分别浓缩得到石油醚萃取物6.0 g,乙酸乙酯萃取物4.3 g,正丁醇萃取物5.0 g和萃取后母液蒸干得到的水部位8.6 g,分别用于测定其对二化螟3龄幼虫的生物活性。石油醚萃取物的硅胶柱层析:按上述方法称取1 kg香根草茎秆制备石油醚萃取物8.0 g,利用硅胶(100—200目)柱层析,分别以石油醚、石油醚:三氯甲烷(2﹕1)、石油醚:三氯甲烷(1﹕4)、三氯甲烷、乙酸乙酯各3 L梯度洗脱,分别收集流份减压浓缩干燥,获得5部分组合物,编号为Fr1—Fr5,分别测定其对二化螟3龄幼虫的生物活性。
香根草提取物的生物活性测定:采用饲料混毒法[18]。利用丙酮将香根草各提取物进行超声溶解,然后加入到人工饲料中配制成浓度为0.05 g·mL-1的含样品饲料,将饲料切成大小形状一致的小块,放入无菌的6孔培养板中,每孔接入5头3龄幼虫,覆盖吸水纸及盖子防止试虫逃逸。每板为1个重复,共重复3次,总计90头试虫。对照采用混入相应比例丙酮的常规人工饲料。所有处理均置于人工气候室内饲养(条件设置同试虫饲养),每3 d记录试虫死亡情况,共记录15 d,以毛笔轻触不动的幼虫记为死亡,其中Fr1—Fr5记录3 d的死亡率。
1.4 水稻和香根草中营养物质含量测定
将剪取的水稻和香根草茎秆自然阴干,分别磨成粉末状。水溶性蛋白含量测定:采用考马斯亮蓝法[19]。每个样品称取100 mg,加入1 mL pH 7.0的磷酸缓冲液(PBS),充分匀浆后,50℃振荡2 h,然后在4℃,12 000 ×g条件下离心15 min,取40 μL上清液,利用酶标仪(Infinite M200,Tecan)在595 nm下测定水稻和香根草样品中的蛋白含量。以牛血清白蛋白为标准蛋白制作标准曲线,计算各样品中蛋白含量,每个处理重复3次。
纤维素含量测定:采用比色法测定。准确称取样品粉末100 mg,加入100 mL预冷的60%的硫酸(H2SO4),在冰上消化处理30 min,过滤。取滤液2 mL于15 mL的玻璃试管中,加0.5 mL 2%蒽酮试剂和5 mL浓H2SO4,盖上塞子,沸水浴中加热10 min,取出冷却。取250 μL反应液在620 nm下测定OD值,以纤维素标准曲线计算各样品的纤维素含量,每个处理重复3次。
总糖含量测定:采用容量分析法测定。称取样品干粉100 mg,送至农业部农产品及转基因产品质量安全监督检测测试中心完成(杭州),每个处理重复3次。
氨基酸含量测定:采用离子交换色谱法测定。准确称取样品300 mg,送至农业部农产品及转基因产品质量安全监督检测测试中心完成(杭州),每个处理重复3次。
单宁含量测定:采用单宁含量试剂盒(苏州科铭生物技术有限公司)按说明书测定。分别称取水稻和香根草样品粉末100 mg,加入1 mL蒸馏水,充分匀浆后,80℃水浴提取30 min,然后在25℃,8 000×g条件下离心10 min,取15 μL上清液,利用酶标仪分别测定两个样品在760 nm下的吸光度,根据说明书计算单宁含量。每个处理重复3次。
1.5 取食水稻和香根草后二化螟幼虫体内解毒酶活性测定
利用水稻和香根草茎秆饲喂二化螟3龄幼虫,分别于饲喂3、6、9、12和15 d后取样,幼虫饥饿3 h后,在液氮中速冻并立即转置-80℃冰箱中保存,供后续酶活性测定试验用。每处理收集幼虫5头,共3次重复。羧酸酯酶(CarE)活性测定:参照LU等[20]的方法稍加改进。用0.1 mol·L-1,pH 7.0的PBS制备酶液。每个样品称取100 mg,加入pH 7.0的PBS 1 mL,充分匀浆后,在4℃,12 000×g条件下离心15 min,取40 μL上清液加入96孔酶标板中,每孔加入100 μL底物与显色剂的混合液(用0.2 mol·L-1、pH 6.0的PBS,配制含10 mmol α-乙酸萘酯和1 mmol固蓝RR盐,混合后经过滤得到的滤液即为底物与显色剂的混合液)。然后加入5 μL酶液和45 μL 0.1 mol·L-1、pH 7.6的PBS,利用动力学方法测定10 min内OD450值变化,以反应速度表示酶活力(mol·min-1·mg-1 protein)。
谷胱甘肽-S-转移酶(GSTs)活性测定:参照HABIG等[21]的方法稍加改进。用0.1 mol·L-1,pH 6.5的PBS制备酶夜,在96孔酶标板中每孔依次加入45 μL 0.1 mol·L-1、pH 7.6的磷酸缓冲液(PBS)、5 μL酶液(制备方法同蛋白酶)、50 μL 1.2 mmol·L-1的1-氯-2,4-二硝基苯(CDNB)和50 μL 6 mmol·L-1的还原型谷胱甘肽(GSH)。利用动力学方法测定10 min内OD340值变化,以反应速度表示酶活力(nmol·min-1·mg-1 protein)。
细胞色素P450酶活性测定:参考ROSE等[22]的方法稍加改进。利用0.1 mol·L-1、pH 7.6的PBS(含1 mmol·L-1 EDTA,1 mmol·L-1 DTT,1 mmol·L-1苯基硫,1 mmol·L-1 PMSF)制备酶液。在96孔酶标板中依次加入2 μmol·L-1对硝基苯甲醚(溶解于0.1 mol·L-1 pH 7.8的磷酸缓冲液)50 μL,9.6 mmol·L-1的NADPH 5 μL和45 μL制备好的酶液。利用动力学方法测定10 min内OD405值变化,以反应速度表示酶活力(nmol·min-1·mg-1 protein)。
1.6 取食水稻和香根草后二化螟幼虫体内消化酶活性测定
样品处理及收集同1.5。蛋白酶活力的测定:参考曹挥等的方法测定[23-24]。将收集的二化螟幼虫转移至预冷的组织研磨器中,加入1 mL pH 7.0 PBS(含0.3%的Triton X-100)研磨,在4℃,12 000×g条件下离心15 min,取上清液作为酶源。取20 µL酶液,在37℃放置5 min,加入50 µL 37℃预热的0.5%酪蛋白溶液,混匀后37℃放置15 min,然后加入50 µL 10%的三氯乙酸,混合后离心(10 000×g,4℃,15 min),取上清液20 µL,放入酶标板内,加入100 µL 0.55 mol·L-1 Na2CO3,20 µL Folin-酚试剂,室温放置30 min,利用酶标仪在680 nm下测定OD值。以酪氨酸标准曲线计算蛋白酶活性。
淀粉酶活力测定:参照张炬红等[24]的方法测定。取10 µL 2%的淀粉,20 µL 0.2 mol·L-1 PBS(pH 6.0),10 µL酶液(制备方法同蛋白酶),依次加入1.5 mL的离心管内,室温下反应10 min后,37℃水浴60 min,加250 µL 3,5-二硝基水杨酸终止反应,然后沸水浴5 min,自然冷却后,从中取出250 µL样品加在酶标板内,用酶标仪于550 nm波长测定OD值。以麦芽糖标准曲线计算淀粉酶活性。
海藻糖酶活力测定:参考张炬红等[24]的方法。取10 µL 3%的海藻糖、20 µL 0.2 mol·L-1 PBS(pH 5.5)和10 µL酶液(制备方法同蛋白酶),依次加入1.5 mL的离心管内,室温下反应10 min后,37℃中水浴60 min,加入250 µL 3,5-二硝基水杨酸终止反应,然后在沸水浴中放置5 min,自然冷却后从中取出250 µL样品加在酶标板内,在酶标仪550 nm下测定OD值。以葡萄糖标准曲线计算海藻糖酶活性。
蔗糖酶活力测定:参考张炬红等[24]的方法。取10 µL 4%的蔗糖、20 µL 0.2 mol·L-1 PBS(pH 5.5)和10 µL酶液(制备方法同蛋白酶),依次加入1.5 mL的离心管内,室温下反应10 min后,37℃中水浴60 min,随后加入250 µL 3,5-二硝基水杨酸终止反应,然后沸水浴持续5 min,自然冷却后从中取出250 µL样品加在酶标板内,用酶标仪在550 nm下测定OD值。以葡萄糖标准曲线计算蔗糖酶活性。
1.7 数据统计与分析
采用Microsoft Excel 2010处理数据,采用DPS 14.50软件进行差异显著性分析(P≤0.05)[25]。2 结果
2.1 香根草活性物质提取及对二化螟3龄幼虫的生物活性测定
香根草总提取物、水部位、石油醚萃取物、乙酸乙酯萃取物和正丁醇萃取物5种组分对二化螟3龄幼虫的生物活性测定结果表明,乙酸乙酯萃取物处理二化螟3龄幼虫15 d后,死亡率为1.85%,与对照无显著差异;香根草粗提物、水部位和正丁醇萃取物处理后随时间延长,二化螟死亡率逐渐升高,但直至处理15 d后,死亡率均低于20%;石油醚萃取物处理9、12、15 d后,二化螟死亡率分别为38.89%、57.41%、85.19%(图1)。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1香根草提取物对二化螟3龄幼虫的生物活性标准误由3次生物学重复计算得出。统计分析采用DPS的One-Way ANOVA-Fisher’s LSD方法(P≤0.05)。不同小写字母表示香根草不同提取物对二化螟3龄幼虫的致死率具有显著差异。
-->Fig. 1Bioassay of vetiver extracts against the 3rd instar larvae of C. suppressalisStandard error bars were based on three replicates. One-Way ANOVA-Fisher’s LSD method of DPS software was used for statistical analysis (P≤0.05). Different lowercases indicated significant differences among mortalities of the 3rd instar larvae of C. suppressalis. The same as
-->
进一步将石油醚萃取物通过硅胶柱层析,获得编号为Fr1—Fr5的5个组分。生测结果表明,处理3 d后,Fr1对二化螟的死亡率为85.00%,显著高于其他组分。Fr3和Fr5对二化螟幼虫的死亡率分别为48.33%和66.67%,二者无显著差异;Fr2对二化螟幼虫的死亡率为30.00%,与Fr3无显著差异;Fr4对二化螟幼虫的死亡率最低,仅为8.33%(图2)。
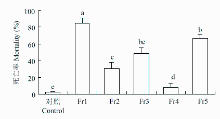 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2香根草石油醚萃取物中不同组分对二化螟3龄幼虫的生物活性
-->Fig. 2Bioassay of sherwood oil extracts of vetiver against the 3rd instar larvae of C. suppressalis
-->
2.2 水稻和香根草茎秆中营养物质含量
除单宁外,水稻中其他营养物质含量均显著高于香根草中的含量。其中水稻和香根草中总蛋白的含量分别为13.26和9.45 μg,纤维素含量分别为2.92%和1.61%,总糖含量分别为9.60%和1.45%(图3)。而单宁在香根草中的含量是水稻中含量的1.31倍(图3)。此外,从水稻和香根草中均鉴定出17种氨基酸,总含量分别为14.23%和4.00%,且这17种氨基酸在水稻中的含量为香根草中含量的2.5—7.0倍。其中水稻中蛋氨酸含量是香根草中的7.0倍(表1)。说明除单宁外,水稻中各种营养物质含量均比香根草中丰富。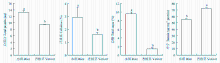 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3水稻和香根草中的营养物质含量统计分析采用DPS的t-test方法(P≤0.05)。不同小写字母表示同一种营养物质在水稻和香根草中的含量具有显著差异
-->Fig. 3Nutrient contents in rice and vetiver t-test was used for statistical analysis (P≤0.05) using DPS software. Different lowercases indicated significant differences of nutrient contents between rice and vetiver
-->
Table 1
表1
表1水稻和香根草茎秆中氨基酸种类和含量
Table 1Amino acid compositions and contents in rice and vetiver stem
| 氨基酸 Amino acid | 氨基酸含量 Content of amino acid (%) | 水稻/香根草Rice/Vetiver | |
|---|---|---|---|
| 水稻Rice | 香根草Vetiver | ||
| 天门冬氨酸ASP | 1.86±0.04a | 0.59±0.10b | 3.2 |
| 苏氨酸THR | 0.69±0.03a | 0.20±0.02b | 3.5 |
| 丝氨酸SER | 0.76±0.06a | 0.23±0.03b | 3.3 |
| 谷氨酸GLU | 2.10±0.11a | 0.52±0.06b | 4.0 |
| 脯氨酸PRO | 0.68±0.02a | 0.21±0.02b | 3.2 |
| 氨NH3 | — | — | — |
| 甘氨酸GLY | 0.82±0.06a | 0.24±0.03b | 3.4 |
| 丙氨酸ALA | 1.09±0.07a | 0.30±0.04b | 3.6 |
| 胱氨酸CYS | — | — | — |
| 缬氨酸VAL | 0.86±0.03a | 0.24±0.03b | 3.6 |
| 蛋氨酸MET | 0.14±0.00a | 0.02±0.00b | 7.0 |
| 异亮氨酸ILE | 0.63±0.03a | 0.17±0.03b | 3.7 |
| 亮氨酸LEU | 1.28±0.08a | 0.34±0.05b | 3.8 |
| 酪氨酸TYR | 0.34±0.04a | 0.09±0.01b | 3. 8 |
| 苯丙氨酸PHE | 0.78±0.06a | 0.22±0.02b | 3.6 |
| 组氨酸HIS | 0.53±0.01a | 0.21±0.04b | 2.5 |
| 赖氨酸LYS | 0.91±0.06a | 0.29±0.04b | 3.1 |
| 精氨酸ARG | 0.80±0.07a | 0.18±0.02b | 4.4 |
| 总量Total (%) | 14.23±0.72a | 4.00±0.49b | 3.6 |
新窗口打开
2.3 取食水稻和香根草后二化螟幼虫体内解毒酶活性
对取食水稻和香根草后二化螟幼虫体内解毒酶活性测定结果表明,取食不同寄主植物9、12、15 d后,取食水稻的幼虫体内CarE活性显著高于取食香根草的幼虫体内CarE活性;GSTs在取食水稻和香根草的幼虫体内无显著差异;细胞色素P450酶活性则在取食不同寄主植物6—15 d,二者均呈现显著差异(图4)。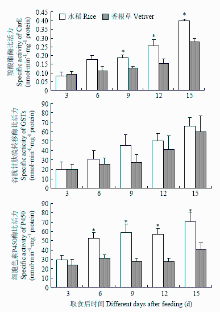 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4二化螟3龄幼虫取食水稻和香根草后体内解毒酶活性变化星号表示取食水稻和香根草的二化螟3龄幼虫酶活力存在显著差异(t-test,P≤0.05)。
-->Fig. 4Activity changes of detoxifying enzymes after the 3rd instar larvae feeding on rice and vetiver stemsAsterisks indicated significant differences between the enzyme activities in 3rd instar larvae fed on rice and vetiver stems (t-test, P≤0.05). The same as
-->
2.4 取食水稻和香根草后二化螟幼虫体内消化酶活性比较
分别取食不同寄主植物3、6、9、12、15 d后,取食香根草的二化螟幼虫体内蛋白酶、淀粉酶、海藻糖酶和蔗糖酶活性均显著低于取食水稻的二化螟幼虫体内相应酶的活性(图5)。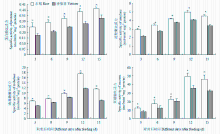 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5二化螟3龄幼虫取食水稻和香根草后体内消化酶活性变化
-->Fig. 5Activity changes of digestive enzymes in the 3rd instar larvae after fed on rice and vetiver stems
-->
3 讨论
香根草作为诱集植物防治二化螟已成为稻螟绿色防控的重要措施之一,具有很大的开发潜力[12,14,26-27]。笔者实验室前期研究结果表明,香根草能诱集二化螟雌成虫在其上大量产卵,但孵化出的幼虫在香根草上不能存活[11],因此香根草对二化螟而言属于典型的致死诱集型植物。如果能够开发鉴定出香根草中的杀虫活性物质,明确其对二化螟幼虫致死的作用机制,将为以香根草为基础的稻螟绿色防控新技术提供更充分的理论依据。本研究从解毒代谢和营养代谢两方面揭示了香根草对二化螟幼虫的致死作用机制。首先,发现香根草的石油醚萃取物对二化螟幼虫具有很强的毒杀作用,15 d后幼虫死亡率高达85.19%,说明香根草中含有对二化螟幼虫具有毒杀作用的活性成分,且存在于石油醚萃取物中;进一步对石油醚萃取物进行分离,在获得的5种组分中,Fr1和Fr5对二化螟幼虫具有较高的致死作用,死亡率分别为85%和67%。笔者实验室前期研究表明,香根草石油醚提取物对二化螟初孵幼虫的毒性较高,3 d后死亡率高达84%左右[13]。这均说明香根草中的杀虫活性物质主要存在于Fr1和Fr5组分中,这些有毒物质进入二化螟体内后可能通过抑制其解毒代谢酶活性而降低了对有毒物质的代谢能力,最终引起昆虫中毒死亡。研究还表明,昆虫取食不同寄主植物后对其解毒代谢酶及相关代谢能力的影响不同[28]。昆虫体内参与外来物质代谢的解毒酶主要包括CarE、GSTs和细胞色素P450酶等酶系。本研究表明,香根草中含有对二化螟幼虫具杀虫活性的有毒物质,二化螟取食后,幼虫体内CarE和细胞色素P450酶活性受到显著抑制,导致其对香根草有毒物质的解毒代谢能力下降,引起昆虫死亡。但Fr1和Fr5只是活性成分的粗提物,究竟是哪些活性物质发挥了杀虫作用目前还不能确定,有待进一步研究证实。
此外,香根草中营养成分匮乏可能是二化螟幼虫不能存活的另一重要因素。本研究表明,香根草中总蛋白、纤维素、总糖、氨基酸等营养物质的含量均显著低于水稻中相应物质的含量,而这些营养物质是二化螟生长发育所必需的。一般来讲,植食性昆虫通过取食寄主植物获得营养物质,满足其生长发育的基本营养物质有糖类、蛋白质、氨基酸、纤维素等。其中糖类主要为昆虫的生命活动提供能量需求,纤维素是昆虫的主要碳水化合物[29-30];蛋白质和氨基酸的利用会影响昆虫取食、生长和繁殖[30-31]。香根草中单宁含量高可能是二化螟取食后营养吸收受阻的一个重要因素。单宁对昆虫生长发育具有强烈抑制作用,是常见的植物抗虫物质。单宁与消化酶结合,可抑制消化酶活性,影响其正常生理功能,进而阻碍昆虫对营养成分的吸收[32]。所以,取食香根草的二化螟幼虫可能由于取食植物中的有毒物质或无法利用植物中的营养物质,造成由于营养物质缺乏,结合有毒物质的毒杀作用,最终导致其死亡。
4 结论
香根草中有毒物质抑制了二化螟体内CarE和细胞色素P450酶的活性;香根草内营养物质缺失,单宁含量高,从而影响二化螟体内消化酶活性的变化,这些可能是二化螟幼虫取食香根草后致死的两大重要因素。研究结果可为开发以香根草为基础的稻螟绿色防控新技术提供理论依据。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}