 ,, 张函青, 李萍, 张东升, 林文, 薛建福, 高志强, 郝兴宇,山西农业大学农学院,山西太谷 030801
,, 张函青, 李萍, 张东升, 林文, 薛建福, 高志强, 郝兴宇,山西农业大学农学院,山西太谷 030801Effects of Elevated Atmospheric CO2 Concentration and Temperature on Photosynthetic Characteristics, Carbon and Nitrogen Metabolism in Flag Leaves and Yield of Winter Wheat in North China
ZONG YuZheng,, ZHANG HanQing, LI Ping, ZHANG DongSheng, LIN Wen, XUE JianFu, GAO ZhiQiang, HAO XingYu,College of Agriculture, Shanxi Agricultural University, Taigu 030801, Shanxi通讯作者:
责任编辑: 杨鑫浩
收稿日期:2021-02-18接受日期:2021-07-5
| 基金资助: |
Received:2021-02-18Accepted:2021-07-5
作者简介 About authors
宗毓铮,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (567KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
宗毓铮, 张函青, 李萍, 张东升, 林文, 薛建福, 高志强, 郝兴宇. 大气CO2与温度升高对北方冬小麦旗叶光合特性、碳氮代谢及产量的影响. 中国农业科学, 2021, 54(23): 4984-4995 doi:10.3864/j.issn.0578-1752.2021.23.005
ZONG YuZheng, ZHANG HanQing, LI Ping, ZHANG DongSheng, LIN Wen, XUE JianFu, GAO ZhiQiang, HAO XingYu.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】我国是世界上小麦(Triticum aestivum)种植面积最大、产量最高的国家,种植面积高达 2 400万hm2,占世界小麦总产量的17.6%[1]。我国北方是冬小麦主产区,小麦产量变化会对中国及世界粮食安全产生巨大影响。气候条件是影响小麦产量形成的重要因素。工业革命以来,大气CO2浓度不断升高,目前已达410 μmol·mol-1,到21世纪末将达421—936 μmol·mol-1[2]。大气中CO2等温室气体浓度升高将引发气温持续升高。据预测,到2080s全球平均气温将比工业革命前增加2℃左右[3]。然而,目前关于大气CO2浓度与温度同时升高的交互作用对小麦产量形成的认识仍不十分清楚。【前人研究进展】已有大量研究证明,CO2浓度升高将减小叶片气孔导度、降低蒸腾速率,提高叶片水分利用效率[4]。另外,在水分与养分正常供应下,CO2浓度升高可提高大多数作物叶片光合碳同化能力[4,5],改变植株体内物质运输与分配格局[6,7],提高产量[4]。位于美国亚利桑那州、伊利诺伊州与我国北京昌平等多个FACE平台的长期试验结果均表明,在水分与养分正常供应条件下,大气CO2浓度升高将使小麦产量提高约19%—31%[4, 8-10]。气温升高会缩短禾谷类作物籽粒灌浆时间,导致灌浆不充分,收获指数降低,尤其对热带地区影响更大[11,12]。ASSENG等[12]发现开花期升温0.6℃使小麦减产3.6%,开花期温度超过30℃将引起产量大幅度降低[13]。生育期气温持续升高1—2℃将会使非洲玉米产量下降超过50%[14,15]。同时,升温也会导致高温热害发生的概率增加,对作物生长发育及产量造成不利影响[16,17]。但是,目前关于大气CO2浓度与温度同时升高下小麦生理生态适应与响应特征的研究仍然较少。前人通过整合分析分布于全球5个大洲14个FACE观测位点186项研究的数据表明,在没有非生物胁迫条件下,18种C3作物的产量在单一大气CO2浓度升高环境中将平均提高18%,但在CO2与温度(增温2℃)同时持续升高的交互作用下仅提高约10%[4]。在我国,有研究表明,在CO2浓度为560 μmol·mol-1条件下,气温升高1.7℃对北方冬小麦没有显著影响[11];在CO2浓度为500 μmol·mol-1条件下,气温升高1.5—2℃将使我国南方小麦减产10%—12%[18]。【本研究切入点】我国目前的研究仍主要集中于CO2浓度单因子变化对小麦产量与品质形成过程的影响,对其与温度升高双因子的研究仍然较少,结果也存在争议。且升温大多是基于红外加热装置进行,而红外加热在小麦拔节以后主要是增加小麦冠层叶片温度,其他下层组织难以达到目标温度,这或将对试验结果产生影响。有必要开展整个植株升温的控制试验,以更精准研究大气CO2浓度和气温升高复合条件下小麦生长发育及产量变化。【拟解决的关键问题】本研究在人工气候生长室中,利用液态CO2钢瓶提供CO2以及空调控制气温,研究了大气CO2浓度升高200 μmol·mol-1和气温升高2℃条件下小麦生长发育、光合生理、生理代谢及产量构成的变化,为未来气候变化背景下小麦生产提供参考。1 材料与方法
1.1 试验材料及处理
供试小麦品种中科2011(Zhongke2011)由中国科学院遗传发育研究所培育。本试验在山西省晋中市山西农业大学农作站(37.42°N,112.58°E)进行。试验设置4个处理,分别为对照CK(CO2浓度和气温与外界大田环境相同)、EC(CO2浓度为大田浓度+200 μmol·mol-1,气温与外界大田相同)、ET(CO2浓度与大田环境相同,气温为大田温度+2℃)、ECT(CO2浓度为大田浓度+200 μmol·mol-1,气温为大田气温+2℃)。试验在人工控制气候室(铝合金框架玻璃结构,每个气室长8 m,宽3 m,高3.2 m)中进行,气室采用自然光,玻璃透光率80%—90%。利用自动控制系统(邯郸冀南新区盛炎电子科技有限公司)在小麦生长全生育期进行大气CO2浓度和温度控制,越冬期除外。CK处理与大田气温和CO2浓度变化一致(实时控制,包括日变化及季节变化),其他气室按设定(按CK处理作为参照)进行控制。2017年10月25日和2019年10月27日,在塑料箱中(长60 cm,宽40 cm,高35 cm,底部打5个孔)播种小麦种子(中科2011),箱内装土28 cm高。供试土壤为附近农田0—20 cm的表层土壤,经风干过筛混匀后装盆,土壤类型为褐潮土,土壤特性为pH 8.2,有机质含量为 2.45 g·kg-1,全氮含量 1.4 g·kg-1。播种时施底肥,施肥量按105.32 N kg·hm-2,65.49 g P2O5 kg·hm-2折算,拔节期追施氮肥105.32 N kg·hm-2。每个气室每个处理种植4箱,每箱播种2行小麦,每行60粒小麦,定期浇水,确保无干旱胁迫,分别于2018年5月30日和2020年6月5日收获。两个生长季各气室气温变化如表1。
Table 1
表1
表1两个生长季各处理气温变化
Table 1
| 年份 Year | 处理 Treatment | 生长季平均温度 Mean temperature of growing season (℃) | 最高温度及出现日期 Maximum temperature and its date | 最低气温及出现日期 Minimum temperature and its date | ||||
|---|---|---|---|---|---|---|---|---|
| 最高温度 Maximum temperature (℃) | 日平均温度 Mean daily temperature (℃) | 日期 Date (M-D) | 最低温度 Minimum temperature (℃) | 日平均温度 Mean daily temperature (℃) | 日期 Date (M-D) | |||
| 2017-2018 | CK | 11.4 | 38.3 | 28.7 | 06-07 | -11.9 | 0.2 | 02-04 |
| EC | 11.6 | 38.6 | 28.9 | 06-07 | -11.3 | -0.1 | 02-04 | |
| ET | 13.5 | 40.5 | 30.9 | 06-07 | -11.9 | 0.3 | 02-04 | |
| ECT | 13.6 | 40.8 | 30.9 | 06-07 | -11.8 | 0.2 | 02-04 | |
| 2019-2020 | CK | 12.4 | 40.3 | 28.7 | 06-04 | -10.9 | -0.2 | 12-31 |
| EC | 12.4 | 40.5 | 28.9 | 06-04 | -10.3 | 0.1 | 12-31 | |
| ET | 14.1 | 42.4 | 30.9 | 06-04 | -10.9 | 0.3 | 12-31 | |
| ECT | 14 | 42.5 | 31.0 | 06-04 | -9.9 | 0.3 | 12-31 | |
新窗口打开|下载CSV
1.2 测定指标
1.2.1 小麦发育期观测 记录小麦播种、出苗、开花、成熟时间。出苗:该处理所有整理箱内小麦幼苗露出土壤表面。开花:一半以上的小麦穗开花。成熟:所有的植株变黄,籽粒变硬。1.2.2 叶片光合色素含量及碳氮代谢指标的测定 在2020年小麦灌浆期(为保证发育期一致,ET处理、ECT处理在5月10日取样,其他处理于5月20日取样),选取长势相似的小麦旗叶剪下后液氮保存,每个处理每盆取5片叶,带回实验室-80℃冰箱保存,用于光合色素含量、可溶性糖、淀粉、纤维素、可溶性蛋白、硝酸还原酶活性的测定。采用分光光度计法测定光合色素,蒽酮-硫酸法测定可溶性糖、淀粉和纤维素含量,采用考马斯亮蓝法测定可溶性蛋白含量,采用离体法测定硝酸还原酶活性[19]。
1.2.3 气体交换的测定 在2018年拔节期(4月3日)、孕穗期(为保证发育时期一致,ET处理、ECT处理在4月12日,其他处理于4月18日测定)和灌浆期(ET处理、ECT处理在5月4日,其他处理于5月15日测定),2020年拔节期(4月8日)、孕穗期(ET处理、ECT处理在4月16日,其他处理于4月25日测定)和灌浆期(ET处理、ECT处理在5月10日,其他处理于5月20日测定),选择晴朗天气,各处理随机选取4株(每盆1株)长势一致的小麦,选择完全展开的最上面功能叶片(拔节期选倒二叶,其他发育期选择旗叶),测定净光合速率(Pn)、气孔导度(Gs)。测定仪器为便携式气体交换系统(Li-6400XT,美国Li-COR),测定时间每日9:00—11:30。叶室参数设定如下:光合有效辐射(PAR)1 600 μmol·m-2·s-1,VPD 1 500 kPa,流速500 μmol·s-1,参比室CO2浓度CK和ET处理为400 μmol·mol-1,EC和ECT处理为600 μmol·mol-1;叶室温度CK和EC处理24℃,ET和ECT处理26℃。
1.2.4 光响应曲线和CO2响应曲线的测量 2020年小麦灌浆期(ET处理、ECT处理在5月9日,其他处理于5月19日),每种处理随机选择3株长势一致的小麦旗叶,利用便携式光合测量系统(LI-6400XT)于每日9:00—14:00测定光响应曲线。叶室参数设定如下:VPD设为1 500 kPa,流速500 μmol·s-1,参比室CO2浓度CK和ET处理为400 μmol·mol-1,EC和ECT处理为600 μmol·mol-1;叶室温度CK和EC处理设为24℃,ET和ECT处理的叶室温度设为26℃;红蓝光光源控制光合有效辐射(PAR),光强梯度为 2 000、1 600、1 400、1 200、1 000、800、600、400、200、100、50、0 mmol·mol-1共12个不同的水平,分别测定不同光强下小麦旗叶的净光合速率(Pn),测定完成后对测得数值进行光响应曲线的模拟,并计算最大净光合速率(Pnmax)、光饱和点(LSP)、光补偿点(LCP)以及暗呼吸速率(Rd)[20]。
2020年小麦灌浆期(ET处理、ECT处理在5月11日,其他处理于5月21日),进行CO2响应曲线测定,叶室参数设置如下:光合有效辐射(PAR)1 600 μmol·m-2·s-1,VPD为1 500 kPa,流速为500 μmol·s-1,叶室温度CK和EC处理设为24℃,ET和ECT处理设为26℃,CO2浓度控制梯度变化为400、300、200、100、0、400、600、800、1 000、1 200 μmol·mol-1,测得小麦叶片的净光合速率(Pn)和胞间CO2浓度(Ci),利用测定数据进行CO2响应曲线的模拟,得出CO2补偿点(Γ)、饱和胞间CO2浓度(Cisat)、最大光合能力(Amax)、光呼吸速率(Rp),求得Rubisco最大羧化速率(Vc.max)、最大电子传递速率(Jmax),再计算Jmax/Vc.max[20,21]。
1.3 数据处理
采用Excel 2007对所有试验数据进行整理和图表绘制。运用SPSS23进行单因素ANOVA和Duncan法进行方差分析和多重比较(P=0.05)。2 结果
2.1 大气CO2浓度和气温升高对小麦生育期的影响
大气CO2浓度升高和气温升高均加快小麦生育进程,升温对生育期的影响大于CO2浓度升高的影响。大气CO2浓度升高使小麦收获期提前2—3 d,使2018年小麦开花到成熟的时间增加1 d,对2020年小麦开花到成熟的时间没有影响。升温使小麦收获期提前10—12 d,使小麦开花到成熟的时间缩短4 d。大气CO2浓度升高和气温升高复合条件下小麦收获期提前11—13 d,小麦开花到成熟的时间缩短2 d(表2)。Table 2
表2
表2CO2浓度和气温升高对小麦生育期的影响
Table 2
| 年份 Year | 处理 Treatment | 播种期 Sowing date (M-D) | 出苗时间 Emergence (d) | 开花时间 Flowering date (d) | 成熟期 Mature date (d) | 开花到成熟时间 The days from flowering to ripening (d) |
|---|---|---|---|---|---|---|
| 2017-2018 | CK | 10-25 | 9 | 180 | 219 | 39 |
| EC | 10-25 | 9 | 176 | 216 | 40 | |
| ET | 10-25 | 8 | 174 | 209 | 35 | |
| ECT | 10-25 | 8 | 171 | 208 | 37 | |
| 2019-2020 | CK | 10-27 | 9 | 185 | 225 | 40 |
| EC | 10-27 | 9 | 183 | 223 | 40 | |
| ET | 10-27 | 8 | 177 | 213 | 36 | |
| ECT | 10-27 | 8 | 174 | 212 | 38 |
新窗口打开|下载CSV
2.2 大气CO2浓度和气温升高对小麦叶片净光合速率和气孔导度的影响
2年的试验结果表明,大气CO2浓度升高提高了小麦拔节期和孕穗期叶片净光合速率,但灌浆期无显著影响。升温使小麦叶片净光合速率在拔节期与灌浆期无显著变化,在孕穗期显著增加24.7%。CO2浓度和温度升高对孕穗期净光合速率的交互作用显著,CO2浓度和气温升高的交互作用小于CO2浓度和升温的单独效应之和,气温升高会减弱孕穗期CO2浓度升高对小麦叶片净光合速率的正效应(图1)。图1
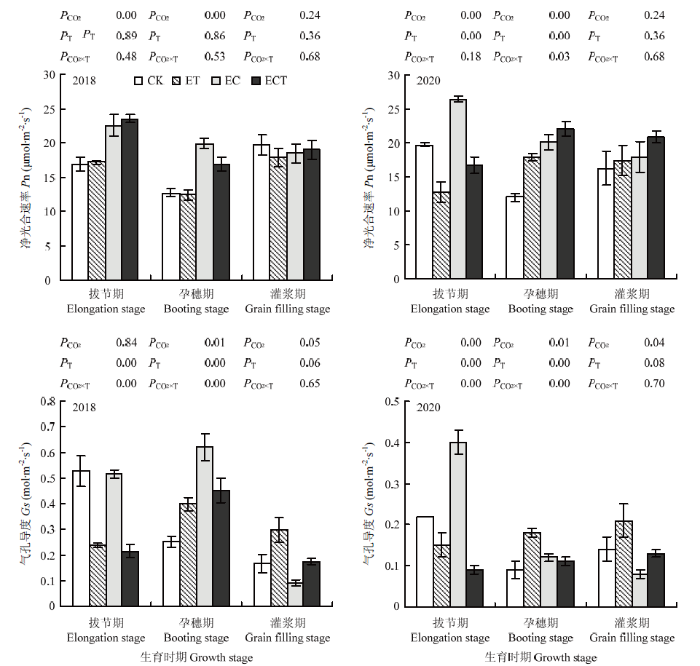 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1CO2浓度和气温升高对小麦净光合速率与气孔导度的影响
CK:CO2浓度和气温与外界大田环境相同;EC:CO2浓度为大田浓度+200 μmol·mol-1,气温与外界大田环境相同;ET:CO2浓度与大田环境相同,气温为大田气温+2℃;ECT:CO2浓度为大田环境浓度+200 μmol·mol-1,气温为大田气温+2℃。P<0.05表示差异显著;P<0.01表示差异极显著。下同
Fig. 1Effects of elevated CO2 and increased temperature on net photosynthetic rate (Pn) and stomatal conductance (Gs) of wheat
CK: Ambient CO2 concentration, ambient temperature; EC: Ambient concentration +200 μmol·mol-1; ET: Ambient temperature +2℃; ECT: Ambient concentration +200 μmol·mol-1, ambient temperature +2℃. P<0.05, significant difference; P<0.01, extremely significant difference. The same as below
随生育期推进,CO2浓度升高使气孔导度前期较高,后期显著降低;温度升高及其与CO2浓度升高的交互作用下气孔导度在生育期内变化幅度相对较小。CO2浓度和温度升高对拔节期和孕穗期叶片气孔导度的交互作用显著,这2个生育时期CO2浓度和气温升高的交互作用均小于CO2浓度和升温的单独效应之和,气温升高会减弱拔节期和孕穗期CO2浓度升高对小麦叶片气孔导度的正效应(图1)。
2.3 大气CO2浓度和气温升高对小麦叶片光响应相关参数的影响
大气CO2浓度升高提高了小麦灌浆期叶片光补偿点和最大净光合速率,增幅分别为54.1%和16.7%,对光饱和点和暗呼吸速率无显著影响。升温使灌浆期小麦叶片光饱和点下降38.3%,对光补偿点、最大净光合速率和暗呼吸速率无显著影响。CO2浓度和温度升高对叶片光补偿点的交互作用显著,CO2浓度和气温升高的交互作用大于CO2浓度和升温的单独效应之和,气温升高会提高灌浆期CO2浓度升高对小麦叶片光补偿点的正效应(表3)。虽然CO2浓度升高没有增加灌浆期叶片净光合速率,但是光补偿点与最大净光合速率增加,推测对产量形成起到一定作用。增温使光饱和点下降,推测对产量形成起到限制作用。Table 3
表3
表3CO2浓度和气温升高对小麦灌浆期叶片光响应参数的影响
Table 3
| 处理 Treatment | 光饱和点 Light saturation point (LSP) (μmol·m-2·s-1) | 光补偿点 Light compensation point (LCP) (μmol·m-2·s-1) | 最大净光合速率 Maximum net photosynthetic rate (Pnmax) (μmol·m-2·s-1) | 暗呼吸速率 Dark respiration rate (Rd) (μmol·m-2·s-1) | |
|---|---|---|---|---|---|
| CK | 2235.92±508.20ab | 12.83±0.07ab | 18.28±1.35b | 1.01±0.14a | |
| EC | 2554.62±146.51a | 10.47±5.17ab | 22.07±0.58a | 0.88±0.41a | |
| ET | 1475.33±118.13b | 4.19±0.52b | 19.99±0.28ab | 0.42±0.05a | |
| ECT | 1481±150.09b | 15.76±0.97a | 22.59±0.61a | 1.14±0.11a | |
| P-value | PT | 0.01 | 0.54 | 0.21 | 0.49 |
| PCO2 | 0.58 | 0.01 | 0.00 | 0.23 | |
| PT×CO2 | 0.59 | 0.03 | 0.48 | 0.09 |
新窗口打开|下载CSV
2.4 大气CO2浓度和气温升高对小麦叶片CO2响应相关参数的影响
大气CO2浓度升高对小麦灌浆期叶片最大光合能力、饱和胞间CO2浓度、CO2补偿点和光呼吸速率无显著影响。升温使灌浆期小麦叶片饱和胞间CO2浓度增加11.6%,对CO2补偿点、最大光合能力和光呼吸速率无显著影响。CO2浓度和温度升高对叶片饱和胞间CO2浓度的交互作用显著,CO2浓度和气温升高的交互作用大于CO2浓度和升温的单独效应之和,气温升高会减弱灌浆期CO2浓度升高对小麦叶片饱和胞间CO2浓度的负效应(表4)。Table 4
表4
表4CO2浓度和气温升高对小麦灌浆期叶片CO2响应参数的影响
Table 4
| 处理 Treatment | 饱和胞间CO2浓度 Saturated intercellular CO2 concentration (Cisat) (μmol·mol-1) | CO2补偿点 CO2 compensation point (Γ) (μmol·mol-1) | 最大光合能力 Photosynthetic capacity (Amax) (μmol·m-2·s-1) | 光呼吸速率 Photorespiration rate (Rp) (μmol·m-2·s-1) | |
|---|---|---|---|---|---|
| CK | 772.78±70.01a | 62.15±2.32a | 42.64±3.2a | 14.79±1.35a | |
| EC | 717.73±12.78b | 65.41±3.44a | 35.45±4.00ab | 14.33±1.43a | |
| ET | 821.97±10.71a | 68.47±4.28a | 36.49±2.31ab | 13.19±0.63a | |
| ECT | 842.16±14.57a | 59.12±11.37a | 30.64±1.92b | 10.75±1.64a | |
| P-value | PT | 0.00 | 0.99 | 0.10 | 0.08 |
| PCO2 | 0.13 | 0.64 | 0.05 | 0.30 | |
| PT×CO2 | 0.01 | 0.35 | 0.82 | 0.47 |
新窗口打开|下载CSV
大气CO2浓度升高使小麦灌浆期叶片最大羧化速率(Vc.max)、最大电子传递速率(Jmax)和Jmax/ Vc.max无显著变化。升温使灌浆期小麦叶片最大羧化速率(Vc.max)和最大电子传递速率(Jmax)分别降低39.5%和49.3%,使Jmax/Vc.max增加19.8%。CO2浓度和温度升高对叶片最大羧化速率(Vc.max)、最大电子传递速率(Jmax)和Jmax/Vc.max的交互作用不显著(表5)。
Table 5
表5
表5CO2浓度和气温升高对小麦灌浆期叶片最大羧化速率和最大电子传递速率的影响
Table 5
| 处理 Treatment | 最大羧化速率 Maximum carboxylation rate (Vc.max) (μmol·m-2·s-1) | 最大电子传递速率 Maximum electron transport rate (Jmax) (μmol·m-2·s -1) | Jmax/Vc.max | |||||
|---|---|---|---|---|---|---|---|---|
| CK | 102.50±6.05a | 103.78±9.45a | 0.99±0.04b | |||||
| EC | 102.63±8.49a | 104.00±5.60a | 0.98±0.03b | |||||
| ET | 71.47±6.15b | 60.43±6.58b | 1.19±0.03a | |||||
| ECT | 52.52±4.70b | 44.83±3.67b | 1.17±0.03a | |||||
| P-value | PT | 0.00 | 0.00 | 0.00 | ||||
| PCO2 | 0.18 | 0.28 | 0.65 | |||||
| PT×CO2 | 0.18 | 0.26 | 0.90 | |||||
新窗口打开|下载CSV
2.5 大气CO2浓度和气温升高对小麦叶片细胞色素含量的影响
大气CO2浓度升高使小麦灌浆期叶片叶绿素a、叶绿素b和叶绿素a+b分别显著下降13.4%、27.1%和17.0%,对类胡萝卜素含量无显著影响。升温使灌浆期小麦叶片叶绿素b显著增加54.9%,使类胡萝卜素含量显著下降22.6%,但对叶绿素a和叶绿素a+b无显著影响。CO2浓度和温度升高对叶片叶绿素a、叶绿素b、叶绿素a+b和类胡萝卜素含量的交互作用均显著,CO2浓度和气温升高对叶绿素a、叶绿素b和叶绿素a+b的交互作用大于CO2浓度和升温的单独效应之和,气温升高会增加灌浆期CO2浓度升高对小麦叶片叶绿素含量的负效应。CO2浓度和气温升高对类胡萝卜素含量的交互作用小于CO2浓度和升温的单独效应之和,CO2浓度升高会减弱灌浆期气温升高对小麦叶片类胡萝卜素含量的负效应(表6)。Table 6
表6
表6CO2浓度和气温升高对小麦灌浆期叶片细胞色素的影响
Table 6
| 处理 Treatment | 叶绿素a Chl a (mg·g-1 FW) | 叶绿素b Chl b (mg·g-1 FW) | 叶绿素a+b Chl a and Chl b (mg·g-1 FW) | 类胡萝卜素 Chl x∙c (mg·g-1 FW) | |||||
|---|---|---|---|---|---|---|---|---|---|
| CK | 2.44±0.14b | 0.27±0.04b | 2.71±0.18b | 8.94±1.46a | |||||
| EC | 2.51±0.27bc | 0.34±0.11b | 2.85±0.38b | 7.43±1.42a | |||||
| ET | 3.01±0.11a | 0.63±0.00a | 3.63±0.06a | 4.78±2.75b | |||||
| ECT | 2.20±0.03c | 0.32±0.04b | 2.42±0.21c | 7.89±0.71a | |||||
| P-value | PT | 0.28 | 0.00 | 0.08 | 0.02 | ||||
| PCO2 | 0.01 | 0.02 | 0.01 | 0.17 | |||||
| PT×CO2 | 0.00 | 0.00 | 0.00 | 0.05 | |||||
新窗口打开|下载CSV
2.6 大气CO2浓度和气温升高对小麦叶片糖、氮代谢物的影响
大气CO2浓度升高提高小麦灌浆期叶片可溶性糖和淀粉含量,增幅分别为92.9%和100.8%,但使硝酸还原酶活性下降38.5%,对纤维素和可溶性蛋白含量无显著影响。升温使灌浆期小麦叶片纤维素含量、可溶性蛋白含量和硝酸还原酶活性分别下降36.0%、47.9%和34.6%,对可溶性糖和淀粉含量无显著影响。CO2浓度和温度升高对叶片可溶性糖、可溶性蛋白含量和硝酸还原酶活性的交互作用显著,CO2浓度和气温升高对小麦叶片可溶性糖的交互作用小于CO2浓度和升温的单独效应之和,气温升高会减弱灌浆期CO2浓度升高对小麦叶片可溶性糖的正效应。CO2浓度和气温升高对小麦叶片硝酸还原酶的交互作用大于CO2浓度和升温的单独效应之和,CO2浓度升高会减弱灌浆期气温升高对小麦叶片硝酸还原酶活性的负效应(表7)。Table 7
表7
表7CO2浓度和气温升高对小麦灌浆期叶片碳氮代谢物的影响
Table 7
| 处理 Treatment | 可溶性总糖含量 Soluble sugar content (mg·g-1 FW) | 淀粉含量 Starch content (mg·g-1 FW) | 纤维素含量 Cellulose content (mg·g-1 FW) | 可溶性蛋白 Soluble protein (mg·g-1 FW) | 硝酸还原酶活性 Nitrate reductase activity (μg NO2-·g-1 FW·h-1) | |
|---|---|---|---|---|---|---|
| CK | 2.38±0.40d | 0.13±0.02b | 4.39±1.23a | 15.47±2.83b | 4.14±0.74a | |
| EC | 7.01±0.59a | 0.23±0.09a | 4.91±0.90a | 21.15±1.18a | 1.70±0.02b | |
| ET | 4.36±0.30c | 0.13±0.01b | 2.79±0.13b | 10.80±1.63c | 1.84±0.24b | |
| ECT | 5.99±0.72b | 0.28±0.03a | 3.16±0.99b | 8.25±1.15d | 1.98±0.34b | |
| P-value | PT | 0.15 | 0.47 | 0.02 | 0.00 | 0.00 |
| PCO2 | 0.00 | 0.00 | 0.51 | 0.22 | 0.00 | |
| PT×CO2 | 0.00 | 0.41 | 0.91 | 0.01 | 0.00 |
新窗口打开|下载CSV
2.7 大气CO2浓度和气温升高对小麦形态指标的影响
大气CO2浓度升高使小麦株高增加6.0%,对节数、穗长和茎粗无显著影响。升温使小麦穗长、茎粗和节数分别下降6.5%、5.2%和3.9%,对小麦株高无显著影响。CO2浓度和温度升高对小麦节数的交互作用显著,CO2浓度和气温升高的交互作用小于CO2浓度和升温的单独效应之和,CO2浓度升高会减弱气温升高对小麦节数的负效应(表8)。Table 8
表8
表8CO2浓度和气温升高对小麦形态指标的影响
Table 8
| 年份 Year | 处理 Treatment | 株高 Plant height (cm) | 穗长 Spike length (cm) | 茎粗 Stem-diameter (mm) | 节数 Internode number |
|---|---|---|---|---|---|
| 2017-2018 | CK | 63.34±0.90ab | 7.21±0.16a | 3.419±0.072a | 4.8±0.1a |
| EC | 65.76±2.30a | 7.64±0.26a | 3.458±0.066a | 4.9±0.0a | |
| ET | 60.41±0.29b | 7.3±0.09a | 3.418±0.059a | 4.8±0.1a | |
| ECT | 62.83±1.55ab | 7.52±0.13a | 3.574±0.026a | 4.7±0.1a | |
| 2019-2020 | CK | 66.19±2.32bc | 9.81±0.20a | 3.468±0.166a | 5.9±0.1a |
| EC | 71.26±1.16a | 9.08±0.24b | 3.520±0.102a | 5.9±0.1a | |
| ET | 65.03±0.91c | 8.49±0.20bc | 3.096±0.114b | 5.1±0.1b | |
| ECT | 70.53±1.02ab | 8.25±0.18c | 3.058±0.077b | 5.1±0.1b | |
| P-value | Pyear | 0.00 | 0.00 | 0.01 | 0.001 |
| PT | 0.07 | 0.00 | 0.01 | 0.01 | |
| PCO2 | 0.00 | 0.55 | 0.42 | 0.00 | |
| PT×CO2 | 0.91 | 0.61 | 0.91 | 0.01 |
新窗口打开|下载CSV
2.8 大气CO2浓度和气温升高对小麦生物量、产量及产量构成的影响
大气CO2浓度升高使小麦单位面积生物量和单位面积产量分别显著增加24.4%和26.1%。升温使小麦单位面积生物量和单位面积产量分别显著下降19.7%和23.0%。尽管CO2浓度和温度升高对小麦生物量和产量的交互作用不显著,但正常气温条件下大气CO2浓度升高对小麦生物量(18.3%)和产量(21.3%)的增幅小于气温升高条件下CO2浓度升高对小麦生物量(32.6%)和产量(32.9%)的增幅。表明CO2浓度升高可以一定程度上减缓升温对小麦产量和生物量的负面影响。大气CO2浓度升高对小麦收获指数无显著影响,气温升高使小麦收获指数减少3.9%,CO2浓度和温度升高对小麦收获指数的交互作用不显著(图2)。图2
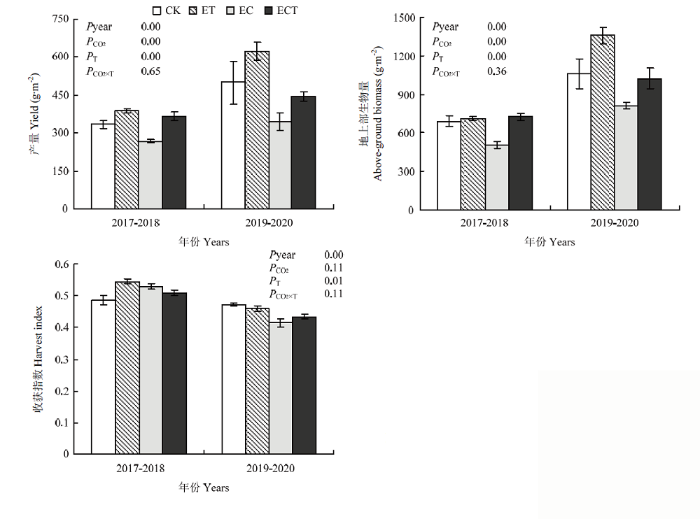 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2CO2浓度和气温升高对小麦生物量、产量和收获指数的影响
Fig. 2Effects of elevated CO2 and increased temperature on the biomass, yield and harvest index of wheat
大气CO2浓度升高使小麦单位面积穗数显著增加23.3%,对穗粒数和千粒重无显著影响。升温使小麦穗粒数和千粒重分别显著下降12.4%和6.9%,对小麦单位面积穗数无显著影响。CO2浓度升高对小麦产量的增加主要靠提高小麦单位面积穗数,而气温升高对小麦产量的负面影响主要是由于其减小了小麦穗粒数和千粒重。CO2浓度和温度升高对小麦穗粒数、单位面积穗数和千粒重的交互作用均不显著(表9)。
Table 9
表9
表9CO2浓度和气温升高对小麦产量构成的影响
Table 9
| 年份 Year | 处理 Treatment | 单穗粒数 Grain number per spike | 单位面积穗数 Spikes number (No./m2) | 千粒重 1000-seed weight (g) |
|---|---|---|---|---|
| 2017-2018 | CK | 22.7±1.7ab | 327.1±9.8ab | 45.44±1.84a |
| EC | 25.3±1.2a | 349.0±23.2a | 44.45±0.66a | |
| ET | 21.1±1.0b | 292.7±16.4b | 43.68±0.74a | |
| ECT | 23.9±0.3ab | 344.8±7.3a | 44.55±0.66a | |
| 2019-2020 | CK | 45.0±1.4a | 230.0±21.9c | 43.53±1.12a |
| EC | 46.3±6.7a | 371.5±23.4a | 38.39±4.00a | |
| ET | 37.2±4.4a | 254.8±11.6bc | 37.00±1.55a | |
| ECT | 39.8±2.1a | 297.2±12.9b | 36.32±0.64a | |
| P-value | Pyear | 0.00 | 0.00 | 0.00 |
| PT | 0.03 | 0.08 | 0.05 | |
| PCO2 | 0.21 | 0.00 | 0.23 | |
| PT×CO2 | 0.83 | 0.16 | 0.21 |
新窗口打开|下载CSV
3 讨论
前人研究认为,大气温度升高条件下作物冠层温度升高,叶绿素合成受阻,光合作用受抑制,生育期缩短,作物产量形成受到抑制[4,22-23]。本研究基本支持上述结论,气温持续升高2℃条件下,小麦拔节期与开花期净光合速率没有显著变化,但叶片叶绿素含量降低,且花后到成熟时间缩短4 d,灌浆期最大羧化速率(Vc.max)和最大电子传递速率(Jmax)也下调,硝酸还原酶活性、叶片可溶性蛋白与纤维素含量显著下降,进而使小麦地上部生物量与穗粒数、千粒重同时明显降低,产量形成受到限制。经典理论认为,C3植物叶片光合作用的最佳温度与Rubisco在羧化反应和加氧反应之间的分配比例存在较大关系。增温下加氧反应增强,光合作用的最佳温度降低,羧化反应易受抑制;CO2升高下羧化反应增强,光合作用的最佳温度升高[4,24],因此二因素的交互作用有利于弥补单纯升温带来的危害[24]。
本研究中,灌浆期小麦叶片最大羧化速率和最大电子传递速率在升温下被显著抑制,在CO2浓度升高下得到缓解,但最大光合能力仍然没有得到缓解。这与叶绿素a、叶绿素b和叶绿素a+b浓度显著下降、光补偿点升高、硝酸还原酶活性下降有关,此时光合适应出现,光合能力下降。所以,推测光合色素浓度下降[25]、光补偿点增加、硝酸还原酶活性下降低或是造成后期(灌浆期)CO2肥效减弱的原因。尽管如此,大气CO2浓度升高下,小麦叶片净光合速率在拔节期和孕穗期得到提高,且叶片可溶性糖和淀粉含量在灌浆期仍然维持较高,表明前期净光合速率的增加仍可对灌浆期碳水化合物的积累起到积极作用,有助于为后期产量形成提供物质基础[26]。同时,CO2浓度升高在一定程度上弥补了升温对花后到成熟时间缩短的负效应,灌浆时间对产量形成的限制作用减弱。所以,CO2浓度升高可以通过提高前期光合速率,增加碳水化合物积累,并延长灌浆时间,提高小麦产量,补偿升温的负面影响。
在我国南方,CO2浓度升高增加小麦产量,升温减少小麦产量,交互作用下小麦产量略有下降[18],但在北方(黑龙江省)交互作用提高了大豆和玉米的产量[27],CO2浓度升高可以部分补偿升温对北方小麦(河北)产量的负面影响[11]。本研究表明,增温通过减少穗粒数与千粒重使产量降低。而CO2浓度升高可通过增加单位面积穗数使小麦产量增加,前期叶片净光合速率较高与有机物积累增加或是促使其分蘖易成穗的原因之一。但在交互作用下,小麦产量、穗粒数、千粒重、单位面积穗数均较对照无明显变化。另外,本研究是在小麦生长季无极端天气事件(高温、干热风等)的情况下进行,未来气候变化条件下,极端天气事件发生概率将增加,高温热害、干热风危害情况下,CO2浓度升高是否可以抵消升温的负面影响有待深入研究。
4 结论
气温持续升高2℃会缩短小麦全生育期及开花到成熟时间,使拔节期小麦叶片净光合速率下降,而对灌浆期光合作用无显著影响。升温使灌浆期小麦叶片纤维素含量、可溶性蛋白含量和硝酸还原酶活性下降,使小麦穗粒数和千粒重下降,产量、生物量和收获指数下降;CO2浓度升高200 μmol·mol-1提高了小麦光合能力,增加灌浆期叶片碳水化合物含量,虽然生长后期出现光适应,但仍可通过增加单位面积穗数增加小麦产量;在增温条件下,CO2浓度升高可通过增加开花到成熟时间、提高小麦净光合速率、增加光合代谢物等弥补升温对小麦生物量和产量的负效应。本研究未考虑极端天气事件(高温、干热风等)对小麦生产的影响,高温热害、干热风危害情况下,CO2浓度升高与气温升高对小麦生产的影响有待深入研究。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/gcb.1995.1.issue-1URL [本文引用: 7]
DOI:10.1016/j.envexpbot.2020.104154URL [本文引用: 1]
[本文引用: 1]
DOI:10.1016/S2095-3119(16)61428-4URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.agee.2015.04.007URL [本文引用: 1]
[本文引用: 3]
DOI:10.1038/nclimate2470URL [本文引用: 2]
[本文引用: 1]
DOI:10.1111/gcb.2010.17.issue-2URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1038/s41477-018-0263-1URL [本文引用: 1]
DOI:10.1111/gcb.13065URL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.3773/j.issn.1005-264x.2010.06.012 [本文引用: 2]

光合作用对光和CO2响应模型是研究植物生理和植物生态学的重要工具, 可为植物光合特性对主要环境因子的响应提供科学依据。该文综述了当前光合作用对光和CO2响应模型的研究进展和存在的问题, 并在此基础上探讨了这些模型的可能发展趋势。光合作用涉及光能的吸收、能量转换、电子传递、ATP合成、CO2固定等一系列复杂的物理和化学反应过程。光合作用由原初反应、同化力形成和碳同化3个基本过程构成, 任一个过程均可对光合作用速率产生直接的影响。光合作用对光响应模型只涉及光能的转换, 而光合作用的生化模型包含了同化力形成和碳同化这两个基本过程。把光合作用的原初反应, 即把参与光能吸收、传递和转换的捕光色素分子的物理参数(如捕光色素分子数、捕光色素分子光能吸收截面、捕光色素分子处于激发态的平均寿命等)结合到生化模型中, 可能是今后光合作用对光响应机理模型的发展方向。
DOI:10.3773/j.issn.1005-264x.2010.06.012 [本文引用: 2]
光合作用对光和CO2响应模型是研究植物生理和植物生态学的重要工具, 可为植物光合特性对主要环境因子的响应提供科学依据。该文综述了当前光合作用对光和CO2响应模型的研究进展和存在的问题, 并在此基础上探讨了这些模型的可能发展趋势。光合作用涉及光能的吸收、能量转换、电子传递、ATP合成、CO2固定等一系列复杂的物理和化学反应过程。光合作用由原初反应、同化力形成和碳同化3个基本过程构成, 任一个过程均可对光合作用速率产生直接的影响。光合作用对光响应模型只涉及光能的转换, 而光合作用的生化模型包含了同化力形成和碳同化这两个基本过程。把光合作用的原初反应, 即把参与光能吸收、传递和转换的捕光色素分子的物理参数(如捕光色素分子数、捕光色素分子光能吸收截面、捕光色素分子处于激发态的平均寿命等)结合到生化模型中, 可能是今后光合作用对光响应机理模型的发展方向。
DOI:10.1007/BF00386231URL [本文引用: 1]
DOI:10.1590/S0103-84782005000300041URL [本文引用: 1]
DOI:10.1104/pp.109.137265URL [本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.scitotenv.2019.02.149URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}