 ,, 刘志, 李振洲, 付江鹏, 慕瑞瑞, 康建宏宁夏大学农学院,银川 750021
,, 刘志, 李振洲, 付江鹏, 慕瑞瑞, 康建宏宁夏大学农学院,银川 750021Photosynthetic Response Characteristics of Maize Under Drip Irrigation Based on Machine Learning
LIU HuiFang, HE Zheng, JIA Biao,, LIU Zhi, LI ZhenZhou, FU JiangPeng, MU RuiRui, KANG JianHongSchool of Agriculture, Ningxia University, Yinchuan 750021通讯作者:
责任编辑: 杨鑫浩
收稿日期:2019-04-2接受日期:2019-07-3网络出版日期:2019-09-01
| 基金资助: |
Received:2019-04-2Accepted:2019-07-3Online:2019-09-01
作者简介 About authors
刘慧芳,E-mail:18747555998@163.com。

摘要
关键词:
Abstract
Keywords:
PDF (615KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
刘慧芳, 贺正, 贾彪, 刘志, 李振洲, 付江鹏, 慕瑞瑞, 康建宏. 基于机器学习的滴灌玉米光合响应特征[J]. 中国农业科学, 2019, 52(17): 2939-2950 doi:10.3864/j.issn.0578-1752.2019.17.003
LIU HuiFang, HE Zheng, JIA Biao, LIU Zhi, LI ZhenZhou, FU JiangPeng, MU RuiRui, KANG JianHong.
0 引言
【研究意义】光合作用是作物光合生产力大小的重要生理生态过程[1,2,3,4],可反映作物在不同生长条件下的生理生态状况[5,6]。而净光合速率是反应作物光合能力的主要指标[7],光响应曲线则是研究作物净光合速率随着光合有效辐射变化的模型,通过光响应曲线模拟可以得到一些不同生理过程及生态环境变化的重要参数[8],【前人研究表明】大量研究指出直角双曲线修正模型是拟合各种不同处理条件下光响应动态变化规律的最优模型,而如何精准计算光合参数和提高模型精度是模型应用的难点[9,10,11,12]。近年来,随着人工智能技术发展,机器学习方法越来越多地应用于作物建模。基于机器学习方法可进行作物钾素等营养诊断[13]、SPAD值的含量监测[14,15],其预测精度均高于传统回归方法。曾继业等[16]选用机器学习的近似贝叶斯法来确定Farquhar光合模型的生理参数,发现模拟值和实测值的线性回归曲线斜率1.04与理论1.0基本保持一致。钾素作为影响玉米光合代谢变化的重要因子,直接参与光合磷酸化等光合产物的运输过程[17]。研究表明,作物缺钾会明显影响叶绿素的合成,引起作物叶脉叶尖黄化和卷曲,加速叶片衰老。低钾胁迫会破坏叶片光合器官,使作物净光合速率下降,叶片气孔关闭,RuBPcase活性下降,籽粒明显减少。同时,使PSⅡ反应中心受损,导致Fv/Fm、ΦPSⅡ、qP下降。适量施钾肥可提高叶片光合速率和蒸腾效率,增强叶片气孔调节能力,还能提高叶片光合酶活性和氮代谢酶活性,加速作物对氮素的吸收和同化作用[18,19,20,21]。【本研究切入点】前人研究丰富了基于机器学习的作物预测模型建立方法,为作物光响应曲线模型精度的提高提供新思路,但近似贝叶斯法在数据拟合时易出现过拟合问题,而机器学习网格搜索法将估计函数的参数通过交叉验证的方法进行优化,可避免近似贝叶斯法的缺点[16]。目前,国内外关于钾肥在玉米叶绿素、荧光、光合等方面研究相对较多[22,23,24],而钾肥对玉米光响应曲线的关系研究较少。【拟解决的关键问题】本文从不同钾肥水平下研究吐丝期玉米叶片光响应曲线特征,选用直角双曲线修正模型,采用网格搜索的机器学习法来计算最优参数,提高模型精度,为研究宁夏干旱半干旱地区滴灌玉米光合生理机制及光合响应特征提供新思路。1 材料与方法
1.1 试验地概况
试验于2017—2018年4—9月在宁夏农垦平吉堡农场(106 °01′44″E,38°26′39″N)进行,田间土壤类型为壤土,肥力中等,前茬作物为玉米。试验田0—20 cm耕层土壤基础肥力如表1所示。2006—2016年降雨量和日平均气温分别为272.6 mm和8.6℃,2017年降雨量和日平均气温分别为501.9 mm和13℃,2018年总降雨量和日平均气温分别为875.5 mm和14℃,其中2017年和2018年玉米全生育期降雨量及日平均气温变化如图1所示。Table 1
表1
表1试验地土壤基础肥力
Table 1
| 试验地 Test location | 年份 Year | pH | 有机质 OM (g·kg-1) | 全氮 Total N (g·kg-1) | 全磷 Total P (g·kg-1) | 碱解氮 Available N (mg·kg-1) | 速效磷 Available P (mg·kg-1) | 速效钾 Available K (mg·kg-1) |
|---|---|---|---|---|---|---|---|---|
| 平吉堡农场 Pingjipu farm | 2017 | 7.92 | 11.51 | 0.82 | 0.59 | 37.42 | 19.11 | 102.48 |
| 2018 | 7.57 | 12.78 | 0.70 | 0.45 | 36.64 | 17.42 | 95.34 |
新窗口打开|下载CSV
图1
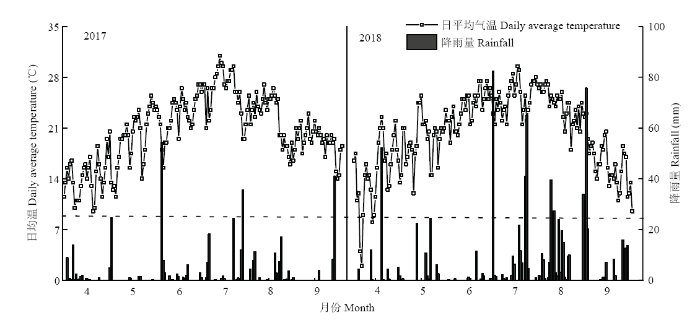 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1玉米不同生育期气象条件
Fig. 1Meteorological conditions of different growth stages of maize
1.2 试验设计
试验采用单因素随机区组设计,设置6个施钾水平,分别为K0(0,CK)、K1(90 kg·hm-2)、K2(180 kg·hm-2)、K3(270 kg·hm-2)、K4(360 kg·hm-2)、K5(450 kg·hm-2),3次重复,共18个小区,小区面积为67.5 m2(长15 m×宽4.4 m),采用宁夏大力推广的机械播种模式,宽窄行种植,宽行70 cm,窄行40 cm,株距20 cm,密度为9×104株/hm2。供试品种为天赐19(TC19),生育期为137 d左右,株型为半紧凑,属中晚熟型杂交品种。供试氮肥为尿素(N﹕P﹕K=46﹕0﹕0),磷肥为磷酸一铵(N﹕P﹕K= 12﹕62﹕0),钾肥为硫酸钾(N﹕P﹕K=0﹕0﹕52),均为水溶性肥料,其中磷肥(138 kg·hm-2)、氮肥(300 kg·hm-2)为常规用量,肥料用量均以元素态计算。采用水肥一体化滴灌模式随水施入,遵循“随水施肥,少量多次”的原则[25,26,27],结合宁夏当地滴灌玉米推荐施肥模式,全生育期共施肥8次,分别为苗期1次、拔节期3次、抽雄期1次、灌浆期3次。各生育时期施肥量分别占总量的10%、45%、20%、25%。其他管理措施与当地大田生产相同。灌水方式为滴灌灌溉。玉米于4月底播种,9月底收获。
1.3 测试项目与方法
在玉米吐丝期,应用Li-6400XT(Li-Cor,Lincoln,USA)型便携式光合作用测定系统进行光合响应曲线测定。选择晴朗天气上午的9:00-11:30(在自然光诱导1 h后),每个处理随机选取3 株长势一致、受光方向一致无破损的穗位叶植株进行测定,测定玉米叶片在每一光强下的净光合速率(Pn),蒸腾速率(Tr),气孔导度(Gs),胞间CO2浓度(Ci)等光合参数。测定时采用 Li6400-02B 红蓝人工光源测量叶室,通过开放式气路,设定温度为 25℃,CO2 浓度为400 μmol·mol-1(大气CO2浓度),空气相对湿度为50%—70%,光合有效辐设(PAR)梯度为2 000、1 700、1 400、1 100、800、600、400、200、110、80、50、20、0 μmol·m-2·s-1,仪器自动记录数据,仪器最小等待时间和最大等待时间分别为120 s和180 s。每个叶片重复测定3次,取平均值进行分析。通过直角双曲线修正模型确定最大净光合速率(Pnmax)、光饱和点(LSP)、光补偿点(LCP)、暗呼吸速率(Rd)和表观量子效率(AQE)。根据测得数据计算水分利用效率(WUE)和呼吸效率(RE)。计算公式如下:1.4 作物光合响应模型
本文采用直角双曲线修正模型对不同钾肥水平下玉米光响应曲线进行拟合分析,并用SPSS非线性回归法和机器学习网格搜索法对各处理玉米光合参数进行计算,从而提高模型精度。模型如下:式中,Pn是净光合速率,μmol·m-2·s-1;α是表观量子效率;I是光量子通量密度,μmol·m-2·s-1;Pnmax是最大净光合速率,μmol·m-2·s-1;Rd是植物的暗呼吸速率,μmol·m-2·s-1;β是修正系数;γ是一个与光强无关的系数。
1.5 机器学习网格搜索法对修正模型的拟合
通过寻找最优的超参值组合,进一步提高模型的性能。本文使用机器学习网格搜索法来实现直角双曲线修正模型精度的优化,通过指定不同的超参列表进行穷举搜索,并评估每个组合对模型性能的影响,获得参数的最优组合。本研究将玉米光响应曲线参数可能的取值进行排列组合,列出所有可能的组合结果生成“网格”。尝试拟合函数所有参数组合后,返回1个合适的分类器,自动调整至最佳参数组合,从而得到最优参数(图2)。图2
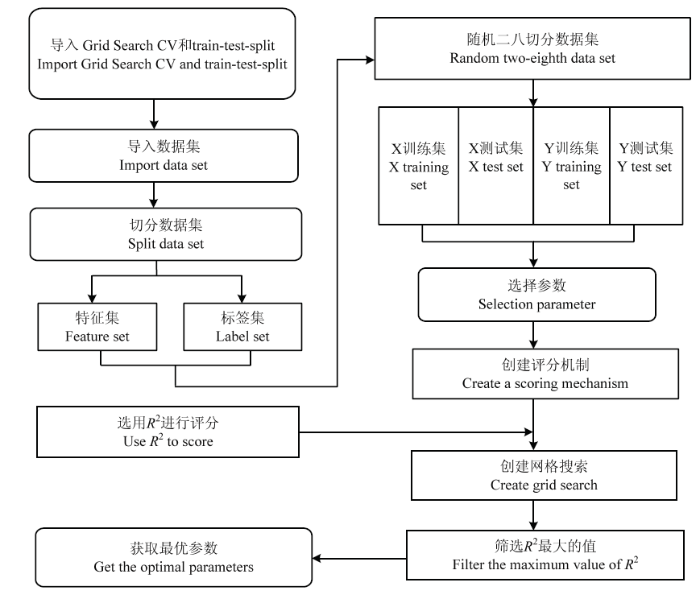 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2机器学习网格搜索法流程图
Fig. 2Machine learning grid search method flow chart
1.6 数据处理与模型检验
本文选取决定系数(R2)、均方根误差(RMSE)及平均绝对误差(MAE)对模型精度进行评价,其中RMSE和MAE越小、R2越接近于1,模型的精度越高。模型检验表达式如下:式中,Oi是观测值,Si是模拟值,$\bar{O}$是观测值的平均值,n是样本数。
2 结果
2.1 不同钾肥处理下玉米吐丝期光响应曲线动态变化特征
由图3可知,2017—2018年玉米吐丝期Pn随钾肥施用量增加的变化规律近乎一致。2年的数据表明,不同钾肥施用条件下,玉米在PAR<1 000 μmol·m-2·s-1时,光合有效辐射(Pn)随PAR的增加呈直线迅速增加;当PAR>1 000 μmol·m-2·s-1,光合有效辐射(Pn)随PAR的增加而迅速减缓,最后趋于稳定。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同施钾量下净光合速率对光合有效辐射的响应
Fig. 3Net photosynthetic rate-light response curves of maize under different potassium application rates
在玉米吐丝期,钾肥施用量小于360 kg·hm-2(K4)时,随着钾肥施用量和光合有效辐射的增加,玉米的净光合速率逐渐升高,当施肥量达到450 kg·hm-2(K5)时,玉米的净光合速率反而低于360 kg·hm-2(K4)处理。低肥(K1)处理和CK条件下,PAR>1 500 μmol·m-2·s-1时就达到光饱和,出现光抑制现象。由此可见,在一定的施肥范围内,适量的施钾肥有利于提高玉米叶片对光合有效辐射的响应,进而提高玉米叶片的光利用能力。
2.2 施钾对滴灌玉米吐丝期光合生理特性的影响
由图4-(a)、(b)可知,2年间Gs随钾肥施用量和光照强度增加的变化规律基本保持一致。2年不同施肥量下,玉米吐丝期的气孔导度Gs随PAR增加从0.012增至0.328 mol·m-2·s-1,当PAR增加至1 500 μmol·m-2·s-1时,Gs趋于平缓并有下降趋势。当施肥量小于360 kg·hm-2(K4)时,随着钾肥施用量和光合有效辐射的增加,Gs逐渐升高,当施肥量达到450 kg·hm-2(K5)时,Gs反而低于360 kg·hm-2(K4)处理,这与Pn的研究结果一致。不同处理的Gs和光合速率呈正相关,随着PAR的升高,净光合速率越大,气孔开放程度越大,从而增大叶片对CO2的吸收,有利于提高玉米的光合效率。在PAR为1 500 μmol·m-2·s-1时,K4处理的气孔导度开放程度开始下降,与Pn的变化规律相符,同样说明适量的施肥有利于提高玉米光合速率,过量的施肥会出现光抑制现象。图4
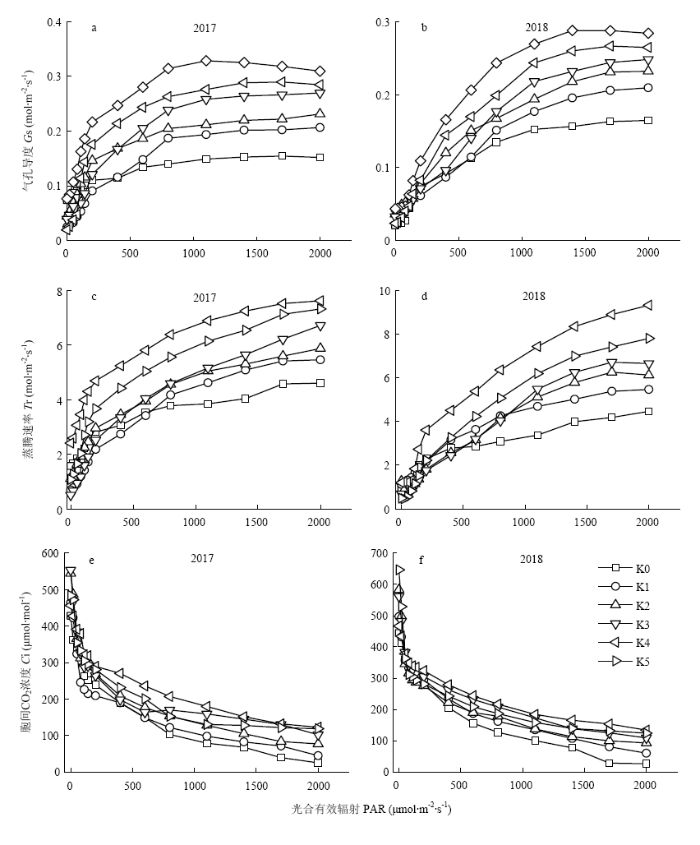 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同施钾量下玉米光合参数的比较
Fig. 4Comparison of photosynthetic parameters of maize under different potassium application rates
由图4-(c)、(d)可知,Tr的变化趋势与Gs几乎保持一致。2年间Tr随着施肥量和光照强度的增加呈先快速升高后逐渐趋于平缓趋势。当PAR≤200 μmol·m-2·s-1时,各处理差异不明显,随PAR的升高,K2、K3、K4、K5 4个处理Tr曲线上升幅度较高,但K0、K1 2个处理表现相反,由于施肥量过低,随PAR的升高,为了保证正常生长,导致气孔开放程度减小,从而使蒸腾速率降低。
由图4-(e)、(f)可知,2年间各处理玉米Ci均在PAR<200 μmol·m-2·s-1时,Ci下降幅度极大,在PAR为 200—1 500 μmol·m-2·s-1时,Ci下降幅度减缓。这是由于在光强增大的初级阶段,玉米叶片净光合速率迅速增加,需要足够多的CO2来做原料,从而进行光合作用。在PAR大于1 600 μmol·m-2·s-1时,光合作用增幅减缓,Ci亦趋于平缓。2年间各处理下降趋势为K0>K1>K2>K3>K5>K4,说明施肥不足或者过量施肥引起气孔阻力增大,光合速率降低,从而导致Ci降低。
2.3 玉米吐丝期水分利用效率与呼吸效率对钾肥的响应
由图5-(a)、(b)可知,2年的玉米叶片WUE在不同施肥条件下有明显差异,变化规律基本相似。2年间各处理在PAR<200 μmol·m-2·s-1时,叶片WUE上升幅度极大,在PAR为200—1 500 μmol·m-2·s-1时,叶片WUE上升幅度减缓,当PAR大于1 600 μmol·m-2·s-1时,叶片WUE亦趋于平缓。低钾胁迫下(CK、K1)玉米叶片WUE较高,适量的钾肥(K4)和高钾(K5)处理下,结果相反。这可能与钾是控制叶片气孔导度开闭的重要因素有关,低钾胁迫下,植物叶片气孔导度开放程度较小,引起叶片光合速率降低,导致水分利用效率增大。图5
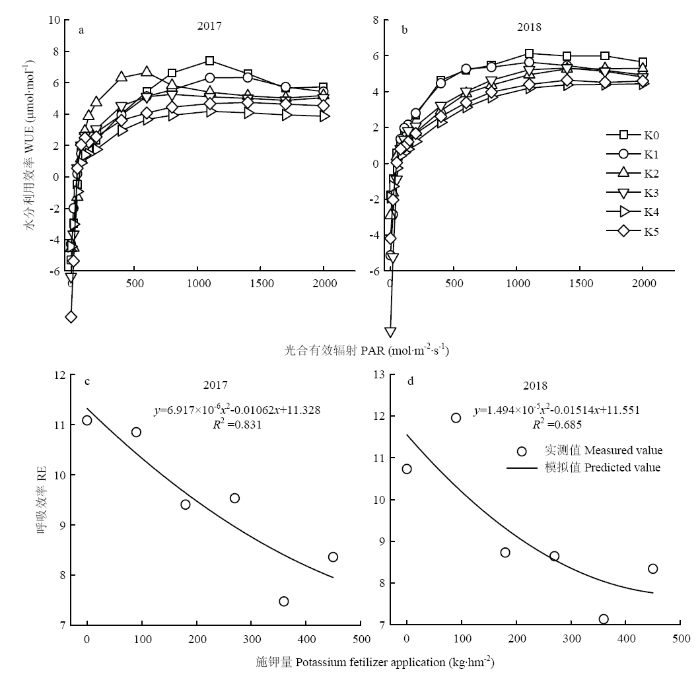 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5不同施钾量下玉米光合参数的计算
Fig. 5Calculation of photosynthetic parameters of maize under different potassium application rates
由图5-(c)、(d)可知,2年间玉米RE随钾肥施用量的增加变化明显,呈二次函数递减趋势,当施肥量小于360 kg·hm-2(K4)时,随着施钾量的增加,RE逐渐减小,说明玉米在低钾胁迫下能够调节自身生理机制,降低呼吸消耗从而维持正常的生理功能。当施钾量为450 kg·hm-2(K5)时,RE反而增加。说明过量的施肥量,导致植物营养生长过旺,会加速能量的消耗。
2.4 网格搜索法和非线性回归分析法对修正模型拟合度比较
运用基于机器学习的网格搜索法和非线性回归分析对直角双曲线修正模型进行拟合。采用RMSE、MAE和R2误差计算的大小来判断模拟值和观测值之间的差异,RMSE、MAE越小、R2越接近于1,说明模型模拟精度越高,反之,则模拟精度越差。模型评价参数如表2所示,2种方法均能较好拟合各处理玉米光响应动态变化规律,R2均大于0.990,RMSE均未超过3.250,MAE均未超过2.410,且RMSE和MAE均随着施钾量的增加而逐渐减小。说明机器学习方法和传统非线性回归分析法对直角双曲线修正模型拟合效果相当,是模型拟合的较好方法。但机器学习网格搜索法在不施钾肥(K0)和低钾(K1)条件下模拟效果更加突出,其R2均大于0.991,RMSE均小于1.487、MAE均小于1.350,其他处理下(K2—K5)机器学习模拟效果与传统方法的模拟效果接近,R2均大于0.993,RMSE均小于0.952、MAE均小于0.860,表明机器学习网格搜索法是提高直角双曲线修正模型精度的重要方法。Table 2
表2
表22种计算分析方法对玉米光响应曲线的模拟精度
Table 2
| 年份 Year | 方法 Method | 处理 Treatment | RMSE | MAE | R2 |
|---|---|---|---|---|---|
| 2017 | 非线性回归分析法 Nonlinear regression analysis | K0 | 2.030 | 1.869 | 0.990 |
| K1 | 1.319 | 1.079 | 0.992 | ||
| K2 | 1.147 | 0.899 | 0.993 | ||
| K3 | 0.828 | 0.656 | 0.995 | ||
| K4 | 0.610 | 0.498 | 0.994 | ||
| K5 | 0.446 | 0.379 | 0.995 | ||
| 网格搜索法 Grid search method | K0 | 1.335 | 1.096 | 0.993 | |
| K1 | 1.312 | 1.092 | 0.993 | ||
| K2 | 0.952 | 0.622 | 0.994 | ||
| K3 | 0.708 | 0.617 | 0.995 | ||
| K4 | 0.546 | 0.504 | 0.996 | ||
| K5 | 0.662 | 0.557 | 0.995 | ||
| 2018 | 非线性回归分析法 Nonlinear regression analysis | K0 | 1.827 | 1.445 | 0.987 |
| K1 | 1.731 | 1.323 | 0.991 | ||
| K2 | 0.872 | 0.733 | 0.993 | ||
| K3 | 0.858 | 0.717 | 0.994 | ||
| K4 | 0.644 | 0.566 | 0.996 | ||
| K5 | 0.312 | 0.226 | 0.997 | ||
| 网格搜索法 Grid search method | K0 | 1.487 | 1.350 | 0.991 | |
| K1 | 0.975 | 0.860 | 0.993 | ||
| K2 | 0.839 | 0.716 | 0.993 | ||
| K3 | 0.774 | 0.620 | 0.995 | ||
| K4 | 0.750 | 0.637 | 0.996 | ||
| K5 | 0.330 | 0.315 | 0.996 |
新窗口打开|下载CSV
2.5 基于网格搜索的机器学习法对滴灌玉米吐丝期光响应曲线参数计算
由表3可知,2年间玉米吐丝期光响应曲线的各光合参数随施钾量增加的变化规律几乎保持一致,随施钾量的增加最大净光合速率(Pnmax)、光补偿点(LCP)、光饱和点(LSP)及暗呼吸速率(Rd)等光合生理特征参数均呈先增加后降低的趋势。其中Pnmax的变化范围为23.96—34.65 μmol·m-2·s-1和25.36—35.67 μmol·m-2·s-1。在玉米吐丝期,K4处理下的Pnmax值分别为34.65 μmol·m-2·s-1和35.67 μmol·m-2·s-1,比CK、K1、K2、K3、K5分别提高了40.6%—44.6%、13.8%—27.2%,8.7%—12.9%,3.6%—5%,1.1%—1.9%,K4处理下光饱和点达到最大,分别为 2 034.09 μmol·m-2·s-1和2 042.04 μmol·m-2·s-1,且表观量子效率高于其他处理,可知在施肥量为360 kg·hm-2(K4)时,玉米叶片光能转化率较高。当施肥量为450 kg·hm-2(K5)时,Ic和Is分别比K4降低20%—32%和9%—17%。可知,施用适量的钾肥能明显提高玉米对强光的适应性及光能利用率,过低和过高的施肥量都会导致光合速率下降,说明适量的钾素能加速对氮素的同化吸收和利用。Table 3
表3
表3不同施钾量玉米光响应曲线光合参数
Table 3
| 年份 Year | 处理 Treatment | 表观量子效率 Q (μmol·μmol-1) | 最大光合速率 Pnmax (μmol·m-2·s-1) | 光补偿点 Ic (μmol·m-2·s-1) | 光饱和点 Is (μmol·m-2·s-1) | 暗呼吸速率 Rd (μmol·m-2·s-1) | 决定系数 R2 |
|---|---|---|---|---|---|---|---|
| 2017 | K0 | 0.041 | 23.959 | 55.922 | 1453.189 | 2.161 | 0.987 |
| K1 | 0.046 | 27.239 | 59.061 | 1627.178 | 2.510 | 0.991 | |
| K2 | 0.050 | 30.672 | 68.087 | 1965.095 | 3.260 | 0.993 | |
| K3 | 0.053 | 32.953 | 73.295 | 2126.701 | 3.456 | 0.994 | |
| K4 | 0.061 | 34.653 | 76.505 | 2694.469 | 4.635 | 0.996 | |
| K5 | 0.056 | 33.997 | 63.495 | 2034.091 | 4.066 | 0.997 | |
| 2018 | K0 | 0.042 | 25.368 | 56.110 | 1504.405 | 2.364 | 0.99 |
| K1 | 0.043 | 31.341 | 61.278 | 1568.254 | 2.622 | 0.992 | |
| K2 | 0.047 | 32.807 | 63.761 | 1984.520 | 3.758 | 0.993 | |
| K3 | 0.056 | 34.407 | 67.300 | 2282.035 | 3.982 | 0.995 | |
| K4 | 0.063 | 35.678 | 78.092 | 2893.226 | 5.00 | 0.994 | |
| K5 | 0.058 | 35.256 | 71.462 | 2462.256 | 4.228 | 0.995 |
新窗口打开|下载CSV
3 讨论
光合作用是农作物生长发育和产量形成的基础,钾素能够直接参与农作物光合作用、电子传递等重要光合过程[28]。研究发现,低钾条件下,K+能明显减弱气孔导度,使气孔阻力增加,CO2供应受阻,蒸腾效率降低,阻碍作物正常生长;钾素能明显提高作物对氮素的吸收和利用,并很快转化为蛋白质[21-24,28-29]。本研究结果表明,在玉米吐丝期,叶片施钾肥量小于360 kg·hm-2(K4)时,Pn、Gs、Tr均随施肥量和光照强度的增加呈先迅速增加后趋于缓慢的趋势,而Ci相反(图3—4)。这是因为K+活性降低,在保卫细胞离子传输中,减少碳酸酐酶和水通道蛋白的通过,从而限制CO2通过气孔和叶肉细胞的扩散。其中在K0和K1条件下,光合参数增加趋势缓慢且不稳定,分析其原因是玉米受低钾胁迫时,引起玉米光合酶和氮代谢酶活性下降,导致玉米植株不能正常生长。当施肥量达到450 kg·hm-2(K5)时,Pn、Gs、Tr反而低于K4处理,这由于土壤溶液中的高K+浓度会抑制玉米对Mg的吸收,而Mg是玉米叶绿体中主要的中心原子,进而导致玉米叶片光合速率下降[30,31]。本研究结果发现,2018年宁夏平吉堡玉米各处理穗位叶光合参数增长趋势普遍大于2017年,这由于2018年降雨充足(图1),又恰逢玉米吐丝时期,玉米由营养生长转为生殖生长阶段,是玉米一生中生长最旺盛的阶段,导致光饱和点较高。在适宜的钾肥下,作物能有效利用水分,降低蒸腾作用。本研究表明,在玉米吐丝期,不施钾肥(K0)和低钾(K1)处理下的玉米叶片WUE较高,适量的钾肥(K4)和高钾(K5)处理下,结果恰好相反(图5)。这由于玉米受低钾胁迫时,玉米叶片气孔导度开放程度较小,叶片光合速率降低,导致水分利用效率增大,但在高PAR下水分利用效率仍保持较高水平,表明玉米在受低钾胁迫时,能够通过自身生理调节来适应养分胁迫,维持一定的光合作用和较好的水分利用效率,从而保障正常生长[32]。作物光合参数初始量子效率α可反映其对光能利用效率的大小,α值的高低与作物叶片光能转化效率呈正比[10,11],Pnmax是衡量作物叶片最大光合能力的指标[33,34],而Rd和LCP是作物在逆境条件下自身的保护机制,通过降低暗呼吸速率和光补偿点来获得最大碳积累,进而维持自身生长[35]。在适宜生长条件下,作物光合参数α一般在0.04—0.07 μmol·μmol-1[9,10],本研究结果表明,α随施钾量的增加呈现先增加后降低的趋势,2017年在0.041—0.061 μmol·μmol-1之间,2018年在0.042—0.063 μmol·μmol-1之间,在过量施钾(K5)条件下会产生光合抑制现象,与赵丽[36]等研究结果一致,说明在K4条件下,玉米叶片吸收的光能和转化的光能色素蛋白体较多(表3)。本研究发现,Pnmax、Rd、LSP和LCP均随着施钾量的增加呈先升高后降低的变化趋势,但降幅较小,在施肥量大于360 kg·hm-2(K4)时,开始下降(表3)。由此表明,适宜的施肥量有利于提高玉米叶片对弱光的利用能力,增强玉米叶片对PAR的适应性。
光响应曲线是精准反映作物光化学过程中的光化学效率高低的有效方法[4]。大量研究表明,直角双曲线修正模型是反映作物不同生长环境条件下光合特性的最优模型[11],但如何计算光合参数使模型达到最佳效果一直以来是科技工作者的难点[9,10]。许多研究采用传统的非线性回归方法拟合直角双曲线修正模型,但运算过程繁琐且模型精度不高。而基于机器学习的网格搜索法简捷、快速,且能解决这一难题。如曾继业等[16]选用近似贝叶斯法来获取Farquhar光合模型的生理参数,效果可观;依尔夏提·阿不来提等[15]通过随机森林法估算BP神经网络和PLSR模型精度,结果表明PLSR模型精度高于BP神经网络。本研究采用基于机器学习的网格搜索法和传统的非线性回归法2种方法拟合,结果表明2种方法均能较好的拟合各钾肥处理下玉米光响应动态变化规律(表2),模型的决定系数R2均大于0.990,由此可见,机器学习方法拟合效果与传统非线性回归分析法拟合效果相当,但机器学习网格搜索法在不施钾肥(K0)和低钾(K1)等钾肥胁迫条件下模拟效果优于传统方法,其R2均大于0.991,RMSE均小于1.487,MAE均小于1.350,在K2—K5处理下机器学习模拟效果与传统方法的模拟效果接近,R2均大于0.993,RMSE均小于0.952,MAE均小于0.860。这主要是低钾胁迫下,玉米生长不稳定,传统的非线性回归方法在设定参数初始值范围时存在误差,而网格搜索法是在所有可能性范围内搜索最优参数,可找到一些因系统误差漏掉的有意义参数,从而改善模型精度。本研究通过机器学习方法仅对玉米吐丝期不同钾肥条件下的光响应曲线进行探讨,此方法较为通用,可较好作为其他各生育时期光合响应曲线的拟合方法。
4 结论
玉米吐丝期,叶片光响应参数表观量子效率α、最大净光合速率Pnmax、暗呼吸速率Rd、光饱和点LSP和光补偿点LCP对钾的响应趋势与光合参数相似,各处理在钾肥使用量达总量的75%(0、67.5、135、202.5、270、337.5 kg·hm-2)时,叶片光合参数Pn、Tr和Gs在K4(360 kg·hm-2)达到最大,而Ci、WUE和RE则相反。施钾量为360 kg·hm-2时,各处理光响应特征参数均达到最大。在低钾(0、90 kg·hm-2)胁迫下,机器学习的网格搜索法优于传统的非线性回归方法。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2017.11.015Magsci [本文引用: 1]

【目的】本研究旨在探讨大田淹水条件下夏玉米冠层结构和光合特性的变化规律。【方法】供试夏玉米品种为登海605(DH605)和郑单958(ZD958),通过设置3叶期(V3)淹水3 d(V3-3)和6 d(V3-6),拔节期(V6)淹水3 d(V6-3)和6 d(V6-6),以不淹水处理为对照(CK),比较不同淹水时期(V3和V6)和淹水持续时间(3 d和6 d)对夏玉米光合势、叶面积指数、净光合速率、冠层透光率及其半球灰度图像和产量的影响。【结果】淹水后夏玉米叶面积指数显著下降,群体透光率提高,群体光能截获率显著降低。V3淹水后夏玉米穗位层和底层透光率的提高幅度大于V6淹水,且其提高幅度随淹水持续时间的延长而增加。DH605和ZD958的V3-6处理的穗位层透光率较CK分别提高96.0%和70.2%,底层透光率分别提高了68.9%和71.9%。在淹水胁迫条件下夏玉米光合势和叶片净光合速率随淹水持续时间的延长而显著降低,V3淹水后净光合速率和光合势的下降幅度大于V6淹水。DH605和ZD958的V3-6处理在开花期的叶片净光合速率较CK分别下降23.5%和20.3%。DH605的V3-6处理在播种—拔节期、拔节期—大喇叭口期、大喇叭口期—开花期、开花期—乳熟期和乳熟期—成熟期各生育阶段的光合势较CK分别下降68.5%、45.0%、31.6%、25.0%和37.5%,ZD958分别下降62.4%、37.1%、25.8%、21.7%和38.5%。淹水后夏玉米光合势和叶片净光合速率的下降导致夏玉米光合同化物的积累与分配受到抑制,干物质积累量显著下降,成熟期DH605的V3-3、V3-6、V6-3和V6-6处理的干物质重较CK分别下降12.4%、24.8%、9.3%和21.1%,ZD958分别下降17.3%、26.7%、12.5%和23.9%。此外,淹水后夏玉米收获指数显著下降,3叶期淹水对其影响大于V6淹水,且影响随淹水持续时间的延长而加剧,DH605和ZD958的V3-6处理的收获指数较CK分别下降13.3%和13.8%。淹水后夏玉米冠层结构劣化与光合性能降低导致夏玉米产量显著下降。DH605的V3-3、V3-6、V6-3和V6-6处理的产量较CK分别下降23.2%、35.9%、17.0%和22.7%,ZD958分别下降20.0%、35.7%、15.0%和27.1%。【结论】淹水导致夏玉米群体光合势和叶面积指数显著降低,透光率提高,进而显著降低群体光能有效截获率和净光合速率,最终导致夏玉米产量显著下降。3叶期淹水对夏玉米冠层结构和光合特性的影响大于拔节期淹水,且其影响随淹水持续时间的延长而加剧。
DOI:10.3864/j.issn.0578-1752.2017.11.015Magsci [本文引用: 1]
【目的】本研究旨在探讨大田淹水条件下夏玉米冠层结构和光合特性的变化规律。【方法】供试夏玉米品种为登海605(DH605)和郑单958(ZD958),通过设置3叶期(V3)淹水3 d(V3-3)和6 d(V3-6),拔节期(V6)淹水3 d(V6-3)和6 d(V6-6),以不淹水处理为对照(CK),比较不同淹水时期(V3和V6)和淹水持续时间(3 d和6 d)对夏玉米光合势、叶面积指数、净光合速率、冠层透光率及其半球灰度图像和产量的影响。【结果】淹水后夏玉米叶面积指数显著下降,群体透光率提高,群体光能截获率显著降低。V3淹水后夏玉米穗位层和底层透光率的提高幅度大于V6淹水,且其提高幅度随淹水持续时间的延长而增加。DH605和ZD958的V3-6处理的穗位层透光率较CK分别提高96.0%和70.2%,底层透光率分别提高了68.9%和71.9%。在淹水胁迫条件下夏玉米光合势和叶片净光合速率随淹水持续时间的延长而显著降低,V3淹水后净光合速率和光合势的下降幅度大于V6淹水。DH605和ZD958的V3-6处理在开花期的叶片净光合速率较CK分别下降23.5%和20.3%。DH605的V3-6处理在播种—拔节期、拔节期—大喇叭口期、大喇叭口期—开花期、开花期—乳熟期和乳熟期—成熟期各生育阶段的光合势较CK分别下降68.5%、45.0%、31.6%、25.0%和37.5%,ZD958分别下降62.4%、37.1%、25.8%、21.7%和38.5%。淹水后夏玉米光合势和叶片净光合速率的下降导致夏玉米光合同化物的积累与分配受到抑制,干物质积累量显著下降,成熟期DH605的V3-3、V3-6、V6-3和V6-6处理的干物质重较CK分别下降12.4%、24.8%、9.3%和21.1%,ZD958分别下降17.3%、26.7%、12.5%和23.9%。此外,淹水后夏玉米收获指数显著下降,3叶期淹水对其影响大于V6淹水,且影响随淹水持续时间的延长而加剧,DH605和ZD958的V3-6处理的收获指数较CK分别下降13.3%和13.8%。淹水后夏玉米冠层结构劣化与光合性能降低导致夏玉米产量显著下降。DH605的V3-3、V3-6、V6-3和V6-6处理的产量较CK分别下降23.2%、35.9%、17.0%和22.7%,ZD958分别下降20.0%、35.7%、15.0%和27.1%。【结论】淹水导致夏玉米群体光合势和叶面积指数显著降低,透光率提高,进而显著降低群体光能有效截获率和净光合速率,最终导致夏玉米产量显著下降。3叶期淹水对夏玉米冠层结构和光合特性的影响大于拔节期淹水,且其影响随淹水持续时间的延长而加剧。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 4]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.17521/cjpe.2016.0067Magsci [本文引用: 3]
<p>长期以来, 光合作用机理模型中参数的确定都是一个难点。该文提出一种参数反演的方法, 称为近似贝叶斯法(APMC), 用来确定Farquhar光合模型的生理参数。通过将整个冠层抽象为一片大叶的思维抽象, 笔者进一步将APMC应用到冠层尺度的生理参数求解, 使直接求算冠层尺度生理参数成为可能。该文详细介绍了使用APMC估算光合模型参数的具体算法, 并用实测数据进行了验证。结果表明, APMC可以很好地应用于冠层光合模型参数的估计, 估计所得的参数落在参数生理上下限值之间, 应用1 948个实测数据进行检验, 得到决定系数0.75。模拟值和实测值的线性回归曲线斜率为1.04, 与理论上的1.0非常接近。这个方法对光合模型参数的获取或许有积极的意义。</p>
DOI:10.17521/cjpe.2016.0067Magsci [本文引用: 3]
<p>长期以来, 光合作用机理模型中参数的确定都是一个难点。该文提出一种参数反演的方法, 称为近似贝叶斯法(APMC), 用来确定Farquhar光合模型的生理参数。通过将整个冠层抽象为一片大叶的思维抽象, 笔者进一步将APMC应用到冠层尺度的生理参数求解, 使直接求算冠层尺度生理参数成为可能。该文详细介绍了使用APMC估算光合模型参数的具体算法, 并用实测数据进行了验证。结果表明, APMC可以很好地应用于冠层光合模型参数的估计, 估计所得的参数落在参数生理上下限值之间, 应用1 948个实测数据进行检验, 得到决定系数0.75。模拟值和实测值的线性回归曲线斜率为1.04, 与理论上的1.0非常接近。这个方法对光合模型参数的获取或许有积极的意义。</p>
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3773/j.issn.1005-264x.2010.06.012Magsci [本文引用: 1]
<FONT face=Verdana>光合作用对光和CO<SUB>2</SUB>响应模型是研究植物生理和植物生态学的重要工具, 可为植物光合特性对主要环境因子的响应提供科学依据。该文综述了当前光合作用对光和CO<SUB>2</SUB>响应模型的研究进展和存在的问题, 并在此基础上探讨了这些模型的可能发展趋势。光合作用涉及光能的吸收、能量转换、电子传递、ATP合成、CO<SUB>2</SUB>固定等一系列复杂的物理和化学反应过程。光合作用由原初反应、同化力形成和碳同化3个基本过程构成, 任一个过程均可对光合作用速率产生直接的影响。光合作用对光响应模型只涉及光能的转换, 而光合作用的生化模型包含了同化力形成和碳同化这两个基本过程。把光合作用的原初反应, 即把参与光能吸收、传递和转换的捕光色素分子的物理参数(如捕光色素分子数、捕光色素分子光能吸收截面、捕光色素分子处于激发态的平均寿命等)结合到生化模型中, 可能是今后光合作用对光响应机理模型的发展方向。</FONT>
DOI:10.3773/j.issn.1005-264x.2010.06.012Magsci [本文引用: 1]
<FONT face=Verdana>光合作用对光和CO<SUB>2</SUB>响应模型是研究植物生理和植物生态学的重要工具, 可为植物光合特性对主要环境因子的响应提供科学依据。该文综述了当前光合作用对光和CO<SUB>2</SUB>响应模型的研究进展和存在的问题, 并在此基础上探讨了这些模型的可能发展趋势。光合作用涉及光能的吸收、能量转换、电子传递、ATP合成、CO<SUB>2</SUB>固定等一系列复杂的物理和化学反应过程。光合作用由原初反应、同化力形成和碳同化3个基本过程构成, 任一个过程均可对光合作用速率产生直接的影响。光合作用对光响应模型只涉及光能的转换, 而光合作用的生化模型包含了同化力形成和碳同化这两个基本过程。把光合作用的原初反应, 即把参与光能吸收、传递和转换的捕光色素分子的物理参数(如捕光色素分子数、捕光色素分子光能吸收截面、捕光色素分子处于激发态的平均寿命等)结合到生化模型中, 可能是今后光合作用对光响应机理模型的发展方向。</FONT>
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}