 ,1, ����1, �ߴ��1
,1, ����1, �ߴ��1Effects of Artificial Warming from Late-Winter to Early-Spring on Photosynthesis and Flag Leaf Senescence of Winter Wheat
YAN Peng1, SUN XiaoNuo2, DU Xiong,1, GAO Zhen1, BIAN DaHong1ͨѶ����:
�ո�����:2019-03-11��������:2019-04-25�����������:2019-08-01
| ��������: |
Received:2019-03-11Accepted:2019-04-25Online:2019-08-01
����� About authors
����,E-mail��

ժҪ
�ؼ��ʣ�
Abstract
Keywords��
PDF (604KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر���
�������ø�ʽ
����, ��Сŵ, ����, ����, �ߴ��. �����紺�����¶Զ�С�������ܼ���Ҷ˥�ϵĵ�������[J]. �й�ũҵ��ѧ, 2019, 52(15): 2581-2592 doi:10.3864/j.issn.0578-1752.2019.15.003
YAN Peng, SUN XiaoNuo, DU Xiong, GAO Zhen, BIAN DaHong.
0 ����
���о����塿����ƽԭ�������ҹ���Ҫ�Ķ�С����ֲ��,�����ڴ������¿�ʹС���뷢��ʱ������,��������������[1];����������������������ܸ��ȷ�Σ��,ʹ�ཬʱ������,�������ؽ���[2],�ӱ���λ���С������������ں��ϡ�ɽ����ʡ�������һ����ĸ���ԭ�������¶ȱ仯ͬС�������������������������ġ����Գ��ԡ�[3],������������ʹС�������ҷ������,���������¿졢�¶ȸ���ʹС�����Խ��͡��ཬʱ����ҿ���˥�ϡ����,ͨ�������紺���´�ʹС��������������,������¶����ߺ�����Խ���,�Ӷ�ʹ�����¶��붬С��������������Ӧ,����ھ�С�����DZ����������������ԭ�����������������Ӱ�����ʮ����Ҫ�����塣��ǰ���о���չ����С�������¶ȱ仯����Ӱ����С�������������˥�Ͻ���,����Ӱ��������ʻ��ۺ�ϲ�����������ת��[4]�����о�����,�ν�ǰ���»�ʹС������������������[5],����ֲ��MDA������������,�����ӿ���С��˥�Ϲ���[6],�ν�ǰ����в�Ȼ��ᵼ������������ҶҶ���غ�����������ʡ��������ʺ�ҶƬ�����ȵ�ָ��������½�[7]��������ȫ���������¿���������С��������ǧ����[8],����ҹ��������ʹ��С���������ǰ�����������̡��ཬ�������̶�����[9]���о�����,��С����������Ҷ������ʺ����ʻ���������[10,11],Դ��֮�����ʷ������ʧ��[12],��ɲ�����Ʒ��Эͬ���͵ĸ���ЧӦ[13,14]��ԭ�����ڻ�����½�������Ҷ�����Ե�������SOD��POD��CAT�Ļ���,MDA��������,��ʹ��Ҷ��˥[15]�����̽����α���С���ڰν�ǰ���ºͻ�����µ�в�ȵ��·������˽�һ����߲����Ĺؼ����ڡ������о�����㡿Ŀǰ,�����¶ȶ�С������γɼ�˥��ЧӦ���о��ּ����ڶ����������ȫ���������µȷ���,�����������紺�������¶���Ҷ������Լ�˥�ϵ��о�,���С������γɵ�����ԭ��Ҳ�д���һ��̽�����������Ĺؼ����⡿���о��ڴ��ﻷ����,�ڶ�С����ɴ�����������紺,ͨ������ܱոֹ�����ʵ����������ǰС���������������,�νں�ҳ���Ĥ��Խ����緢С�������Ļ����¶�,���������紺�������¶Զ�С����Ҷ������Լ���˥�����Ե�Ӱ��,����Ϊ���������ھ�С�����DZ����ͨ�������¶��ӻ�С��˥���ṩ���ۺͷ������ݡ�1 �����뷽��
1.1 ����ظſ�
���������2015��2017������2���������ںӱ�ʡũ�ֿ�ѧԺ���ݺ�����ˮũҵ����վ��37��54��12.5��N,115��42��10.94��E���С���������վ��ƽ������ 12.7��,�������������Ϊ���ʳ���,�����������л���12.53 g?kg-1����65.8 mg?kg-1����Ч��15.3 mg?kg-1����Ч��121.9 mg?kg-1������̬����������������������ƽԭ��1.2 �������
1.2.1 ������ ���á����35����Ϊ�������,��ÿ���10��8�ղ��֡�С����ǰ,ǰ�����ո�ȫ�����鷭ѹ����,��135 kg N ?hm-2��135 kg P2O5?hm-2��180 kg K2O?hm-2ʩ��,�ν������ˮ����N��135 kg?hm-2�����þ���С�����ֻ����в���,������187.5 kg?hm-2,15 cm���о���ֲ,����������ʩͬ���ظ߲��1.2.2 ���鴦�� ������4������,�ֱ�ΪM2E������1��25�ո���2����Ĥ,�����ڲ��пס������,3��5�սҳ������Ĥ,3��25�սҳ��ڲ��п�Ĥ��;M2L������2��5�ո���2����Ĥ,�����ڲ��пס������,3��5�սҳ������Ĥ,3��25�սҳ��ڲ��п�Ĥ��;M1������2��20�ո���1������Ĥ,3��25�սҳ�����CK��������գ���ÿ����3���ظ�,��12��С��,�����������,С�����Ϊ16 m2��4 m��4 m��,С��֮����0.8 m�۲����С����,�����Ϊÿ�����ȴ������30 cm�ĸֹ����ң�4 m��4 m��,�������渲������Ĥ,�Ը����пס�����Ĥ�Լ�����ʱ��������ͳ����������������¶Ⱥ�С������ʼʱ��,��Ĥ���Ǻ�ѹʵ����,������Ӱ�������¶�,����������µĻ���,ͨ��������Ĥ���������������¶ȡ��ɴ˻����ʱ����ͬ��С�������������̲�ͬ������������ͬ���������ں��¶Ȳ�ͬ�Ľ����
��Ĥ��Ⱦ�Ϊ0.03 mm�Ұ�ɫ��,��Ĥ�ɵ�����ȡ���ɣ���ֱ��1 cm,�ڼ��Ϊ2 cm���ң������Ҵ��,¶�غ���������MicroLite5008��U��ʽ�¶ȼ�¼�Dzⶨ�¶�,�¶ȼ�¼�Ƿ�����С��Ҷ����,ÿСʱ�Զ���¼1���¶�,�Դ˼�����������½��վ��¶ȣ�ͼ1������������ǰ8 d������32 d�վ��¶ȣ�ͼ2����
ͼ1
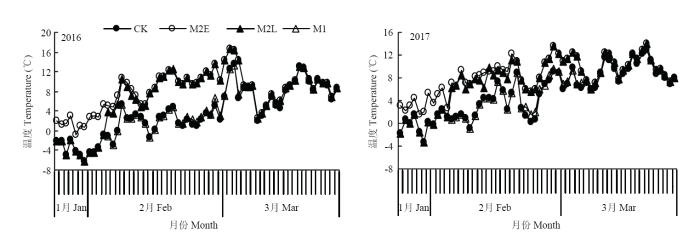 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ12016��2017�괦�����½��վ��¶�
Fig. 1Daily average temperature of treatment heating stage in 2016 and 2017
ͼ2
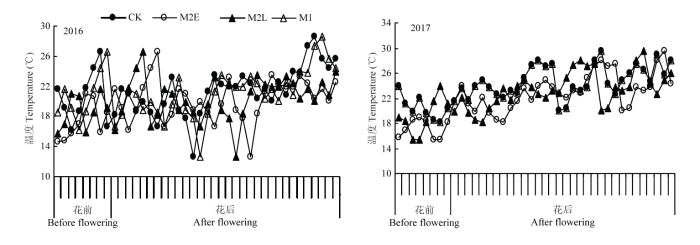 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ22016��2017�괦������ǰ8 d������32 d�վ��¶�
Fig. 2Daily average temperature from 8 days before flowering to 32 days after flowering in 2016 and 2017
M2E��M2L��M1��3�������ڸ������½�2016��ֱ����ӻ���274.3�桢204.4���15.3��,2017��ֱ����ӻ���189.9�桢140.6���20.3�档
����ǰ8 d������32 d����M2E��M2L��M1��CK�Ļ��¼�����2016�ֱ�Ϊ62.07�桢46.64��,��7.13��,2017��ֱ�Ϊ76.1�桢53.79������͡�
1.3 �ⶨ��Ŀ������
1.3.1 ����ʱ�ڵ��� �۲첢��¼������������ʱ�ڳ��ֵ�����,�ﻨ�ڱ�Ǹ�С���ڿ���ʱ����ͬ������200�ꡣ��������ˮ������18%ʱΪ����,��ʱС������ݻ�,������ɫ��Ӳ��1.3.2 ��ҶҶ����IJⶨ �ڿ�����ÿС��������Ҷ20Ƭ,3���ظ�,��Ҷ���=Ҷ�������Ҷ����0.83[16]��
1.3.3 ��Ҷ������ԵIJⶨ ��������CID��˾������CL-340�ֳ�ʽ������ò���ϵͳ,�ڿ���ʱ�ͻ����8��16��24��32�������10��00-11��00������Ҷ��������ʣ�Pn���������ȣ�Gs���������ٶȣ�Tr����ϸ����CO2Ũ�ȣ�Ci����ʹ���ձ���SPAD-502Ҷ�����DzⶨҶ������Ժ�����
1.3.4 ��Ҷ��������ָ��IJⶨ ���������� ÿ8 dȡ���ֲ�����Ҷ,��Һ���ٶ�������-80�泬���±����б��档��0.5 g�����ҶƬ��������Ԥ����в���,����1 mL���Ỻ��Һ,�ڱ�ԡ����ĥ�ɽ�,�ӻ���Һ�����������Ϊ5 mL,4��������10 000 r/min����20 min,ȡ����Һ����ø���Բⶨ�����õ������ۣ�NBT���ԭ��[17]�ⶨSOD����,Sigma[18]���ⶨPOD����,��ʿ�ܵ�[19]�Ľ��ķ����ⶨCAT���Լ�MDA������
1.3.5 �����������乹�����صIJⶨ ������ÿС��������ȡС��20��,�ⶨ��������ѡȡ�м�δ��ȡ���ƻ�������,��1 m�������ڼ����������ۺϵ�λ�������,3���ظ�,��������ջ����������г��غͲⶨ������ˮ��,��14%�ĺ�ˮ�����㵥λ���������
1.3.6 �����ں�ˮ���ⶨ��ˮ������Ч�ʼ��� �ڲ���ʱ���ջ��������ÿ10 cmΪһ����ȡ0��200 cm��������,���ú�ɷ��ⶨ������ˮ����С��ȫ�����������Զ�����վ���ټ�⽵ˮ��,2015��2016��2016��2017��������ˮ���ֱ�Ϊ62.3 mm��99.8 mm���������й�ʽ����С������ˮ����ˮ������Ч��[2]��
������ˮ����mm��= �����ȣ�cm�����������أ�g?cm-3��������������ˮ����10;
����ˮ����mm��= ����ʱ������ˮ����mm��+��ˮ����mm��+��ˮ����mm��-�ջ�ʱ������ˮ����mm��;
ˮ������Ч�ʣ�kg?hm-2?mm-1��= ����������kg?hm-2��/����ˮ����mm����
1.3.7 ����ÿ8 d���º�����Լ�ø����������ϵ�Ľ��� ��M2E��M2L��M1��CK����ÿ8 d����Ϊ������,��8 d�й�����Լ�ø���Ե�ƽ��ֵΪ�����꽨��ɢ��ͼ,Ȼ�����ӻ���8 d��������Ҷ������Լ�ø���ԵĶ�����ʽ������,��������վ�������Ҷ������Լ�ø���Ե�������ϵ��
1.4 ���ݴ���
����Excel 2010������������ͳ�Ʒ�������ͼ,����SPSS�������в�ͬ�����IJ��������Լ���(LSD��)������2015��2017�����Ҷ������Լ�ø���Ա仯��������,����ͼ�����ݾ�Ϊ2������ƽ��ֵ�����2 ���
2.1 ��С������Ҷ����Ϳ���������ʱ��
����1���Կ���,�����紺���µ�������Խ��,С��������ǰ��Խ����,�ҿ���ʱ����Ҷ�����Խ��;���ҽҳ�����Խ������緢С�������������¶�,���µĽ�����Խ��,С��ཬʱ����ӳ�Ч����Խ������2�������M2E������CK������ǰ��8��9 d,��Ҷ���������24.6%��25.5%,�ཬʱ���ӳ���5��6 d;M2L��������ʱ��Ҷ�����CK�����14.6%��20.3%,������ǰ��6��7 d,�ཬʱ���ӳ���4 d;��������ʱ����Ƴٺ���ʱ�������,��Ҷ����������ͳ���������CK�IJ���Խ��ԽС,������M1������CK�������������졣����������紺�����Ľ�������ʹС������������Ҷ��������翪�����ڡ��ӳ��ཬʱ�䡢��ø����ϲ���,ΪС��߲�����������Table 1
��1
��1С������Ҷ����Ϳ���������ʱ��
Table 1
| ������ Growing season | ���� Treatment | ��Ҷ��� Flag leaf area (cm2/leaf) | �������� Anthesis date (M-D) | �������� Maturity date (M-D) | ����-�������� Anthesis-maturity duration (d) |
|---|---|---|---|---|---|
| 2015-2016 2016-2017 | M2E M2L M1 CK M2E M2L M1 CK | 21.91a 21.01a 17.89b 17.46b 22.17a 20.40b 17.87c 17.80c | 4-26 4-28 5-3 5-4 4-28 5-1 5-7 5-7 | 6-4 6-5 6-7 6-7 6-6 6-7 6-10 6-10 | 40 39 36 35 40 38 34 34 |
�´��ڴ�|����CSV
2.2 ��С����ҶҶ������Ժ������������
2.2.1 ��ҶҶ������Ժ���SPAD ��ͼ3���Կ���,����������ҶҶ������Ժ��������������̵��ƽ����½�,ͬһʱ�䲻ͬ�����������������½�����ʱ��ϳ���M2E��M2L�����ڹ۲���������ҶҶ������Ժ���ȫ����������M1������CK,M2E��M2L������Ҷ������Ժ�����CK�ڻ���16 d�ֱ������19.0%��13.3%;����24 d�ֱ������15.6%��12.2%;����32 d�ֱ������56.4%��49.9%�������½�����ʱ��϶̵�M1������CK֮������첻����������������紺�����º�ٽ��������̵�Ч���������緢С��ཬ�ڼ���Ե��µĻ���,��Ч��������ҶҶ���ص�˥���ٶȡ�ͼ3
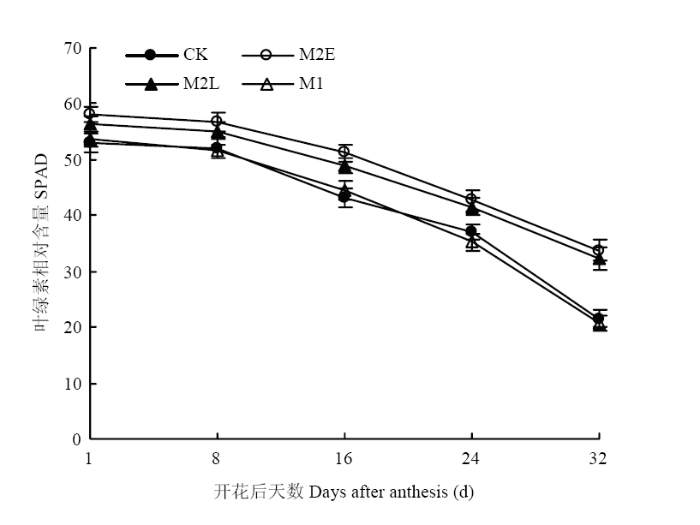 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ3������ͬ��������ҶҶ������Ժ���SPADֵ
Fig. 3SPAD value of chlorophyll content in flag leaves after flowering under different treatments
2.2.2 ��Ҷ������� ��ͼ4���Կ���,��������Ҷ��������ʣ�Pn����ҶƬ�����ȣ�Gs�������������̶��½�,�������ʣ�Tr����������֮���漴�½�,ϸ���������̼Ũ�ȣ�Ci��һֱ����,�Ҵ���������������졣����24 d��,M2E������������ʱ�CK�����16.0%��31.5%,������������10.9%��20.8%,�����ٶ�������12.6%��22.7%,ϸ����CO2Ũ�Ƚ�����11.4%��19.7%�����Ŵ���ʱ����Ƴټ�����ʱ�������,Pn��Gs��Tr�����ӷ��ȼ�Ci���½�����������С,���»���Pn������3��ָ����M1��CK�����������졣��ͼ5��֪,�����������ҶPn��Gs�����������,��Ci�����������,�������վ���Ϊ25.9��26.0��ʱ,���������Ѹ�ٽ��Ͳ��ﵽ��Сֵ������CK��M1���������Ļ����¶Ƚϸ�,��������Ҷ�����������������,��������ͬ�����ڵ�M2E��M2L���������Ļ����¶���Խϵ�,������Ҷ��������ʵ���������ߡ��ɴ˿ɼ�,�����紺�����������������Ҷ�������,Ϊ��߹ཬ�ٶȺͽ��������ṩ�����ʻ�����
ͼ5
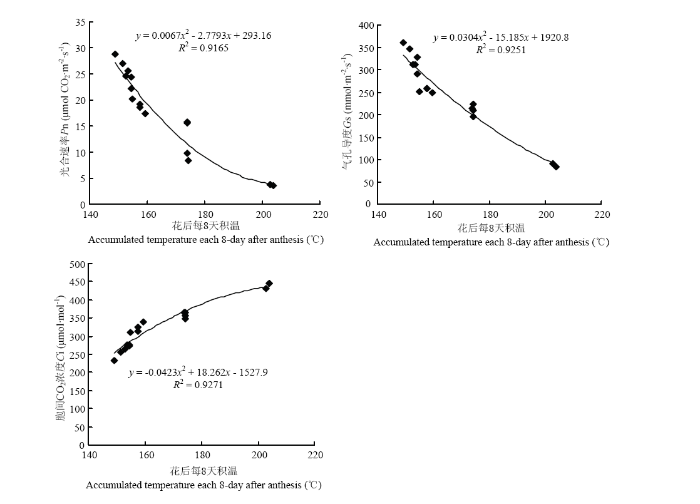 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ5������ÿ8����º�����Թ�ϵ
Fig. 5Relationship between accumulated temperature and photosynthetic characteristics every 8 days after flowering
ͼ4
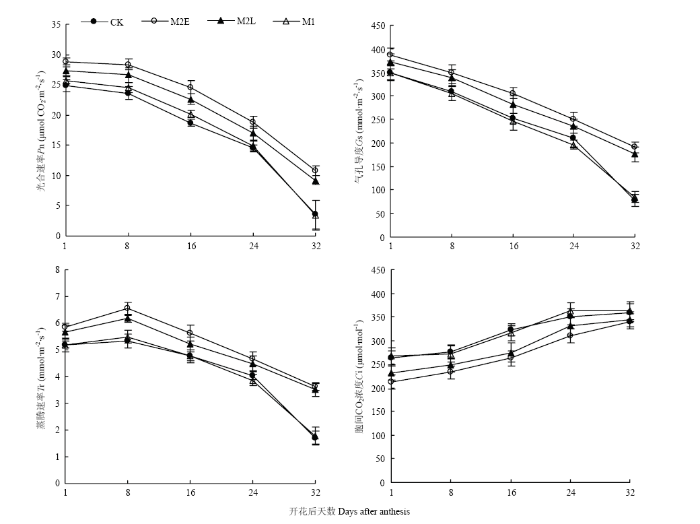 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ4������ͬ��������Ҷ������Ա仯
Fig. 4Changes of photosynthetic characteristics of flag leaves under different treatments after flowering
2.3 ��Ҷ������ø���Լ��뻨���¶ȵĹ�ϵ
��ͼ6���Կ���,��������ҶSOD��POD��CAT���Ծ������������̵��������������½�,�һ����8��ﵽ���,����ָ�꼰����ڴ���������������졣����24 d ��,M2E��M2L�����Ŀ�����ø���Ծ���������CK,������ʱ����Ƴ�,M2L���������Ҷ������ø���Ե�Ч����M2E�����½�,���������������졣M2E������CK���,SOD����ƽ�������15.0%��POD����ƽ�������14.2%��CAT����ƽ�������17%;M2L������CK��ȵĽ������,ֻ�DZ仯���������½�,SOD��POD�Ļ���ƽ�������13.1%��9.1%,CAT�����������12%����������ʱ��ļ����Ƴټ�����ʱ�������,M1��CK�������������졣����24��32 d,����M1��CK�������¶Ƚϸ�,���¿�����ø������������,��ҶѸ��˥�ϡ���ͼ7��֪,��Ҷ������ø�����뻨����³����������,�������վ��·ֱ�ﵽ33.0�桢25.6�桢27.7��ʱ,�ᵼ��SOD��POD��CAT�Ļ���Ѹ�ٽ��Ͳ��ﵽ��Сֵ�������紺����������ǰ��С���������������,�Ƴ����Һ�����緢��С������Խ����������������¶�,��������������Ҷ������ø�Ļ���,�Ӷ��ӻ���С���˥�ϡ�ͼ6
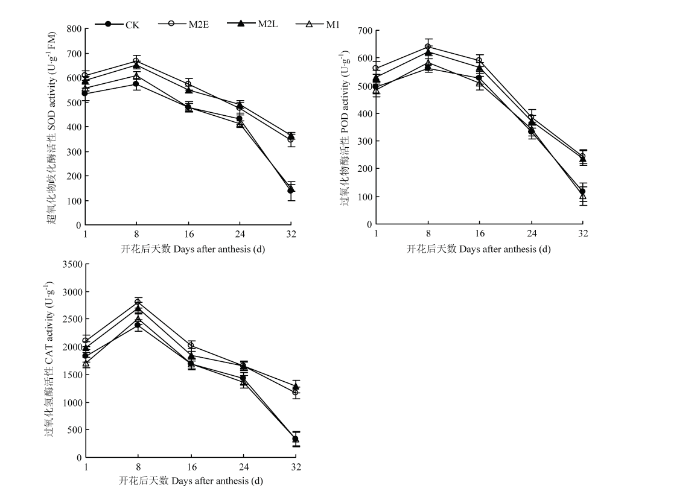 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ6������ͬ��������Ҷ������ø���Ա仯
Fig. 6Changes of antioxidant enzyme activities in flag leaves treated with different treatments after flowering
ͼ7
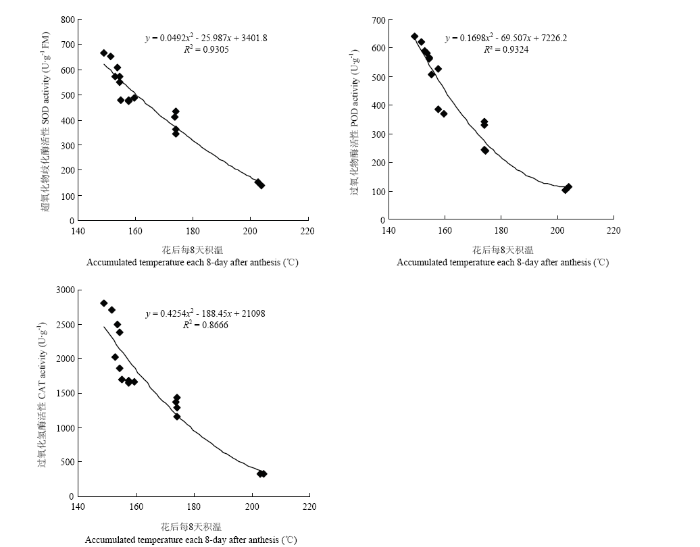 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ7����ÿ8����ºͿ�����ø���Թ�ϵ
Fig. 7Relationship between accumulated temperature each 8-day after anthesis and antioxidant enzyme activity
2.4 ��ҶĤ֬�������̶ȼ��뻨���¶ȵĹ�ϵ
��ͼ8���Կ���,����������ҶMDA���������������̾�����������,����1��8 d���ӻ���,����8��32 d���ӽϿ�,����������ﱾ����˥�ϻ����й�,˥��ʱ����������������˥�ˡ�����8 d��,�������MDA�������첻����;����8�Ժ�,�������MDA������������,����M2E��M2L������MDA��������������CK��M1����,M2E��M2L��MDA������CK�ֱ���20.2%��14.6%����ͼ9��֪,��ҶMDA�����뻨����³���������ع�ϵ,�ϸߵĻ����¶����MDA����Ѹ������,�������վ��´ﵽ28.2��ʱ,MDA�����ﵽ���ֵ,�����Ҷ����˥�ϡ����о����õ������紺�������ºͽ�Ĥ����Խ������緢С���¶ȵķ���,��Ч��������ҶMDA�ĺ���,�ӳ�����Ҷ�����ڡ�ͼ8
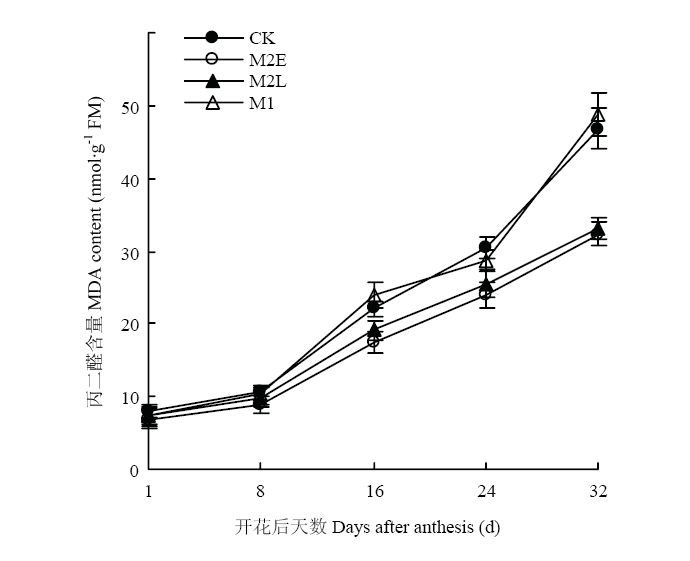 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ8������ͬ��������ҶMDA����
Fig. 8MDA content in flag leaves under different treatments
ͼ9
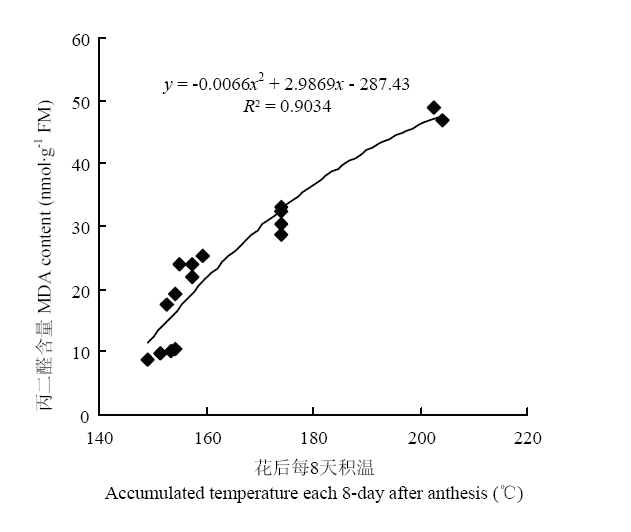 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ9����ÿ8����º�MDA������ϵ
Fig. 9Relationship between accumulated temperature each 8-day after anthesis and MDA content
2.5 ����������ˮ������Ч��
����2���Կ���,2���������M2E��M2L�����IJ����������������ؾ�����M1������CK,������������ǧ���غͲ����������������ڲ�����ߵ�M2E����,��CK���2016������1 723.1 kg?hm-2,����Ϊ20.5%;2017������1 857.6 kg?hm-2,����Ϊ22.8%��2��������ݲ����������ضԲ��������ʵ�˳���Ϊǧ����>������>��λ�������������������紺������������߲�������Ҫԭ����������ǧ���غ�������,��������ʱ��Խ����ʱ����Խ��,��߲�����Ч����Խ������Table 2
��2
��2��ͬ�����Ķ�С�������ˮ������Ч��
Table 2
| ������ Growing season | ���� Treatment | ���� Spike number (104��hm-2) | ������ Kernel number per spike | ǧ���� 1000-kernel weight (g) | �������� Grain yield (kg��hm-2) | ����ˮ�� Field water consumption (mm) | ˮ������Ч�� Water use efficiency (kg��hm-2��mm-1) |
|---|---|---|---|---|---|---|---|
| 2015-2016 2016-2017 | M2E M2L M1 CK M2E M2L M1 CK | 614.7a 611.4a 608.6a 614.4a 613.6a 610.4a 600.6a 597.3a | 37.5a 37.2a 34.0b 33.8b 36.0a 35.8a 33.6b 33.1b | 46.7a 46.5a 41.3b 40.9b 46.0a 44.6b 41.1c 40.8c | 10160.9a 9905.1a 8492.1b 8437.8b 9995.4a 9691.1b 8198.4c 8137.8c | 462.2a 458.3a 442.7b 441.4b 481.2a 472.8b 460.8c 456.5c | 22.0a 21.6a 19.2b 19.1b 20.8a 20.5a 17.8b 17.8b |
�´��ڴ�|����CSV
�����紺����������������ͬʱ,Ҳ��Ӧ������С�������ˮ��,����С���������ٶ�Ҫ���ں�ˮ������,������������С��ˮ������Ч�ʡ����ڲ�����ߵ�M2E����,��CK���,2016������20.5%,��ˮ������4.7%,ˮ������Ч�����15.2%;2017������22.8%,��ˮ������5.4%,ˮ������Ч�����16.9%������������紺������������������ͬʱ,Ҳ�����ٽ���WUE����ߡ�
3 ����
С�������������γ���Ҫ������Ҷ�Ĺ��ͬ������[20],���ཬ�ڸ��»�������Ҷ�����,������Ҷ���غ���,�������¹�����������½�[21,22],ͬʱ����Ҳ����ɹ�ϲ�������ӿ�,ͬ��Ч�ʽ���,�������²�����Ʒ���½�[23]�����о���,�ڴ���������С����������紺��������������С���༰����������������,����ʹС����ǰ����Ҷ�����������;�ҳ���Ĥ����Խ������緢С��Ļ����¶�,�ּ�����Ҷ���طֽ��ת���ٶ�,���������С����Ҷ�Ĺ������,Ϊ��ཬ�ṩ�˸���Ĺ�ϲ���¸��Ƶ�[24]�о���Ϊ,������������CO2Ũ�ȳʸ����,˵������������ǿ���Ӷ�������Ҫ��Ҷ��ϸ���Ĺ�ϻ�������Ľ��,������Ci���͵Ľ�������о���,�����¶�����ҶPn��Gs�����������,��Ci�����������,�����Ż�����µĽ���,Pn��Gs����,Ci�������½�,�������ҶPn��������ҪԴ�ڷ���������,�Ʋ�������ͺ�ϵͳ���˳̶ȼ�Ҷ����˥���̶��йء�����������,ֲ��ϸ���ڵĻ�������ROS�����ϵͳ�Ὣ������ROS�����ά��ϸ����������ԭ״̬��ƽ�⡣������в�Ȼ��������ƽ��,ʹϸ����ROS��������,���ƻ�ROS���ϵͳ[25],��������Ĥ֬�������̶ȼӾ�,��Ĥ���������,��Ҷ˥�ϼӿ�[26,27],��������½�,�л����������,���յ������ؽ���[28]�����о���,�����紺����������ǰС�����������ͽ�Ĥ�������Ե���,��������˻������������Ҷ����øSOD��POD��CAT�Ļ���,������MDA����,�����˻����������������������ɻ���ϸ��Ĥϵͳ���˺�,ʹϸ��ά���ڿ��������ĵ���״̬,�ӻ�����Ҷ˥��,����������ʱ����Ƴٺͳ���ʱ�������,��Ҷ˥���ӻ���Ч��������ͬʱ���о�Ҳ����,�����¶�����Ҷ������ø���Գ����������,��MDA���������������,�����Ż�����µļ���,��Ҷ������ø�������,Ĥ֬�������̶Ƚ���,�Ӷ�˥���ٶ������仺��
ǰ���о�����,����ǰ6��8 d���´���С��,��ʹС�������½�60%����,����������������������,���Ҹ��¶�С�����Ե�Σ�������ۼ�ЧӦ[29];WARDLAW��[30]�о����������ڸ�����������������,��Ҫԭ������С�����Ρ����۰�������֯��ˮ��С�������ཬ�ڵij���Ҳ��Ҫȡ�����¶�,�¶���������ҶƬ�ر�����Ҷ����˥[31,32],�����ӿ�ཬ����,���̹ཬʱ��,��������½�[33]�����о���,2015��2017����������ȴ�����С��������ƽ���¶ȷֱ�Ϊ22.3���24.1��,���������ֱ�Ϊ35 d��34 d,�վ��´���25.0��������ֱ�Ϊ1 d��11 d;������Ч����õ�M2E��������������ƽ���¶ȷֱ�Ϊ21.0���22.3��,����������Ϊ40 d,�վ��´���25.0��������ֱ�Ϊ1 d��8 d���������´�����С����������ǰ,��Ĥ�ҳ�����Խ������緢С���������¶�,�Ӷ������������Ч������,�ӻ�����Ҷ˥��,�����ӳ��˹ཬʱ��,����������,����ʹ��������ǰ,��Ч������С�������������ܸ��ȷ��Ӱ��̶ȡ�
�о�����,�˹�����Խ��ǰ����´���60��ʱ,���뷢���������Լӿ�,��С���������ǰ,ʹС����������������,�������ܴ�������Σ��,���С��������������[3]�����о���,2��������ȳ�����ն�С��ν�������ƽ���¶ȷֱ�Ϊ17.0���17.3��,���������ֱ�Ϊ31 d��34 d,���亦����;������Ч����õ�M2E�����ν�������ƽ���¶ȷֱ�Ϊ14.3���14.0��,���������ֱ�Ϊ40 d��42 d,Ҳ���亦�������������紺�¶Ƚϵ�ʱͨ����������´�ʹС�����緢��,�������¶����ߺ��Ĥ����,���������ڵ��¶Ȳ���������̫��,��3�µ��¶��ȶ����ȫ���ҳ���Ĥ,��ɱ���С�����緢����������Σ��,���ڽ�Ĥ����Խ����˰ν���������¶�,�Ӷ��ӻ�����ֻ�,��������ֻ�ʱ��,����������������[34]����ȫ�������ů������,���ֵ�[35]ͨ��SUCROSģ���о�����,�ƻ���ƽԭ�������ȫ�������ʶ�����,��С������ȫ����������,���������������ӳ��Ľ��,�������������������ʻ���;����������ʹ�ö�С��Ӳ��������������������������������,��ʹС�����������½������о���,ͨ�������紺��������,������С��ԭ������¶��������õ�ʱ��,����ӳ��˷��ൽ�����ڵ����ڿ��,�����ӳ�����Ҷ��Ϲ�����,�������˸߲���
4 ����
�����紺��1�µ���3�µף���������ʹС��������8 d,�Ƴ����Һ����Խ���,��Ч�ӳ��˺����������εij���ʱ��,���������С���������;������Ҷ����ȶ���������27%,�������������Ҷ���������绯ø��SOD������������ø��CAT������������ø�Ļ��ԣ�POD��,��Ч��������Ҷ��˥�ϡ��ཬ����ҶҶ������Ժ����ȶ�������13.3%,��������������24.9%,�Ӷ�����˸��ߵ����ء������紺�������º���������������ԴͬС��������������,����������������22.8%,ˮ������Ч�����16.9%������Ч�����ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
[��������: 1]
[��������: 1]
DOI:10.5846/stxb201404100697URLMagsci [��������: 2]

ȫ������仯������,ũҵ�����ֺ�������̬�ơ����ȷ��ֺ�������������ǿ�ȶ����������Եı仯���о����ȷ��ֺ���ũ�����Ӱ������ҹ�ũҵ�ɳ�����չ��������ʳ��ȫ�Ⱦ�������Ҫ����ʵ���塣���ûƻ�������68������̨վ1961-2010���������������,��54��ũҵ��������վ1981-2006��С��ķ����ڡ����������ȷ��ֺ�������,���ù��ϵ��й������2007�귢����������ҵ����С����ȷ��ֺ��ȼ����ж�С����ȷ��ֺ�ָ��,������ȷ�Σ��ָ��,��һ��ϸ��������,ȷ����С�����ǰ�����������������Ӱ��Ĺؼ���������,������ȷ��궬С���������,�����ضȸ��ȷ�Ӱ���¸��ȷ�Σ��ָ���붬С�����-����������������������Ӱ���ͳ��ģ��,����1981-2006��ƻ���������С����ȷ�������������������:(1)�ضȸ��ȷ�Σ����,1981-2006���ڼ�ƻ�����������С��ͬ����ʱ�εĸ��ȷ�Σ��ָ��ƽ���ڳ���-����ʱ�����,����-����ʱ�ξ���,����-����ʱ����С,�ֱ�Ϊ0.17��0.15��0.14,ƽ��0.15;(2)��С�����ǰ�����������������Ӱ��Ĺؼ���������Ϊ:����-�����ڼ��������¡��ν�-�����ڼ��ƽ�����º�����-�����ڼ��ƽ������, �������������ϵ���ֱ�Ϊ0.64��0.86��0.99,���ﵽ������ˮƽ�����в���-�����������¿ɾ���С�����ij��������;�ν�-�����ڼ�,��С��ԭ���γ���-�ķ����γ�������ƫ�Ϳ��ӳ�С�롢С���ֻ�ʱ��,��ֹ�˻�,��߽�ʵ��;����-�����ƽ������ƫ���������������,�ӳ����ڹཬʱ��,�����������ڿ����ڷ�;(3)������ȷ��궬С�����������,�������ضȸ��ȷ�Ӱ���¸��ȷ�Σ��ָ���붬С�����-����3���������������������Ӱ���ͳ��ģ��,��֤���������ģ�Ϳ����ܹ��ۺϵط�ӳ���ȷ��ڲ�ͬ�����ζ�С�������Ӱ�졣��һ��������������:�ضȸ��ȷ�Σ����,�ƻ���������С���������21.52%-39.80%֮��,ƽ��Ϊ27.83%��
DOI:10.5846/stxb201404100697URLMagsci [��������: 2]
ȫ������仯������,ũҵ�����ֺ�������̬�ơ����ȷ��ֺ�������������ǿ�ȶ����������Եı仯���о����ȷ��ֺ���ũ�����Ӱ������ҹ�ũҵ�ɳ�����չ��������ʳ��ȫ�Ⱦ�������Ҫ����ʵ���塣���ûƻ�������68������̨վ1961-2010���������������,��54��ũҵ��������վ1981-2006��С��ķ����ڡ����������ȷ��ֺ�������,���ù��ϵ��й������2007�귢����������ҵ����С����ȷ��ֺ��ȼ����ж�С����ȷ��ֺ�ָ��,������ȷ�Σ��ָ��,��һ��ϸ��������,ȷ����С�����ǰ�����������������Ӱ��Ĺؼ���������,������ȷ��궬С���������,�����ضȸ��ȷ�Ӱ���¸��ȷ�Σ��ָ���붬С�����-����������������������Ӱ���ͳ��ģ��,����1981-2006��ƻ���������С����ȷ�������������������:(1)�ضȸ��ȷ�Σ����,1981-2006���ڼ�ƻ�����������С��ͬ����ʱ�εĸ��ȷ�Σ��ָ��ƽ���ڳ���-����ʱ�����,����-����ʱ�ξ���,����-����ʱ����С,�ֱ�Ϊ0.17��0.15��0.14,ƽ��0.15;(2)��С�����ǰ�����������������Ӱ��Ĺؼ���������Ϊ:����-�����ڼ��������¡��ν�-�����ڼ��ƽ�����º�����-�����ڼ��ƽ������, �������������ϵ���ֱ�Ϊ0.64��0.86��0.99,���ﵽ������ˮƽ�����в���-�����������¿ɾ���С�����ij��������;�ν�-�����ڼ�,��С��ԭ���γ���-�ķ����γ�������ƫ�Ϳ��ӳ�С�롢С���ֻ�ʱ��,��ֹ�˻�,��߽�ʵ��;����-�����ƽ������ƫ���������������,�ӳ����ڹཬʱ��,�����������ڿ����ڷ�;(3)������ȷ��궬С�����������,�������ضȸ��ȷ�Ӱ���¸��ȷ�Σ��ָ���붬С�����-����3���������������������Ӱ���ͳ��ģ��,��֤���������ģ�Ϳ����ܹ��ۺϵط�ӳ���ȷ��ڲ�ͬ�����ζ�С�������Ӱ�졣��һ��������������:�ضȸ��ȷ�Σ����,�ƻ���������С���������21.52%-39.80%֮��,ƽ��Ϊ27.83%��
DOI:10.3724/SP.J.1006.2016.01530URLMagsci [��������: 2]
<p>��Ի���ƽԭ���������¶ȱ仯�붬С�������������������¶ȼ��ì�ܣ��Լ��ɴ���ɵĶ�С����ԵͲ����⣬��2012—2013��2013—2014����������������ͨ�����������о��������紺����ҽ�����������¶ȶ�С�������Ӱ�졣���������2��20��ǰ��������Ĥ����ÿ��ǰ1�ܻ������23.0~49.7�棬1����Ѯ��3����Ѯ���������½��ڿ����ӻ���167.7~176.8�棬�Ӷ�С������������ǰ����Ĥ�ҳ����γɵ���Ե��»���ʹ�����������γ���ʱ���ӳ�4~10 d���볣����ֲ�����գ���ȣ��������´����Ŀ����ڸ����ʲ������18.8%��Ҷ���ָ�����14.7%��������������43.6%����������ʸ�ֵ�������ӳ�10 d���ϣ������������ε��ӳ��ٽ��˸����ʻ��ۺ�������ת�ƣ���Ч��������48~98��hm<sup><span>-</span></sup><sup>2</sup>��������������3.9~4.5����ǧ��������2.5~5.6 g����ȫ���������½��ٵ�2012—2013��ȣ��������´����������������37.5%���ڻ��½϶��2013-2014�������18.2%������ǰ5 d���졣�����紺ũ����Ը�Ĥ��������Ч���С���������������ͷ�������ǰ���ӳ������������������ۻ���ʱ���Ǹ���С������������غͻ�ø߲���ԭ��</p>
DOI:10.3724/SP.J.1006.2016.01530URLMagsci [��������: 2]
<p>��Ի���ƽԭ���������¶ȱ仯�붬С�������������������¶ȼ��ì�ܣ��Լ��ɴ���ɵĶ�С����ԵͲ����⣬��2012—2013��2013—2014����������������ͨ�����������о��������紺����ҽ�����������¶ȶ�С�������Ӱ�졣���������2��20��ǰ��������Ĥ����ÿ��ǰ1�ܻ������23.0~49.7�棬1����Ѯ��3����Ѯ���������½��ڿ����ӻ���167.7~176.8�棬�Ӷ�С������������ǰ����Ĥ�ҳ����γɵ���Ե��»���ʹ�����������γ���ʱ���ӳ�4~10 d���볣����ֲ�����գ���ȣ��������´����Ŀ����ڸ����ʲ������18.8%��Ҷ���ָ�����14.7%��������������43.6%����������ʸ�ֵ�������ӳ�10 d���ϣ������������ε��ӳ��ٽ��˸����ʻ��ۺ�������ת�ƣ���Ч��������48~98��hm<sup><span>-</span></sup><sup>2</sup>��������������3.9~4.5����ǧ��������2.5~5.6 g����ȫ���������½��ٵ�2012—2013��ȣ��������´����������������37.5%���ڻ��½϶��2013-2014�������18.2%������ǰ5 d���졣�����紺ũ����Ը�Ĥ��������Ч���С���������������ͷ�������ǰ���ӳ������������������ۻ���ʱ���Ǹ���С������������غͻ�ø߲���ԭ��</p>
URLMagsci [��������: 1]
<div >Ϊ�˽�ʾԽ��ǰ�������ӶԶ�С�������������̺Ͳ�����Ӱ�죬��2010—2012������ʩ�ڽ�����Ϊ��������ģ������.�������������ʵ���¶�ֵΪ���գ�����Խ��ǰ����40��50��60 d���о�Խ��ǰ��ͬ����(≥0 ��)��С������ڡ����뷢�����̡������ںͳ�����ҶƬ����������Լ���������Ҫ�ص�Ӱ��.��������� ��ʩ��Խ��ǰ���ӻ�����Խ��ǰ�����뷢��������һ��Ӱ�죬�ν������뷢�����̺�����Ӱ�����ԣ���������ŷ������̵ļӿ�Ӱ���С�������ڴִ����������ѧ��״���첻����.Խ��ǰ�������Ӳ�����25 �������Ӱ���С���������Ӵ���60 �����뷢���������Լӿ죬����Խ�߱仯Խ����.��ǰ�������ӵ�һ�����Ƚ����¶�С���������ǰ���������ӳ���60 �棬�ν���Ҷ����ߪ�0.8�����ϣ������ںͳ����ڷֱ���ǰ1 d����.����ڵ���ǰ�����뷢�����̵ļӿ�ʹС���������������̣��������ܴ�������Σ�������С����������С�붳������ǰ���¹������º�����Ҷ��������½����ཬ�����̣�����ɼ���.</div><div ></br> </div>
URLMagsci [��������: 1]
<div >Ϊ�˽�ʾԽ��ǰ�������ӶԶ�С�������������̺Ͳ�����Ӱ�죬��2010—2012������ʩ�ڽ�����Ϊ��������ģ������.�������������ʵ���¶�ֵΪ���գ�����Խ��ǰ����40��50��60 d���о�Խ��ǰ��ͬ����(≥0 ��)��С������ڡ����뷢�����̡������ںͳ�����ҶƬ����������Լ���������Ҫ�ص�Ӱ��.��������� ��ʩ��Խ��ǰ���ӻ�����Խ��ǰ�����뷢��������һ��Ӱ�죬�ν������뷢�����̺�����Ӱ�����ԣ���������ŷ������̵ļӿ�Ӱ���С�������ڴִ����������ѧ��״���첻����.Խ��ǰ�������Ӳ�����25 �������Ӱ���С���������Ӵ���60 �����뷢���������Լӿ죬����Խ�߱仯Խ����.��ǰ�������ӵ�һ�����Ƚ����¶�С���������ǰ���������ӳ���60 �棬�ν���Ҷ����ߪ�0.8�����ϣ������ںͳ����ڷֱ���ǰ1 d����.����ڵ���ǰ�����뷢�����̵ļӿ�ʹС���������������̣��������ܴ�������Σ�������С����������С�붳������ǰ���¹������º�����Ҷ��������½����ཬ�����̣�����ɼ���.</div><div ></br> </div>
[��������: 1]
[��������: 1]
URL [��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
URLMagsci [��������: 1]
<P><FONT face=Verdana>��Ŀ�ġ�̽��ҹ�����¶Զ�С���������������Ӱ�켰��Ӱ����ơ����������Զ�С����-626(����)Ϊ�Բ�,���ú��������ģ����������µ�ҹ�����»���(���´�4��3��С��������ʼ,��6��15��С���ո����),�о�ҹ�����¶Զ�С���������������乹�ɵ�Ӱ�졣����������������������ʩ�����ȶ�,ʹС��ڲ�����2.5�档ҹ�����µ��¶�С������������,���������ǰ,�����������͡�ҹ�����µ��¶�С����������ǰ4 d,��������ǰ��5 d,�������������ǰ1��2 d,�ཬ��������5 d;����ҹ��ڲ�����2.5�浼�¶�С����Ч������������,С��������������ǧ����������С,��С�������26.6%�������ۡ�����ҹ�����¿ɵ��¶�С�����ȼ����������������Ƕȷ����ó�,�������ǰʹ���������̡��ཬ��������ʹǧ�����½���ҹ�����µ��¶�С���������Ҫԭ��;�Ӳ������ɷ����ó�,������������Ӧ��������Ч������������ӡ�����������ȼ����Dz������͵�����ҪӰ�����ӡ�ͬʱ,���µ�����ɢ����,����ˮ�ֽ��Ϳ����ǵ��²����½�����һ���ӡ�������ȫ���ů������,��ȡ���˴�ʩӦ��ҹ����������,��ȷ����С���Ȳ��߲���<BR></FONT></P>
URLMagsci [��������: 1]
<P><FONT face=Verdana>��Ŀ�ġ�̽��ҹ�����¶Զ�С���������������Ӱ�켰��Ӱ����ơ����������Զ�С����-626(����)Ϊ�Բ�,���ú��������ģ����������µ�ҹ�����»���(���´�4��3��С��������ʼ,��6��15��С���ո����),�о�ҹ�����¶Զ�С���������������乹�ɵ�Ӱ�졣����������������������ʩ�����ȶ�,ʹС��ڲ�����2.5�档ҹ�����µ��¶�С������������,���������ǰ,�����������͡�ҹ�����µ��¶�С����������ǰ4 d,��������ǰ��5 d,�������������ǰ1��2 d,�ཬ��������5 d;����ҹ��ڲ�����2.5�浼�¶�С����Ч������������,С��������������ǧ����������С,��С�������26.6%�������ۡ�����ҹ�����¿ɵ��¶�С�����ȼ����������������Ƕȷ����ó�,�������ǰʹ���������̡��ཬ��������ʹǧ�����½���ҹ�����µ��¶�С���������Ҫԭ��;�Ӳ������ɷ����ó�,������������Ӧ��������Ч������������ӡ�����������ȼ����Dz������͵�����ҪӰ�����ӡ�ͬʱ,���µ�����ɢ����,����ˮ�ֽ��Ϳ����ǵ��²����½�����һ���ӡ�������ȫ���ů������,��ȡ���˴�ʩӦ��ҹ����������,��ȷ����С���Ȳ��߲���<BR></FONT></P>
DOI:10.3724/SP.J.1006.2009.00110URLMagsci [��������: 1]
<div><span >Ϊ�о��ӽ����Կ�����ʵ�ڸ���в�ȵ�������̬��Ӧ�ԣ�ѡ�þߴ����Ե������ӽ�����Ϲ���</span><span >6</span><span >�š�Э��</span><span >46</span><span >���˹���Ƽ�ֵ����</span><span >40~42</span><span >��</span><span >����ʼ�������˺�</span><span >15 d</span><span >ÿ��</span><span >6 h</span><span >���Ⱥ�����������Ȼ��������Ϊ���գ��Ƚ��о�����в�ȶԹ���</span><span >6</span><span >����Э��</span><span >46</span><span >��ʵ������Ӱ�졣�������������</span><span >6</span><span >�ž߽ϸߵ�����С�����ЧӦ������������С���ʵͣ�����������Ҳ�͡�</span><span >����</span><span >6</span><span >��</span><span >�Կ�����ʵ�ڸ��½�Э��</span><span >46</span><span >�۸У�������С�����Ժ��Ⱥ�ָ���ϵIJ���ﵽ����ˮƽ</span><span >(</span><em><span >P</span></em><span ><0.05</span><span >)</span><span >��</span><span >��������ԭ��</span><span >, (1)</span><span >�����ڼ䵾�꽣Ҷ����ת��Ч�ʼ����Ч�ʽϸߣ����ʴ������ʽϷ</span><span >(2)</span><span >�����ڼ䵾���Ծ߽�ǿ��ϵ��ˮ������ҶƬ����ˮƽ��ά�����������ڹ���������䣻</span><span >(3)</span><span >����ཬ�ڵ����Զ��������䶥�����ƣ������ٿ�������������ʵ��ʵ����</span></div>
DOI:10.3724/SP.J.1006.2009.00110URLMagsci [��������: 1]
<div><span >Ϊ�о��ӽ����Կ�����ʵ�ڸ���в�ȵ�������̬��Ӧ�ԣ�ѡ�þߴ����Ե������ӽ�����Ϲ���</span><span >6</span><span >�š�Э��</span><span >46</span><span >���˹���Ƽ�ֵ����</span><span >40~42</span><span >��</span><span >����ʼ�������˺�</span><span >15 d</span><span >ÿ��</span><span >6 h</span><span >���Ⱥ�����������Ȼ��������Ϊ���գ��Ƚ��о�����в�ȶԹ���</span><span >6</span><span >����Э��</span><span >46</span><span >��ʵ������Ӱ�졣�������������</span><span >6</span><span >�ž߽ϸߵ�����С�����ЧӦ������������С���ʵͣ�����������Ҳ�͡�</span><span >����</span><span >6</span><span >��</span><span >�Կ�����ʵ�ڸ��½�Э��</span><span >46</span><span >�۸У�������С�����Ժ��Ⱥ�ָ���ϵIJ���ﵽ����ˮƽ</span><span >(</span><em><span >P</span></em><span ><0.05</span><span >)</span><span >��</span><span >��������ԭ��</span><span >, (1)</span><span >�����ڼ䵾�꽣Ҷ����ת��Ч�ʼ����Ч�ʽϸߣ����ʴ������ʽϷ</span><span >(2)</span><span >�����ڼ䵾���Ծ߽�ǿ��ϵ��ˮ������ҶƬ����ˮƽ��ά�����������ڹ���������䣻</span><span >(3)</span><span >����ཬ�ڵ����Զ��������䶥�����ƣ������ٿ�������������ʵ��ʵ����</span></div>
URL [��������: 1]
[��������: 1]
[��������: 1]
DOI:10.5846/stxb201402280346URLMagsci [��������: 1]
��С��ཬ�ڸ�������Լ�ҹ�������С�������һ����Ҫ��������,����ȫ�������ů,����Ƶ���ļ����¶��¼���������õ�����С������Ĵ��������Ϊ����ȷ��С��ཬ���쳣���¶Զ�С�������Ӱ��̶�,�ںӱ��̳���̬��ũҵ��������վ��Ȼ����������,�������ɿ�������������¼���,��չ�˶�С��ཬ�к��ڶ��ڸ��´���ģ�����顣�������,�ཬ�к��ڶ��ڸ���в����ʹ��С��ཬ�����½����ཬ����ʱ�����̴Ӷ�ʹ���ؽ���,��������Ӱ�����ص��������ص����ú�,���ڸ��´����ڼ�����ƽ���ڲ�����������֮����������ĸ�������ع�ϵ�����¶Զ�С���������в�̶��ɸ���ǿ�ȼ������ʱ���������ؾ���������˸���ǿ�ȼ������ʱ���ۺ����õĸ�����Ч��ʱ��֤����һ����ȫ�淴ӳ�ཬ�к��ڸ���в��Ӱ�����������
DOI:10.5846/stxb201402280346URLMagsci [��������: 1]
��С��ཬ�ڸ�������Լ�ҹ�������С�������һ����Ҫ��������,����ȫ�������ů,����Ƶ���ļ����¶��¼���������õ�����С������Ĵ��������Ϊ����ȷ��С��ཬ���쳣���¶Զ�С�������Ӱ��̶�,�ںӱ��̳���̬��ũҵ��������վ��Ȼ����������,�������ɿ�������������¼���,��չ�˶�С��ཬ�к��ڶ��ڸ��´���ģ�����顣�������,�ཬ�к��ڶ��ڸ���в����ʹ��С��ཬ�����½����ཬ����ʱ�����̴Ӷ�ʹ���ؽ���,��������Ӱ�����ص��������ص����ú�,���ڸ��´����ڼ�����ƽ���ڲ�����������֮����������ĸ�������ع�ϵ�����¶Զ�С���������в�̶��ɸ���ǿ�ȼ������ʱ���������ؾ���������˸���ǿ�ȼ������ʱ���ۺ����õĸ�����Ч��ʱ��֤����һ����ȫ�淴ӳ�ཬ�к��ڸ���в��Ӱ�����������
[��������: 1]
URLMagsci [��������: 1]
<P>��2�������Բ�ͬ�Ķ�С��Ʒ��ɽũ1391����8901Ϊ���ϣ������ϱ�Ĥ�Ƴ�������ֱ��ڻ���8~10 d (T1)�ͻ���15~17 d (T2)���и��´������о��˻���ͬʱ�ڸ���в�ȶ�С����Ҷ����ȩ(MDA)���������������绯ø(SOD)����������ø(CAT)��������ø(POD)���Ե�Ӱ�졣���������T1��������Ҷ�ڻ���������ϵͳ�ܸ���Ч��������ʹĤ֬������ˮƽ�����½�����T2������ɵ��˺����ɻָ�������28 d������8901��ȣ�ɽũ1391������MDA�����ϵ͡�T1������SOD��CAT����������������CAT���Ե�����������SOD������Эͬ�𱣻����ã��ӻ���Ҷ��˥�ϣ�����ɽũ1391SOD���Ե��������Ƚϴ���CAT���ԵĻָ������ӵ�������ǿ��T2������SOD����δ�����������������½���CAT������Ȼ���ڶ��գ�ȴδ����Ч����Ĥ֬�������̶ȣ�����SOD��ø����ϵͳ����������á�T1����������POD���H<SUB>2</SUB>O<SUB>2</SUB>��������T2������POD���Դ�����������ܶ�ҶƬ�����˺����ã���������Ҷ��˥�ϡ���ˣ�С����������滨���������̶��½���Ӧ�ر��ǿ�ཬ�к��ڵ���������������µ��˺���</P>
URLMagsci [��������: 1]
<P>��2�������Բ�ͬ�Ķ�С��Ʒ��ɽũ1391����8901Ϊ���ϣ������ϱ�Ĥ�Ƴ�������ֱ��ڻ���8~10 d (T1)�ͻ���15~17 d (T2)���и��´������о��˻���ͬʱ�ڸ���в�ȶ�С����Ҷ����ȩ(MDA)���������������绯ø(SOD)����������ø(CAT)��������ø(POD)���Ե�Ӱ�졣���������T1��������Ҷ�ڻ���������ϵͳ�ܸ���Ч��������ʹĤ֬������ˮƽ�����½�����T2������ɵ��˺����ɻָ�������28 d������8901��ȣ�ɽũ1391������MDA�����ϵ͡�T1������SOD��CAT����������������CAT���Ե�����������SOD������Эͬ�𱣻����ã��ӻ���Ҷ��˥�ϣ�����ɽũ1391SOD���Ե��������Ƚϴ���CAT���ԵĻָ������ӵ�������ǿ��T2������SOD����δ�����������������½���CAT������Ȼ���ڶ��գ�ȴδ����Ч����Ĥ֬�������̶ȣ�����SOD��ø����ϵͳ����������á�T1����������POD���H<SUB>2</SUB>O<SUB>2</SUB>��������T2������POD���Դ�����������ܶ�ҶƬ�����˺����ã���������Ҷ��˥�ϡ���ˣ�С����������滨���������̶��½���Ӧ�ر��ǿ�ཬ�к��ڵ���������������µ��˺���</P>
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
URLMagsci [��������: 1]
<p>�Գ��������Ρ�����������5����������С��Ʒ��Ϊ���ϣ����ö���ش���(�粥���ʲ�)�����£���2013—2014��Կ����ڡ��ཬ�ڵ���Ҷ���ָ�ꡢҶ���غ�����Ҷ����ӫ��������вⶨ����������Щ����������������.��������� Ʒ�ּ���ӫ�����(��Ҷ������)�����������������������Ҷ���ر���ϵ����С(0.12��0.17)�����չ���Ϊ����������ָ��(<em>PI</em><sub>abs</sub>)����ϵ���ϴ�(0.32��0.39)�������������ƫ���ϵ��Ϊ0.70��0.81���粥�����£����������몫<em>PI</em><sub>abs</sub>��(�ཬ�ڡ�������)���ཬ��Ҷ���س�������ЧӦ����ཬ����Ҷ��I�����Կɱ�ӫ��ǿ��(<em>V</em><sub>i</sub>)��������ЧӦ���Ҳ�����81.1%��82.8% (2013��2014��)������3���صı����������ͬƷ�ֱ��ֳ���ͬ�IJ���ЧӦ��������仯���ƻ���һ�£�����13(���ԡ�������)��Ҷ������ʡ�Ҷ���ؼ��������ӫ������Ͳ�����������������Ʒ�֣��������粥.�粥�����¹ཬ����ҶҶ���غ�������<em>PI</em><sub>abs</sub>��������ʿ���Ϊѡ��߹�ЧС����Դ����Ҫ����ָ��.</p>
URLMagsci [��������: 1]
<p>�Գ��������Ρ�����������5����������С��Ʒ��Ϊ���ϣ����ö���ش���(�粥���ʲ�)�����£���2013—2014��Կ����ڡ��ཬ�ڵ���Ҷ���ָ�ꡢҶ���غ�����Ҷ����ӫ��������вⶨ����������Щ����������������.��������� Ʒ�ּ���ӫ�����(��Ҷ������)�����������������������Ҷ���ر���ϵ����С(0.12��0.17)�����չ���Ϊ����������ָ��(<em>PI</em><sub>abs</sub>)����ϵ���ϴ�(0.32��0.39)�������������ƫ���ϵ��Ϊ0.70��0.81���粥�����£����������몫<em>PI</em><sub>abs</sub>��(�ཬ�ڡ�������)���ཬ��Ҷ���س�������ЧӦ����ཬ����Ҷ��I�����Կɱ�ӫ��ǿ��(<em>V</em><sub>i</sub>)��������ЧӦ���Ҳ�����81.1%��82.8% (2013��2014��)������3���صı����������ͬƷ�ֱ��ֳ���ͬ�IJ���ЧӦ��������仯���ƻ���һ�£�����13(���ԡ�������)��Ҷ������ʡ�Ҷ���ؼ��������ӫ������Ͳ�����������������Ʒ�֣��������粥.�粥�����¹ཬ����ҶҶ���غ�������<em>PI</em><sub>abs</sub>��������ʿ���Ϊѡ��߹�ЧС����Դ����Ҫ����ָ��.</p>
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
URLMagsci [��������: 1]
�����˹������ҿ��£��о�����С������9�ź��н�С������12�ڹཬ���ʱ�ڣ�����19��21 d�����¶ȣ�25�桢30�桢35�桢40�棩���������ʪ��50%�������£��Խ�Ҷ������ø��Ĥ֬��������Ӱ�졣��������������¶����ߣ���Ҷ���������绯ø��SOD������������ø��POD����������ø��CAT�����Գ��½����ƣ�Ĥ֬���������MDA���������������ơ�����в�ȶ��н�С������12 SOD��CAT���Ե��˺���������С������9�ţ�������9��POD���Ե��˺���������12������12 MDA�������ʴ�������9�š�����в�ȵ���ǧ�����½���
URLMagsci [��������: 1]
�����˹������ҿ��£��о�����С������9�ź��н�С������12�ڹཬ���ʱ�ڣ�����19��21 d�����¶ȣ�25�桢30�桢35�桢40�棩���������ʪ��50%�������£��Խ�Ҷ������ø��Ĥ֬��������Ӱ�졣��������������¶����ߣ���Ҷ���������绯ø��SOD������������ø��POD����������ø��CAT�����Գ��½����ƣ�Ĥ֬���������MDA���������������ơ�����в�ȶ��н�С������12 SOD��CAT���Ե��˺���������С������9�ţ�������9��POD���Ե��˺���������12������12 MDA�������ʴ�������9�š�����в�ȵ���ǧ�����½���
DOI:10.3734/SP.J.1006.2015.00136URLMagsci [��������: 1]
<p><span >Ϊ��ʾС��Ҷ���Ҷ���ٿ�����ϵͳ�Թཬ�ڸ���в�ȵķ�Ӧ������̽�ֲ�ͬƷ�ֺͲ�ͬ���������Բ�����ƣ���С��ǿ����Ʒ��ʯ��ׯ</span><span >8</span><span >�ź���������Ʒ�ֺ�ũ</span><span >341</span><span >Ϊ���ϣ��ڹཬ��������Ĥ����������´���</span><span >(</span><span >�����</span><span >8</span><span >������</span><span >22</span><span >��</span><span >)</span><span >���о�����в�ȶ���Ҷ�������</span><span >(<em>P</em><sub>n</sub>)</span><span >��Ҷ���غ�������Ҷ�ͷ�Ҷ�����б���ȩ</span><span >(MDA)</span><span >������</span><span >(Pro)</span><span >���������������绯ø</span><span >(SOD)</span><span >����������ø</span><span >(CAT)</span><span >��������ø</span><span >(POD)</span><span >���Ե�Ӱ�졣���´����£���Ʒ��</span><em><span >P</span></em><sub><span >n</span></sub><span >�������¶���</span><span >(</span><span >����</span><span >)</span><span >��</span><span >18.7%~24.9%</span><span >��Ҷ���غ�����</span><span >5.7%~6.2%</span><span >����Ҷ����Ҷ�ʡ����½ڡ�ӱƬ��������</span><span >MDA</span><span >������</span><span >Pro</span><span >���������ߣ�����</span><span >MDA</span><span >���߷���Ϊ��Ҷ</span><span >></span><span >��Ҷ���٣�</span><span >Pro</span><span >���߷���Ϊ��Ҷ����</span><span >></span><span >��Ҷ����Ҷ��ӱƬ��������</span><span >SOD</span><span >���Ժ���Ҷ����Ҷ�ʡ�������</span><span >CAT</span><span >�����Լ���Ҷ����Ҷ�ʡ�ӱƬ��</span><span >POD</span><span >�����ڸ���в�ȳ��ڼ��յ���ǿ�����������ٵĿ�����ø�������ڸ��³���һ��ʱ����յ���ǿ��֮�����Ÿ��µij��������ٿ�����ø���Զ����Ϊ���ڶ��գ����½������Ҷ�ʡ����½ڡ�ӱƬ��</span><span >SOD</span><span >���Ժ���Ҷ��ӱƬ��������</span><span >POD</span><span >�����лָ������¶��������ٵ�</span><span >SOD</span><span >��</span><span >POD</span><span >�����Լ��������ٵ�</span><span >CAT</span><span >������ɲ�����Ӱ�죻������������Ҷ���ٳ���������������������ǿ��ҶƬ��ʯ��ׯ</span><span >8</span><span >��Ҷ���Ҷ����ϸ��Ĥ�ȶ��ԡ�������ø���Ծ����ں�ũ</span><span >341</span><span >����ʾ������������ǿ�ں�ũ</span><span >341</span><span >������ʯ��ׯ</span><span >8</span><span >���ڸ���в���²����½����ȵ��ں�ũ</span><span >341</span><span >����Ҫ���������������Ϊ����Ҷ������С����Ӧ�ཬ�ڸ����澳�з�����Ҫ���á�</span></p>
DOI:10.3734/SP.J.1006.2015.00136URLMagsci [��������: 1]
<p><span >Ϊ��ʾС��Ҷ���Ҷ���ٿ�����ϵͳ�Թཬ�ڸ���в�ȵķ�Ӧ������̽�ֲ�ͬƷ�ֺͲ�ͬ���������Բ�����ƣ���С��ǿ����Ʒ��ʯ��ׯ</span><span >8</span><span >�ź���������Ʒ�ֺ�ũ</span><span >341</span><span >Ϊ���ϣ��ڹཬ��������Ĥ����������´���</span><span >(</span><span >�����</span><span >8</span><span >������</span><span >22</span><span >��</span><span >)</span><span >���о�����в�ȶ���Ҷ�������</span><span >(<em>P</em><sub>n</sub>)</span><span >��Ҷ���غ�������Ҷ�ͷ�Ҷ�����б���ȩ</span><span >(MDA)</span><span >������</span><span >(Pro)</span><span >���������������绯ø</span><span >(SOD)</span><span >����������ø</span><span >(CAT)</span><span >��������ø</span><span >(POD)</span><span >���Ե�Ӱ�졣���´����£���Ʒ��</span><em><span >P</span></em><sub><span >n</span></sub><span >�������¶���</span><span >(</span><span >����</span><span >)</span><span >��</span><span >18.7%~24.9%</span><span >��Ҷ���غ�����</span><span >5.7%~6.2%</span><span >����Ҷ����Ҷ�ʡ����½ڡ�ӱƬ��������</span><span >MDA</span><span >������</span><span >Pro</span><span >���������ߣ�����</span><span >MDA</span><span >���߷���Ϊ��Ҷ</span><span >></span><span >��Ҷ���٣�</span><span >Pro</span><span >���߷���Ϊ��Ҷ����</span><span >></span><span >��Ҷ����Ҷ��ӱƬ��������</span><span >SOD</span><span >���Ժ���Ҷ����Ҷ�ʡ�������</span><span >CAT</span><span >�����Լ���Ҷ����Ҷ�ʡ�ӱƬ��</span><span >POD</span><span >�����ڸ���в�ȳ��ڼ��յ���ǿ�����������ٵĿ�����ø�������ڸ��³���һ��ʱ����յ���ǿ��֮�����Ÿ��µij��������ٿ�����ø���Զ����Ϊ���ڶ��գ����½������Ҷ�ʡ����½ڡ�ӱƬ��</span><span >SOD</span><span >���Ժ���Ҷ��ӱƬ��������</span><span >POD</span><span >�����лָ������¶��������ٵ�</span><span >SOD</span><span >��</span><span >POD</span><span >�����Լ��������ٵ�</span><span >CAT</span><span >������ɲ�����Ӱ�죻������������Ҷ���ٳ���������������������ǿ��ҶƬ��ʯ��ׯ</span><span >8</span><span >��Ҷ���Ҷ����ϸ��Ĥ�ȶ��ԡ�������ø���Ծ����ں�ũ</span><span >341</span><span >����ʾ������������ǿ�ں�ũ</span><span >341</span><span >������ʯ��ׯ</span><span >8</span><span >���ڸ���в���²����½����ȵ��ں�ũ</span><span >341</span><span >����Ҫ���������������Ϊ����Ҷ������С����Ӧ�ཬ�ڸ����澳�з�����Ҫ���á�</span></p>
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}