 ,1, 骆强伟2, 王跃进,1
,1, 骆强伟2, 王跃进,1Breeding for Grape Germplasm Involved in Seedlessness with Cold-Resistant Using Embryo Rescue
ZHAO YaNan,1, LUO QiangWei2, WANG YueJin,1通讯作者:
责任编辑: 岳梅
收稿日期:2018-05-7接受日期:2018-07-3网络出版日期:2018-11-01
| 基金资助: |
Received:2018-05-7Accepted:2018-07-3Online:2018-11-01

摘要
关键词:
Abstract
Keywords:
PDF (2386KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
赵雅楠, 骆强伟, 王跃进. 利用胚挽救技术创制无核抗寒葡萄新种质[J]. 中国农业科学, 2018, 51(21): 4119-4130 doi:10.3864/j.issn.0578-1752.2018.21.010
ZHAO YaNan, LUO QiangWei, WANG YueJin.
0 引言
【研究意义】葡萄是世界范围内主要的果树种类之一,占世界水果栽培总面积的14.1%[1,2]。无核葡萄因在鲜食和制干上具有独特优势而深受消费者青睐,美国市场中的大部分鲜食葡萄和制干葡萄为无核品种[3,4,5]。一般利用传统杂交育种培育无核葡萄只能采用有核×无核葡萄的方式进行杂交,育种年限长,无核后代效率低。胚挽救技术可将种子败育型无核葡萄作母本,提高杂种后代无核率和育种效率。现栽培无核葡萄品种多为欧亚种(Vitis vinifera),品质优良,但抗寒性差。在中国北方地区冬季需埋土防寒,增加了成本[6],每天达到625—1 000元/hm2[7],耗时耗力。因此,利用胚挽救技术缩短育种年限,通过以无核葡萄作母本,抗寒性强的葡萄作父本,可提高选育出抗寒、优质无核葡萄的育种效率,进而减少葡萄冬季埋土防寒的损耗,具有重要价值[6]。【前人研究进展】1982年,RAMMING等[8]首次报道利用改良的White培养基培养无核葡萄胚珠,获得两株实生苗。此后,EMMERSHAD等[9,10]继续深入研究基因型、取样时期、培养基及其相互作用对无核葡萄胚珠内培养的影响,建立了辅助无核葡萄育种的胚挽救技术程序。亲本基因型的筛选与离体培养基的组成是提高育种效率的重要因素,如何提高成苗率和育种效率仍然是胚挽救育种的重要课题[11]。目前筛选出的优良品种或品系,亲本多为欧洲葡萄[4-5,12-13],突出问题是抗寒性差。抗寒性是受多数基因控制的数量性状,葡萄属植物全部19对染色体上均承担同等分量的抗寒值。我国抗寒葡萄育种始于1952年,主要以山葡萄作为欧亚种或欧美杂种导入优良品质的亲本[14]。中国野生葡萄中的一些种或株系抗寒性强,可作为抗寒无核葡萄胚挽救育种的重要种质资源。【本研究切入点】从胚挽救影响的因素出发,配置无核品种×抗寒品种的杂交组合;优化胚发育基本培养基,对杂种后代进行早期无核、抗寒性状的辅助筛选。【拟解决的关键问题】通过比较亲本基因型和对胚发育培养基的筛选与优化,提高无核葡萄品种胚挽救效率。同时将中国野生葡萄和欧山杂种葡萄抗寒性基因融入到无核品种中,筛选携带无核和抗寒标记的胚挽救苗,以期获得抗寒性强的无核新种质。1 材料与方法
试验于2017年4月至2018年4月在新疆维吾尔自治区葡萄瓜果研究所、旱区作物逆境生物学国家重点实验室和西北农林科技大学种质资源圃完成。1.1 试验材料
以种子败育型无核品种‘昆香无核’‘无核白’‘红脸无核’‘美丽无核’及‘红宝石无核’为母本。以抗寒性较强的山葡萄‘双优’(‘双庆’ב左山一’)、欧山杂种‘北冰红’(‘左优红’× 84-26-53)和‘北醇’(‘玫瑰香’ב山葡萄’)为父本。杂交组合配置详见表1。Table 1
表1
表1杂交组合配置
Table 1
| 杂交组合 Cross combination | 母本Female parent | 父本Male parent | ||
|---|---|---|---|---|
| 种或杂种 Species or hybrid | 特性 Characteristic | 种或杂种 Species or hybrid | 特性 Characteristic | |
| 昆香无核×双优 Kunxiang Seedless × Shuangyou | 欧亚种 V. vinifera | 无核、玫瑰香味 Seedless, aroma of muscat | 山葡萄 V. amurensis | 有核、抗寒 Seeded, cold-resistance |
| 无核白×双优 Thompson Seedles × Shuangyou | 欧亚种 V. vinifera | 无核 Seedless | 山葡萄 V. amurensis | 有核、抗寒 Seeded, cold-resistance |
| 红脸无核×双优 Blush Seedless × Shuangyou | 欧亚种 V. vinifera | 无核 Seedless | 山葡萄 V. amurensis | 有核、抗寒 Seeded, cold-resistance |
| 美丽无核×北冰红 Beauty Seedless × Beibinghong | 欧亚种 V. vinifera | 无核 Seedless | 欧山杂种 V. amurensis × (V. amurensis × V. vinifera) | 有核、抗寒 Seeded, cold-resistance |
| 无核白×北冰红 Thompson Seedless × Beibinghong | 欧亚种 V. vinifera | 无核 Seedless | 欧山杂种 V. amurensis × (V. amurensis × V. vinifera) | 有核、抗寒 Seeded, cold-resistance |
| 红脸无核×北冰红 Blush Seedless × Beibinghong | 欧亚种 V. vinifera | 无核 Seedless | 欧山杂种 V. amurensis × (V. amurensis × V. vinifera) | 有核、抗寒 Seeded, cold-resistance |
| 红宝石无核×北醇 Ruby Seedless × Beichun | 欧亚种 V. vinifera | 无核 Seedless | 欧山杂种 V. vinifera × V. amurensis | 有核、抗寒、抗病 Seeded, cold-resistance, disease-resistance |
新窗口打开|下载CSV
1.2 试验方法
1.2.1 胚挽救技术的基本操作流程 选择长势健壮的母本及结果母枝,对发育良好的花序去雄。去雄后套袋标记。待柱头出现透明黏液时进行人工授粉。连续授粉2—3次,每次间隔24 h。根据母本的授粉日期,采集杂交果实。胚挽救程序参照唐冬梅[1]的方法进行。将长度>2 mm的部分胚珠接种于发育培养基中,所用的发育培养基为MM3+60 g·L-1蔗糖+0.5 g·L-1水解酪蛋白+ 0.1 g·L-1肌醇+3 g·L-1活性炭+7 g·L-1琼脂,每个容器接种15粒胚珠,其余胚珠用于不同组分的发育培养基试验。暗培养10周后,无菌条件下剖取胚,接种至胚萌发培养基(WPM+20 g·L-1蔗糖+0.2 mg·L-1 6-BA+0.1 g·L-1肌醇+1 g·L-1活性炭+7 g·L-1琼脂)上。光照过程中统计不同组合发育胚的个数和正常成苗数,计算最终的胚发育率和成苗率,参考屈田田[15]的方法。
1.2.2 胚发育基本培养基对胚挽救的影响 ‘无核白’ב双优’和‘红宝石无核’ב北醇’的胚珠接种于ER或MM3培养基,比较基本培养基对胚挽救的影响。所用ER培养基和MM3培养基的主要成分参考文献[6]。每种培养基接种40—50粒胚珠,重复3次。
1.2.3 胚发育培养基中添加不同氨基酸对胚挽救的影响 以杂交组合‘昆香无核’ב双优’和‘红宝石无核’ב北醇’杂交后的胚珠接种于MM3基础培养基中,培养基内分别添加2.5 mmol·L-1的半胱氨酸、天冬酰胺、谷氨酰胺和丝氨酸,以不添加氨基酸的MM3培养基为对照。
上述培养基采用固体形式。每种处理接种30—150粒胚珠,重复3次。
1.2.4 杂交后代无核、抗寒性状的早期检测 参考文献[6]的方法,提取杂交后代株系基因组DNA,使用无核标记GLSP1-569、SCF27-2000和SCC8-1018,抗寒标记S241-717进行检测。
1.2.5 数据分析 试验数据使用SPSS 22软件进行差异显著性分析。
2 结果
2.1 无核抗寒葡萄胚挽救杂交组合成苗及基因型对胚挽救成苗的影响
2.1.1 无核抗寒葡萄胚挽救杂交组合成苗情况 2017年共配置7个杂交组合,其中1 168个胚珠接种于MM3胚发育培养基中,获得发育胚331个,成苗97株。杂交组合‘红宝石无核’ב北醇’的胚发育率和成苗率均最高,分别为46.00%和17.33%(表2)。Table 2
表2
表2杂交组合胚挽救成苗情况
Table 2
| 杂交组合 Cross combination | 授粉时间 Pollination time | 取样时间Sampling time | 接种胚珠数Number of ovules | 发育胚Embryos developed | 胚萌发成苗Plantlet formation | ||
|---|---|---|---|---|---|---|---|
| Number | % | Number | % | ||||
| 昆香无核×双优 Kunxiang Seedless × Shuangyou | 05-15 | 06-27 | 141 | 64 | 45.39 | 23 | 16.31 |
| 无核白×双优 Thompson Seedless × Shuangyou | 05-19 | 06-23 | 135 | 26 | 19.26 | 11 | 8.15 |
| 红脸无核×双优 Blush Seedless × Shuangyou | 05-17 | 07-13 | 200 | 78 | 39.00 | 19 | 9.50 |
| 美丽无核×北冰红 Beauty Seedless × Beibinghong | 05-17 | 06-23 | 203 | 21 | 10.34 | 3 | 1.48 |
| 无核白×北冰红 Thompson Seedless × Beibinghong | 05-19 | 06-23 | 157 | 12 | 7.64 | 3 | 1.91 |
| 红脸无核×北冰红 Blush Seedless × Beibinghong | 05-17 | 07-13 | 182 | 61 | 33.52 | 12 | 6.59 |
| 红宝石无核×北醇 Ruby Seedless × Beichun | 05-18 | 07-13 | 150 | 69 | 46.00 | 26 | 17.33 |
| ∑ | 1168 | 331 | — | 97 | — | ||
新窗口打开|下载CSV
2.1.2 不同母本基因型对胚挽救成苗的影响 以‘双优’为父本,‘昆香无核’‘无核白’‘红脸无核’为母本的杂交组合的胚发育率分别为45.39%、19.26%、39.00%,成苗率分别为16.31%、8.15%、9.50%,‘昆香无核’ב双优’的胚发育率和成苗率最高;以‘北冰红’为父本,‘美丽无核’‘无核白’‘红脸无核’为母本的杂交组合的胚发育率分别为10.34%、7.64%、33.52%,成苗率为1.48%、1.91%、6.59%,‘红脸无核’ב北冰红’的胚发育率和成苗率最高。结果表明,‘昆香无核’与‘红脸无核’适宜作为胚挽救母本(表2)。
2.1.3 不同父本基因型对胚挽救成苗的影响 在以‘红脸无核’为母本的两个杂交组合‘红脸无核’ב北冰红’和‘红脸无核’ב双优’中,胚发育率分别为33.52%、39.00%,成苗率分别为6.59%、9.50%;以‘无核白’为母本的杂交组合‘无核白’ב北冰红’和‘无核白’ב双优’的胚发育率分别为7.64%、19.26%,成苗率为1.91%、8.15%。因此,‘双优’要优于以‘北冰红’作为父本的胚挽救效率(表2)。
2.2 不同培养基组分对胚挽救成苗的影响
2.2.1 基本培养基对胚挽救效率的影响 ‘无核白’ב双优’和‘红宝石无核’ב北醇’的胚珠分别接种到MM3或ER基本培养基。结果表明,接种至MM3培养基所获得的胚发育率和成苗率均显著高于ER培养基(表3),说明MM3培养基更有利于胚的发育和成苗。Table 3
表3
表3不同发育培养基组分对不同杂交组合胚挽救成苗的影响
Table 3
| 杂交组合 Cross combination | 培养基 Culture medium | 胚珠数 Number of ovule | 发育胚 Embryos developed | 胚萌发成苗 Plantlet formation | |||
|---|---|---|---|---|---|---|---|
| Number | % | Number | % | ||||
| 无核白×双优 Thompson Seedless × Shuangyou | 基本培养基 Basal medium | MM3 | 135 | 26 | 19.26a | 11 | 8.15a |
| ER | 195 | 26 | 13.33b | 7 | 3.59b | ||
| 红宝石无核×北醇 Ruby Seedless × Beichun | MM3 | 150 | 69 | 46.00a | 26 | 17.33a | |
| ER | 120 | 42 | 35.00b | 11 | 9.17b | ||
| 昆香无核×双优 Kunxiang Seedless × Shuangyou | 添加不同氨基酸的培养基 Medium with different amino acids | MM3+2.5 mmol·L-1半胱氨酸Cysteine | 150 | 62 | 41.33c | 28 | 18.67ab |
| MM3+2.5 mmol·L-1天冬酰胺Asparagine | 140 | 67 | 47.86bc | 30 | 21.43a | ||
| MM3+2.5 mmol·L-1谷氨酰胺Glutamine | 129 | 77 | 59.69a | 27 | 20.93a | ||
| MM3+2.5 mmol·L-1丝氨酸Serine | 165 | 88 | 53.33ab | 22 | 13.33c | ||
| MM3 | 141 | 64 | 45.39bc | 23 | 16.31bc | ||
| 红宝石无核×北醇 Ruby Seedless × Beichun | MM3+2.5 mmol·L-1半胱氨酸Cysteine | 435 | 205 | 47.13b | 50 | 11.49c | |
| MM3+2.5 mmol·L-1天冬酰胺Asparagine | 70 | 39 | 55.71a | 15 | 21.43a | ||
| MM3+2.5 mmol·L-1谷氨酰胺Glutamine | 360 | 165 | 45.83b | 80 | 22.22a | ||
| MM3+2.5 mmol·L-1丝氨酸Serine | 340 | 151 | 44.41b | 64 | 18.82b | ||
| MM3 | 150 | 69 | 46.00b | 26 | 17.33b | ||
新窗口打开|下载CSV
2.2.2 不同氨基酸对胚挽救效率的影响 对于杂交组合‘昆香无核’ב双优’,在MM3培养基中添加2.5 mmol·L-1谷氨酰胺的胚发育率最高达59.69%,显著高于对照45.39%,说明谷氨酰胺能促进该组合胚的发育。添加天冬酰胺和丝氨酸的胚发育率(47.86%和53.33%)虽高于对照,但无显著性差异。添加天冬酰胺、谷氨酰胺培养的杂种胚珠获得的成苗率分别为21.43%、20.93%,显著高于对照16.31%,说明天冬酰胺、谷氨酰胺对该组合成苗均具有促进作用(表3)。
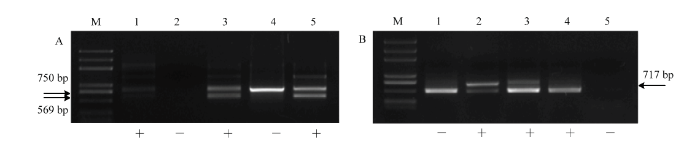
对于杂交组合‘红宝石无核’ב北醇’,添加2.5 mmol·L-1天冬酰胺的胚发育率最高达55.71%,显著高于对照46.00%,说明天冬酰胺对该组合胚发育具有促进作用。添加天冬酰胺、谷氨酰胺培养的杂种胚珠获得的成苗率分别为21.43%、22.22%,显著高于对照17.33%,说明两种氨基酸均能促进‘红宝石无核’ב北醇’的成苗。添加谷氨酰胺的‘红宝石无核’ב北醇’杂交子代成苗率最高,其成苗根系发达,叶色浓绿,植株茎端粗壮,长势较好(图1-D)。添加半胱氨酸的成苗率为11.49%,显著低于对照,对该组合的成苗具有抑制作用(表3)。
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1在胚发育培养基中添加不同氨基酸的‘红宝石无核’ב北醇’成苗
上述试验证明,不同氨基酸对两个杂交组合的胚发育和成苗影响略有差异。通过比较发现添加了酰胺类物质的培养基对两种组合的成苗均有显著促进作用,两种组合成苗率均达20%以上。
Fig. 1Plants of ‘Ruby Seedless’ × ‘Beichun’ with different amino acids added to ovule development medium
A:MM3;B—E:MM3+2.5 mmol·L-1半胱氨酸、天冬酰胺、谷氨酰胺、丝氨酸 MM3+2.5 mmol·L-1 cysteine, asparagine, glutamine, serine
2.3 杂交亲本的无核、抗寒性状检测
2.3.1 不同无核标记对杂交亲本的检测 利用3种无核标记对8个杂交亲本品种进行PCR分析的分子标记检测。无核基因探针GSLP1-569的检测结果表明,‘无核白’携带无核分子标记,其余亲本未扩增出特异条带。因此无核基因探针GSLP1-569可用于杂交组合‘无核白’ב北冰红’和‘无核白’ב双优’的杂种鉴定(图2-A);无核标记SCF27-2000的检测结果中,母本‘红脸无核’‘美丽无核’‘昆香无核’‘红宝石无核’与父本有核品种‘北冰红’‘双优’‘北醇’都具有2 000 bp特异条带,该标记不适用于对本试验杂交后代早期无核性状的鉴定(图2-B);无核标记SCC8-1018检测结果中,母本‘昆香无核’‘美丽无核’‘红脸无核’携带无核标记,适用于对组合‘昆香无核’ב双优’‘美丽无核’ב北冰红’‘红脸无核’ב北冰红’‘红脸无核’ב双优’的杂种进行早期无核性状的检测。‘红宝石无核’和父本有核品种‘北醇’携带无核标记,不适用对杂交组合‘红宝石无核’ב北醇’的杂种后代筛选(图2-C)。图2
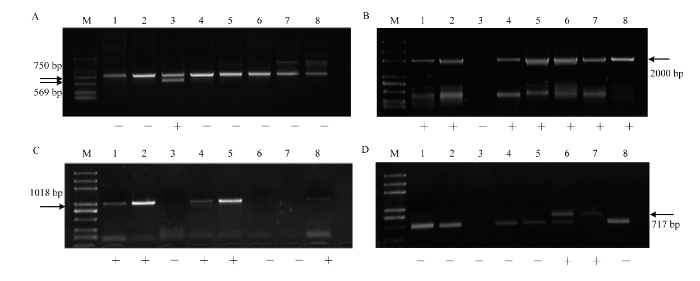 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2葡萄无核、抗寒标记对杂交亲本的扩增
A:无核探针GSLP1-569 The probe of GSLP1-569 linked to seedlessness gene;B:无核标记SCF27-2000 The molecular marker of SCF27-2000 linked to seedlessness gene;C:无核标记SCC8-1018 The molecular marker of SCC8-1018 linked to seedlessness gene;D:抗寒标记S241-717 The molecular marker of S241-717 linked to the hardiness gene;M:Marker (DL2000Plus);1:昆香无核Kunxiang Seedless;2:美丽无核Beauty Seedless;3:无核白Thompson Seedless;4:红脸无核Blush Seedless;5:红宝石无核Ruby Seedless;6:北冰红Beibinghong;7:双优Shuangyou;8:北醇Beichun;“+”表示特异条带出现,“-”表示特异条带不出现,下同 “+” indicates this material DNA contains specific band and “-” indicates this material DNA contains no specific band. The same as below
Fig. 2Amplification of the molecular marker linked to seedlessness and hardiness gene in the cross in parents
2.3.2 抗寒标记S241-717对杂交亲本的检测 抗寒标记S241-717对8个杂交亲本进行PCR扩增,父本‘北冰红’和‘双优’在717 bp处扩增出特异条带。因此可用于以‘北冰红’和‘双优’为父本的杂交组合的杂交子代早期抗寒性状初步鉴定(图2-D)。
2.4 利用不同分子标记对杂交后代无核、抗寒性状鉴定
2.4.1 无核基因探针GSLP1-569和抗寒标记S241-717对杂交后代的检测 无核探针GSLP1-569对‘无核白’ב北冰红’3个杂交后代的DNA扩增结果中,株系C-1和C-3具有569 bp的特异性条带(图3-A)。抗寒标记S241-717对‘无核白’ב北冰红’后代的扩增结果中,株系C-1和C-2扩增出717 bp特异性条带(图3-B)。同扩增出无核和抗寒特异性条带的株系为C-1(图3-A、3-B)。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3葡萄无核、抗寒标记对‘无核白’ב 北冰红’后代中的扩增
A:无核探针GSLP1-569 The probe of GSLP1-569 linked to seedlessness gene;B:抗寒标记S241-717 The molecular marker of S241-717 linked to the hardiness gene;M:Marker (DL2000Plus);1:无核白Thompson Seedless;2:北冰红Beibinghong;3:C-1;4:C-2;5:C-3
Fig. 3Amplification of the molecular marker linked to seedlessness and hardiness gene in the hybrids of ‘Thompson Seedless’ × ‘Beibinghong’
利用无核基因探针GSLP1-569对‘无核白’ב双优’16个杂交后代进行检测,其中10个株系扩增出569 bp的特异性条带(图4-A)。利用抗寒基因标记S241-717对‘无核白’ב双优’杂交后代进行检测,有9个株系扩增出特异性条带(图4-B)。筛选出同时具有无核和抗寒特异性条带的株系7个,编号为D-3、D-4、D-6、D-8、D-9、D-10、D-16(图4-A、4-B)。
图4
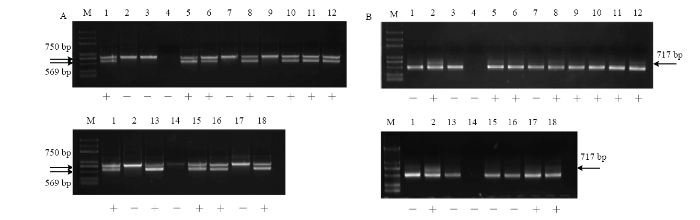 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4葡萄无核、抗寒标记对‘无核白’ב 双优’后代中的扩增
A:无核探针GSLP1-569 The probe of GSLP1-569 linked to seedlessness gene;B:抗寒标记S241-717 The molecular marker of S241-717 linked to the hardiness gene;M:Marker (DL2000Plus);1:无核白Thompson Seedless;2:双优Shuangyou;3:D-1;4:D-2;5:D-3;6:D-4;7:D-5;8:D-6;9:D-7;10:D-8;11:D-9;12:D-10;13:D-11;14:D-12;15:D-13;16:D-14;17:D-15;18:D-16
Fig. 4Amplification of the molecular marker linked to seedlessness and hardiness gene in the hybrids of ‘Thompson Seedless’ × ‘Shuangyou’
2.4.2 无核基因标记SCC8-1018和抗寒基因标记S241-717对杂交后代检测 无核标记SCC8-1018对‘红脸无核’ב北冰红’12个杂交后代进行检测结果表明,4个株系扩增出特异性条带(图5-A)。抗寒基因标记S241-717对该组合杂交后代的扩增结果中,具有特异性条带的株系7个(图5-B)。筛选出同时具有无核和抗寒特异性条带的株系3个,编号为F-5、F-6和F-10(图5-A、5-B)。
图5
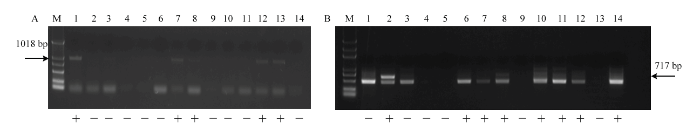 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5葡萄无核、抗寒标记对‘红脸无核’ב北冰红’后代中的扩增
A:无核标记SCC8-1018 The molecular marker of SCC8-1018 linked to seedlessness gene;B:抗寒标记S241-717 The molecular marker of S241-717 linked to the hardiness gene;M:Marker (DL2000Plus);1:红脸无核Blush Seedless;2:北冰红Beibinghong;3:F-1;4:F-2;5:F-3;6:F-4;7:F-5;8:F-6;9:F-7;10:F-8;11:F-9;12:F-10;13:F-11;14:F-12
Fig. 5Amplification of the molecular marker linked to seedlessness and hardiness gene in the hybrids ‘Blush Seedless’ × ‘Beibinghong’
无核标记SCC8-1018对‘红脸无核’ב双优’的19个杂交后代进行PCR扩增结果表明,10个株系扩增出特异性条带(图6-A)。抗寒基因标记S241-717对该组合杂交后代的扩增结果表明,15个株系具有特异性条带(图6-B)。筛选出同时具有无核和抗寒特异性条带的株系9个,编号为G-2、G-3、G-5、G-8、G-10、G-11、G-13、G-16、G-19(图6-A、6-B)。
图6
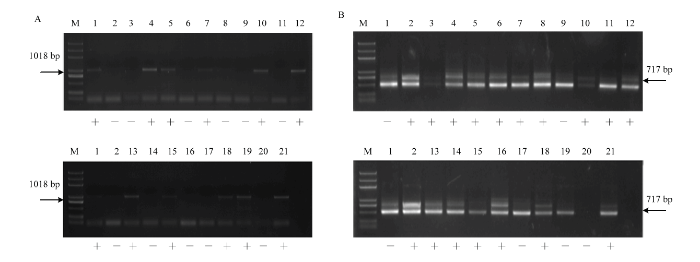 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6葡萄无核、抗寒标记对‘红脸无核’ב双优’后代中的扩增。
A:无核标记SCC8-1018 The molecular marker of SCC8-1018 linked to seedlessness gene;B:抗寒标记S241-717 The molecular marker of S241-717 linked to the hardiness gene;M:Marker (DL2000);1:红脸无核Blush Seedless;2:双优Shuangyou;3:G-1;4:G-2;5:G-3;6:G-4;7:G-5;8:G-6;9:G-7;10:G-8;11:G-9;12:G-10;13:G-11;14:G-12 15:G-13;16:G-14; 17:G-15;18:G-16;19:G-17;20:G-18;21:G-19
Fig. 6Amplification of the molecular marker linked to seedlessness and hardiness gene in the ‘ Blush Seedless’× ‘Shuangyou’
无核基因标记SCC8-1018和抗寒基因标记S241-717检测的‘美丽无核’ב北冰红’3个杂交后代均兼有无核和抗寒的特异性条带,后代得到的无核、抗寒杂交率100%(图7-A、7-B)。
图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7葡萄无核、抗寒标记对‘美丽无核’ב北冰红’后代中的扩增
A:无核标记SCC8-1018 The molecular marker of SCC8-1018 linked to seedlessness gene;B:抗寒标记S241-717 The molecular marker of S241-717 linked to the hardiness gene;M:Marker (DL2000Plus);1:美丽无核Beauty Seedless;2:北冰红Beibinghong;3:B-1;4:B-2;5:B-3
Fig. 7Amplification of the molecular marker linked to seedlessness and hardiness gene in the ‘Beauty Seedless’ × ‘Beibinghong’
无核标记SCC8-1018 对‘昆香无核’ב双优’的30个杂交后代检测结果表明,20个株系扩增出特异性条带(图8-A)。抗寒基因标记S241-717对子代的检测结果表明,19个株系具有特异性条带(图8-B)。同时筛选出具有无核和抗寒特异性条带的株系13个,编号为A-3、A-5、A-6、A-7、A-9、A-10、A-13、A-14、A-15、A-16、A-19、A-24、A-26(图8-A、8-B)。
图8
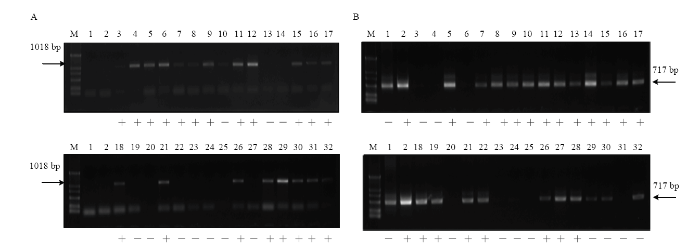 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8葡萄无核、抗寒标记对 ‘昆香无核’ב 双优’后代中的扩增
A:无核标记SCC8-1018 The molecular marker of SCC8-1018 linked to seedlessness gene;B:抗寒标记S241-717 The molecular marker of S241-717 linked to the hardiness gene;M:Marker (DL2000);1:昆香无核Kunxiang Seedless;2:双优Shuangyou;3:A-1;4:A-2;5:A-3;6:A-4;7:A-5;8:A-6;9:A-7;10:A-8;11:A-9;12:A-10;13:A-11;14:A-12;15:A-13;16:A-14;17:A-15;18:.A-16;19:A-17;20:A-18;21:A-19;22:A-20;23:A-21;24:A-22;25:A-23;26:A-24;27:A-25;28:A-26;29:A-27;30:A-28;31:A-29;32:A-30
Fig. 8Amplification of the molecular marker linked to seedlessness and hardiness gene in the ‘Kunxiang Seedless’ × ‘Shuangyou’
3 讨论
提高无核葡萄胚挽救的主要影响因素包括亲本基因型、培养基、取样时期、培养方式等。亲本间杂交的亲和力对胚挽救成苗具有重要作用[16]。本研究中杂交组合‘红宝石无核’ב北醇’获得成苗率最高,可能是由于亲本之间亲和性强,有利于合子胚的形成和萌发。杂交无核葡萄胚的发育很大程度上受到母本基因型的影响[17,18,19,20],不同种子败育型的无核葡萄中合子胚的形成能力有着较大的差异[15]。已有资料显示,胚败育发生过早或可挽救性差的品种如‘无核白’‘火焰无核’‘奇妙无核’等不宜用作母本材料[21,22],而成苗率较高的品种如‘红宝石无核’‘红脸无核’‘底莱特’等适宜作为胚挽救的母本材料[23,24]。本研究发现以‘昆香无核’‘红脸无核’为母本的杂交组合获得的成苗率高于以‘无核白’‘美丽无核’为母本的杂交组合,推测原因可能与‘昆香无核’‘红脸无核’合子胚的发育程度较高有关。欧洲葡萄抗寒性差,不利于培育抗寒性强的无核葡萄新品种[16]。利用中国野生葡萄较强的抗寒性,通过胚挽救技术将其抗寒性基因导入欧洲无核葡萄是一个有效策略。本研究父本材料山葡萄‘双优’、欧山杂种‘北冰红’‘北醇’是通过常规杂交选育出的具有强抗寒性和高商品性的品种[25,26,27]。对以‘北冰红’‘双优’为父本,相同母本杂交后获得子代的成苗率进行比较,发现‘双优’作为父本的胚挽救效率更高。说明父本基因型对胚挽救效率有一定影响,基因型差异导致亲本之间的亲和力不同。
胚发育培养基为离体胚珠提供了幼胚发育所需的激素和营养物质,直接影响到胚能否存活和进一步发育,因此胚发育培养基是胚挽救成功的关键因素之一。1994年,EMERSHED等[10]创造的ER培养基在胚珠离体培养上取得了较好效果。但潘学军[4]认为以“无核品种×中国野生葡萄”组合为材料的胚挽救不适合以ER为基本培养基,并通过测定无核葡萄品种胚败育前浆果中的大量元素含量配制出了MM3培养基,后续结果表明MM3培养基的效果更优。本研究将‘无核白’ב双优’和‘红宝石无核’ב北醇’的杂种胚珠分别接种至MM3和ER培养基中,用MM3作为胚发育培养基的胚发育率和成苗率均优于用ER培养基,结果与潘学军的结论一致[4],MM3培养基具有较高浓度的镁离子和钾离子,钾离子是植物生长发育的必要营养元素,推测提高钾离子浓度有可能会促进无核葡萄葡萄胚的发育,镁离子在植物中主要与光合作用有关,而较高浓度的镁离子可能会促进胚挽救成苗。
氨基酸的种类对胚的发育和成苗会产生不同影响。EMERSHED等研究认为谷氨酰胺、天冬酰胺、半胱氨酸、丝氨酸可促进胚的发育[9-10,13]。由于‘昆香无核’ב双优’‘红宝石无核’ב北醇’胚发育率和成苗率最高,因此用其胚珠作为氨基酸对胚挽救效率影响的试验材料。本研究用MM3作为基本培养基添加上述4种氨基酸对‘昆香无核’ב双优’‘红宝石无核’ב北醇’处理得到的结果不尽相同。谷氨酰胺促进组合‘昆香无核’ב双优’胚的发育,而天冬酰胺对组合‘红宝石无核’ב北醇’胚的发育具有促进作用,说明不同氨基酸的处理对杂交组合的作用会有差异。研究发现,在发育培养基添加天冬酰胺、谷氨酰胺后对两个组合成苗均起到促进作用,与田莉莉[20]在不同品种的研究结果类似。同时研究认为谷氨酰胺不仅提高了‘红宝石无核’ב北醇’的成苗率,也促进其成苗长势。但对于不同杂交组合胚珠的培养,需通过试验获得其最适宜的胚发育培养基和成苗途径[28]。
无核抗寒葡萄胚挽救育种过程中,通过胚挽救技术获得杂种后代后,可选用无核和抗寒的分子标记对杂交子代进行辅助选择,加快所筛选杂种进入结果期,降低育种成本。目前用于无核性状标记辅助的有SCC8-1018、SCF27-2000、GSLP1-569、VMCF7F2-198和p3-VvAGL11-1200[29,30,31,32,33]。本研究利用亲本对3种无核标记SCC8-1018、SCF27-2000、GSLP1-569进行了筛选,得到SCC8-1018可用于对‘美丽无核’ב北冰红’‘红脸无核’ב北冰红’‘红脸无核’ב双优’‘昆香无核’ב双优’4个杂交组合所获得的杂种后代进行早期无核性状的辅助选择;GSLP1-569可用于对杂交组合‘无核白’ב北冰红’‘无核白’ב双优’的杂种后代早期无核性状的辅助选择。同时应用抗寒标记S241-717对上述杂交组合所获得的后代进行了早期抗寒性状的辅助选择。经检测6个杂交组合的83个杂种后代中,存在无核特异性条带的子代为49个,抗寒特异性条带的子代为55个,同时筛选出无核和抗寒特异性条带的子代36个。初步筛选获得了36个无核抗寒新种质。
4 结论
不同的亲本基因型对胚挽救成苗的影响不同,以‘红宝石无核’‘昆香无核’‘红脸无核’作为母本和以‘双优’作为父本的杂交组合胚挽救效率较高。MM3作为基本培养基有利于幼胚的发育和成苗;MM3培养基+酰胺类氨基酸可以促进胚挽救成苗。分子标记检测的6个杂交组合83个杂种后代中,同时携带无核和抗寒分子标记的杂种后代36个,这些杂种后代是今后田间杂种鉴定与观测的重要对象,再结合葡萄抗寒性评价标准,观察无核及抗寒性状的表现,从中选出符合育种目标的优株。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[D].
[本文引用: 2]
[D].
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.3321/j.issn:1671-9387.2002.01.032URL [本文引用: 1]

对美国加州的葡萄种质资源保存与研究、栽培技术、贮藏保鲜、病虫害防治、脱毒技术以及无核葡萄品种与育种等作了较为详细的论述;同时强调指出,我国进行葡萄引种时除重视经济性状外,还应特别注意病虫检疫。
DOI:10.3321/j.issn:1671-9387.2002.01.032URL [本文引用: 1]
对美国加州的葡萄种质资源保存与研究、栽培技术、贮藏保鲜、病虫害防治、脱毒技术以及无核葡萄品种与育种等作了较为详细的论述;同时强调指出,我国进行葡萄引种时除重视经济性状外,还应特别注意病虫检疫。
[D].
[本文引用: 4]
[D].
[本文引用: 4]
[D].
[本文引用: 2]
[D].
[本文引用: 2]
[D].
[本文引用: 4]
[D].
[本文引用: 4]
URL [本文引用: 1]
北镇市农机技术推广站研制成功了100PF-A型葡萄越冬覆土机,该机是 为葡萄越冬保暖在覆盖物上覆土的专用机具,适应于沙壤土和含水率12[%]-15[%]的无石块葡萄园作业。通过试验,该机多项指标均达到设计要求,于 2003年7月通过了省级鉴定,同年8月获得国家专利。本文现将该机的主要技术参数、结构原理及机械覆土的经济效益进行了介绍,实践证明,该机结构简单, 操作方便,性能可靠,覆土厚度均匀、细碎,床面整齐,理土实,可达到防风防冻目的。
.
URL [本文引用: 1]
北镇市农机技术推广站研制成功了100PF-A型葡萄越冬覆土机,该机是 为葡萄越冬保暖在覆盖物上覆土的专用机具,适应于沙壤土和含水率12[%]-15[%]的无石块葡萄园作业。通过试验,该机多项指标均达到设计要求,于 2003年7月通过了省级鉴定,同年8月获得国家专利。本文现将该机的主要技术参数、结构原理及机械覆土的经济效益进行了介绍,实践证明,该机结构简单, 操作方便,性能可靠,覆土厚度均匀、细碎,床面整齐,理土实,可达到防风防冻目的。
[本文引用: 1]
[本文引用: 2]
DOI:10.1007/BF00233289URLPMID:24194218 [本文引用: 3]
Somatic embryo formation occurred from immature zygotic embryos within ovules of stenospermocarpic seedless grapes ( Vitis viniferaL.), when cultured for two months on liquid Emershad/Ramming...
Magsci [本文引用: 1]
<P><FONT face=Verdana>【目的】提高无核葡萄胚挽救育种效率。【方法】以12个无核葡萄品种或株系的自然授粉胚珠和6个杂交组合的胚珠为试材,研究不同配方的培养基、不同培养基相态、低温处理、光照和暗培养对无核葡萄的胚发育率的影响。同时,研究花后不同时间取样,摘取果粒取出胚珠培养的适宜时间。【结果】在供试的8种培养基中,Nitsch+GA3 0.5 mg?L-1+IAA 1.5 mg?L-1和MM3培养基最适宜于无核葡萄离体幼胚的发育,其次是ER培养基。在供试的固相、液相和固液双相的3种相态培养基中,以固液双相培养基进行无核葡萄品种底来特离体胚珠培养为最好。在胚珠离体培养阶段进行低温培养,降低了胚发育率。暗培养有利于幼胚的发育,可提高胚发育率。不同无核葡萄品种和组合在花后摘取果粒,取出胚珠进行离体胚挽救的适宜时间不同,无核白为35 d、森田尼无核为40 d、火焰无核为40 d、底来特为60 d、黎明无核为55 d、无核紫为70 d、优无核为50 d、皇家秋天为70 d、奇妙无核为60 d、奥迪亚无核为40 d、红宝石无核×贵妃玫瑰为70 d。【结论】本研究中不同无核葡萄品种进行胚挽救宜选择Nitsch+GA3 0.5 mg?L-1+IAA 1.5 mg?L-1、MM3或ER培养基,在(25±2)℃常温下进行固液双相暗培养,有助于提高胚挽救效率。<BR></FONT></P>
Magsci [本文引用: 1]
<P><FONT face=Verdana>【目的】提高无核葡萄胚挽救育种效率。【方法】以12个无核葡萄品种或株系的自然授粉胚珠和6个杂交组合的胚珠为试材,研究不同配方的培养基、不同培养基相态、低温处理、光照和暗培养对无核葡萄的胚发育率的影响。同时,研究花后不同时间取样,摘取果粒取出胚珠培养的适宜时间。【结果】在供试的8种培养基中,Nitsch+GA3 0.5 mg?L-1+IAA 1.5 mg?L-1和MM3培养基最适宜于无核葡萄离体幼胚的发育,其次是ER培养基。在供试的固相、液相和固液双相的3种相态培养基中,以固液双相培养基进行无核葡萄品种底来特离体胚珠培养为最好。在胚珠离体培养阶段进行低温培养,降低了胚发育率。暗培养有利于幼胚的发育,可提高胚发育率。不同无核葡萄品种和组合在花后摘取果粒,取出胚珠进行离体胚挽救的适宜时间不同,无核白为35 d、森田尼无核为40 d、火焰无核为40 d、底来特为60 d、黎明无核为55 d、无核紫为70 d、优无核为50 d、皇家秋天为70 d、奇妙无核为60 d、奥迪亚无核为40 d、红宝石无核×贵妃玫瑰为70 d。【结论】本研究中不同无核葡萄品种进行胚挽救宜选择Nitsch+GA3 0.5 mg?L-1+IAA 1.5 mg?L-1、MM3或ER培养基,在(25±2)℃常温下进行固液双相暗培养,有助于提高胚挽救效率。<BR></FONT></P>
[本文引用: 1]
[本文引用: 2]
DOI:10.3969/j.issn.1009-9980.2004.05.016URL [本文引用: 1]
近2个世纪,通过选种和杂交育种,世界葡萄抗寒育种在抗寒性与果实优质性状的结合方面取得了突破,育成许多抗寒性极强(耐-30~-40℃低温)、品质优良的鲜食、酿酒和制汁品种。由于葡萄的抗寒性是受多基因控制的数量性状,杂交后代的抗寒性基本呈以亲中值为峰顶的正态分布,所以我国应积极引进这些品种,在华北、西北等葡萄主产区试验、推广露地越冬栽培的同时,利用它们与现有抗寒性较弱的主栽品种(抗寒性-15~-18℃)杂交,培育适合我国冬季气候条件的抗寒品种(抗寒性在-25℃左右),迅速提高我国葡萄抗寒育种水平。
DOI:10.3969/j.issn.1009-9980.2004.05.016URL [本文引用: 1]
近2个世纪,通过选种和杂交育种,世界葡萄抗寒育种在抗寒性与果实优质性状的结合方面取得了突破,育成许多抗寒性极强(耐-30~-40℃低温)、品质优良的鲜食、酿酒和制汁品种。由于葡萄的抗寒性是受多基因控制的数量性状,杂交后代的抗寒性基本呈以亲中值为峰顶的正态分布,所以我国应积极引进这些品种,在华北、西北等葡萄主产区试验、推广露地越冬栽培的同时,利用它们与现有抗寒性较弱的主栽品种(抗寒性-15~-18℃)杂交,培育适合我国冬季气候条件的抗寒品种(抗寒性在-25℃左右),迅速提高我国葡萄抗寒育种水平。
[D].
[本文引用: 2]
[D].
[本文引用: 2]
URLMagsci [本文引用: 2]
综合分析国内外相关文献及作者以往的研究经验,阐述了无核葡萄胚挽救技术的研究现状,讨论了影响无核葡萄胚挽救成功的主要因素,包括亲本选择、取样时期、花期喷施生长调节剂、培养基、外源激素种类与浓度等,并对胚挽救技术在无核葡萄育种上的发展进行了展望。
URLMagsci [本文引用: 2]
综合分析国内外相关文献及作者以往的研究经验,阐述了无核葡萄胚挽救技术的研究现状,讨论了影响无核葡萄胚挽救成功的主要因素,包括亲本选择、取样时期、花期喷施生长调节剂、培养基、外源激素种类与浓度等,并对胚挽救技术在无核葡萄育种上的发展进行了展望。
[本文引用: 1]
[本文引用: 1]
DOI:10.3969/j.issn.1004-7360.2001.03.011URL [本文引用: 1]
本文结合作者自己的工作实际,对目前世界各国无核葡萄的育种方法及其杂交组合配置中需要注意的问题进行了简要的总结和概括.
DOI:10.3969/j.issn.1004-7360.2001.03.011URL [本文引用: 1]
本文结合作者自己的工作实际,对目前世界各国无核葡萄的育种方法及其杂交组合配置中需要注意的问题进行了简要的总结和概括.
DOI:10.17660/ActaHortic.2000.528.99URL [本文引用: 1]
Abstract Top of page Abstract 1.Introduction 2.Method 3.Results 4.Discussion 5.Conclusions Acknowledgments References Supporting Information [1] During the early Pliocene (roughly 4 Myr ago), the ocean warm water pool extended over most of the tropics. Subsequently, the warm pool gradually contracted toward the equator, while midlatitudes and subpolar regions cooled, establishing a meridional sea surface temperature (SST) gradient comparable to the modern about 2 Myr ago (as estimated on the eastern side of the Pacific). The zonal SST gradient along the equator, virtually nonexistent in the early Pliocene, reached modern values between 1 and 2 Myr ago. Here, we use an atmospheric general circulation model to investigate the relative roles of the changes in the meridional and zonal temperature gradients for the onset of glacial cycles and for Pliocene-Pleistocene climate evolution in general. We show that the increase in the meridional SST gradient reduces air temperature and increases snowfall over most of North America, both factors favorable to ice sheet inception. The impacts of changes in the zonal gradient, while also important over North America, are somewhat weaker than those caused by meridional temperature variations. The establishment of the modern meridional and zonal SST distributions leads to roughly 3.2 C and 0.6 C decreases in global mean temperature, respectively. Changes in the two gradients also have large regional consequences, including aridification of Africa (both gradients) and strengthening of the Indian monsoon (zonal gradient). Ultimately, this study suggests that the growth of Northern Hemisphere ice sheets is a result of the global cooling of Earth's climate since 4 Myr rather than its initial cause. Thus, reproducing the correct changes in the SST distribution is critical for a model to simulate the transition from the warm early Pliocene to a colder Pleistocene climate.
[D].
[本文引用: 2]
[D].
[本文引用: 2]
DOI:10.3969/j.issn.0006-3193.2004.09.007URL [本文引用: 1]
植物的胚挽救技术在无核葡萄的育种研究中发展迅速。利用假单性结实的无核葡萄作母本,无核品种作父本,杂交后在合子胚未败育时,进行胚挽救即可获得杂种实生苗,无核率可达82%,理论上可达100%。还介绍了运用该技术时杂交亲本的选择、胚挽救最佳胚龄的确定及相应的培养基成分、激素等技术方法。
DOI:10.3969/j.issn.0006-3193.2004.09.007URL [本文引用: 1]
植物的胚挽救技术在无核葡萄的育种研究中发展迅速。利用假单性结实的无核葡萄作母本,无核品种作父本,杂交后在合子胚未败育时,进行胚挽救即可获得杂种实生苗,无核率可达82%,理论上可达100%。还介绍了运用该技术时杂交亲本的选择、胚挽救最佳胚龄的确定及相应的培养基成分、激素等技术方法。
[本文引用: 1]
DOI:10.3969/j.issn.1009-9980.2008.03.006URL [本文引用: 1]
通过4a的连续试验,观察了无核葡萄果粒及胚珠发育变化,对胚挽救的多个影响因素包括品种、 接种时间、培养基和培养方式进行了研究。结果表明,红脸无核、底来特、黎明无核作母本的胚珠萌发率较高 各品种适宜的接种时间为无核白花后26 d,红脸无核花后35 d,爱莫无核×火焰无核花后48 d,底来特×红宝石无核花后56d 用Nitseh作基本培养基优于B5;添加BA 0.2 mg/L获得的胚珠萌发率最高;常温黑暗、固体培养有利于胚珠的发育。已定植结果的无核葡萄杂交胚挽救后代的无核比率平均为58.6%。将胚挽救技术用于 无核葡萄育种是十分有效的。
DOI:10.3969/j.issn.1009-9980.2008.03.006URL [本文引用: 1]
通过4a的连续试验,观察了无核葡萄果粒及胚珠发育变化,对胚挽救的多个影响因素包括品种、 接种时间、培养基和培养方式进行了研究。结果表明,红脸无核、底来特、黎明无核作母本的胚珠萌发率较高 各品种适宜的接种时间为无核白花后26 d,红脸无核花后35 d,爱莫无核×火焰无核花后48 d,底来特×红宝石无核花后56d 用Nitseh作基本培养基优于B5;添加BA 0.2 mg/L获得的胚珠萌发率最高;常温黑暗、固体培养有利于胚珠的发育。已定植结果的无核葡萄杂交胚挽救后代的无核比率平均为58.6%。将胚挽救技术用于 无核葡萄育种是十分有效的。
[本文引用: 1]
[本文引用: 1]
DOI:10.3321/j.issn:0513-353X.2008.07.024URLMagsci [本文引用: 1]
<P><FONT face=Verdana>‘北冰红’是‘左优红’与‘84-26-53’杂交育成。果实在吉林市地区9月下旬成熟。果粒圆形、蓝黑色,单果1.30g、 果穗159.5ɡ,果实含可溶性固型物18.9~25.8%,总酸1.322%~1.481%,出汁率67.1%。12月上旬采收树上冰冻果实含可溶性固型物35.2%~37.0%、总酸1.341%~1.592%、出汁率22.0%,冰红酒酒质好,抗寒性强,产量高,抗病性强。<BR></FONT></P>
DOI:10.3321/j.issn:0513-353X.2008.07.024URLMagsci [本文引用: 1]
<P><FONT face=Verdana>‘北冰红’是‘左优红’与‘84-26-53’杂交育成。果实在吉林市地区9月下旬成熟。果粒圆形、蓝黑色,单果1.30g、 果穗159.5ɡ,果实含可溶性固型物18.9~25.8%,总酸1.322%~1.481%,出汁率67.1%。12月上旬采收树上冰冻果实含可溶性固型物35.2%~37.0%、总酸1.341%~1.592%、出汁率22.0%,冰红酒酒质好,抗寒性强,产量高,抗病性强。<BR></FONT></P>
URL [本文引用: 1]
“双优”两性花山葡萄新品种选育研究皇甫淳,张辉,修荆昌,冯玉才(吉林农业大学长春130118)林兴桂,李承弘,国贤贵,钱卫友(中国农科院特产所)(集安市特产局)山葡萄抗寒性特强,色泽浓艳,酿制甜红葡萄酒,醇厚浓郁,独具一格。然而野生资源日趋枯竭,逐渐...更多还原
.
URL [本文引用: 1]
“双优”两性花山葡萄新品种选育研究皇甫淳,张辉,修荆昌,冯玉才(吉林农业大学长春130118)林兴桂,李承弘,国贤贵,钱卫友(中国农科院特产所)(集安市特产局)山葡萄抗寒性特强,色泽浓艳,酿制甜红葡萄酒,醇厚浓郁,独具一格。然而野生资源日趋枯竭,逐渐...更多还原
Magsci [本文引用: 1]
Magsci [本文引用: 1]
Magsci [本文引用: 1]
<P><FONT face=Verdana>【目的】提高无核葡萄胚挽救中幼胚的成苗率。【方法】以7个杂交组合的胚珠为试材,研究不同基因型、不同胚珠发育培养基、添加不同氨基酸及低温处理采后幼果对无核葡萄胚成苗率的影响。【结果】在无核葡萄胚离体发育中,母本和父本对胚的成苗均有影响;在二倍体无核葡萄作母本的杂交组合中,以二倍体无核葡萄作父本比以四倍体有核葡萄作父本更有利于胚的成苗。在3种不同配方的培养基中,适宜于无核葡萄离体幼胚发育和成苗的培养基是MM4。ER添加4 mmol?L-1脯氨酸培养基最有利于无核葡萄胚的发育。低温处理3 d对红宝石无核×森田尼无核和红宝石无核×黑奥林胚的成苗促进最大。【结论】无核葡萄胚挽救的适宜培养基是ER+4 mmol?L-1脯氨酸或MM4培养基,低温处理幼果3 d能有效提高二倍体无核葡萄×二倍体无核葡萄胚挽救中胚的成苗率。<BR></FONT></P>
Magsci [本文引用: 1]
<P><FONT face=Verdana>【目的】提高无核葡萄胚挽救中幼胚的成苗率。【方法】以7个杂交组合的胚珠为试材,研究不同基因型、不同胚珠发育培养基、添加不同氨基酸及低温处理采后幼果对无核葡萄胚成苗率的影响。【结果】在无核葡萄胚离体发育中,母本和父本对胚的成苗均有影响;在二倍体无核葡萄作母本的杂交组合中,以二倍体无核葡萄作父本比以四倍体有核葡萄作父本更有利于胚的成苗。在3种不同配方的培养基中,适宜于无核葡萄离体幼胚发育和成苗的培养基是MM4。ER添加4 mmol?L-1脯氨酸培养基最有利于无核葡萄胚的发育。低温处理3 d对红宝石无核×森田尼无核和红宝石无核×黑奥林胚的成苗促进最大。【结论】无核葡萄胚挽救的适宜培养基是ER+4 mmol?L-1脯氨酸或MM4培养基,低温处理幼果3 d能有效提高二倍体无核葡萄×二倍体无核葡萄胚挽救中胚的成苗率。<BR></FONT></P>
DOI:10.1007/s001220050976URL [本文引用: 1]
DOI:10.17660/ActaHortic.2003.603.74URL [本文引用: 1]
In grapevines, seedlessness or stenospermocarpy is produced due to the abortion of the embryo during the early steps of seeds development, leaving traces or rudiments almost non-detectable. At La Platina Resarch Centre (Chilean Agriculture Research Institute), a table grape breeding program using apirenic commercial varieties as progenitors and in vitro, pre-abortion embryo rescue, was established 10 to 15 years ago. Since the grapevine does not produce berries up to four or five years after the crossing, the identification of molecular markers related to seedlessness is of great interest, because they could be used to exclude seeded segregants in early stages of their development, reducing the cost of generation of new lines. For the search of this type of markers, a bulk segregant analysis (BSA) approach was choosed. In this scheme, pooled groups of plants that present or not the trait are compared, "neutralizing" the individual differences and enhancing the genetic difference(s) that could be related to the trait of interest. To build the groups, there were selected plants previously evaluated for the different stenospermocarpy sub-traits, of the crossing Ruby x Sultanina (crossing #33). Of 336 RAPD primers tested, six fragments were seedless-specific and one was related to the seeded phenotype. RAPD fragment named WF27-2000 was cloned and sequenced, and then converted into a SCAR marker (Sequence-Characterized Amplified Region). This SCAR, called SCF27, generated a specific amplicon of 2.0 kb present in all of the seedless segregants. On the other hand, the analysis of 127 plants of the crossing #33 with the SCF27 marker showed a 3:1 segregation, in agreement with both of their parents being heterozygous for this . When the marker was evaluated on the fruited segregants of crossing #33, an 81% correlation was observed. This SCAR produces an amplicon in a simple PCR reaction, their results being easily interpreted in agarose gels, which makes it usable in a marker assisted selection scheme. These results reinforce partially the theory proposed by Bouquet & Danglot (1996) about the existence of a major gene whose expression would be related to the seedlessness character. The use of the SCAR marker SCF27 in an early stage of the development of the segregants of the crossing #33 (in plantlets still maintained in vitro) would allow to reduce until one third the operational costs of the breeding program, because of the reduction in the number of plants handled in vitro, in greenhouse and in the field.
DOI:10.3321/j.issn:0513-353X.2002.02.003URLMagsci [本文引用: 1]
用葡萄无核基因的特异探针18 bp (5’CCAGTTCGCCCGTAAATG3’) 检测了我国栽培的21 个无核葡萄品种和9 个有核对照品种的无核性, 证实了该探针可以对葡萄无核基因进行准确的检测和鉴定; 通过该探针对有核×无核(红地球×红光无核) 杂交后代无核性状的鉴定, 并对照大田结果, 证明了18 bp检测葡萄无核基因探针具有预测葡萄无核性状的作用。
DOI:10.3321/j.issn:0513-353X.2002.02.003URLMagsci [本文引用: 1]
用葡萄无核基因的特异探针18 bp (5’CCAGTTCGCCCGTAAATG3’) 检测了我国栽培的21 个无核葡萄品种和9 个有核对照品种的无核性, 证实了该探针可以对葡萄无核基因进行准确的检测和鉴定; 通过该探针对有核×无核(红地球×红光无核) 杂交后代无核性状的鉴定, 并对照大田结果, 证明了18 bp检测葡萄无核基因探针具有预测葡萄无核性状的作用。
DOI:10.1139/g06-122URL [本文引用: 1]
DOI:10.1186/1471-2229-11-57URLPMID:3076230 [本文引用: 1]
pAbstract/p pBackground/p pStenospermocarpy is a mechanism through which certain genotypes of itVitis vinifera /itL. such as Sultanina produce berries with seeds reduced in size. Stenospermocarpy has not yet been characterized at the molecular level./p pResults/p pGenetic and physical maps were integrated with the public genomic sequence of itVitis vinifera /itL. to improve QTL analysis for seedlessness and berry size in experimental progeny derived from a cross of two seedless genotypes. Major QTLs co-positioning for both traits on chromosome 18 defined a 92-kb confidence interval. Functional information from model species including itVitis /itsuggested that itVvAGL11/it, included in this confidence interval, might be the main positional candidate gene responsible for seed and berry development./p pCharacterization of itVvAGL11 /itat the sequence level in the experimental progeny identified several SNPs and INDELs in both regulatory and coding regions. In association analyses performed over three seasons, these SNPs and INDELs explained up to 78% and 44% of the phenotypic variation in seed and berry weight, respectively. Moreover, genetic experiments indicated that the regulatory region has a larger effect on the phenotype than the coding region. Transcriptional analysis lent additional support to the putative role of itVvAGL11s /itregulatory region, as its expression is abolished in seedless genotypes at key stages of seed development. These results transform itVvAGL11 /itinto a functional candidate gene for further analyses based on genetic transformation./p pFor breeding purposes, intragenic markers were tested individually for marker assisted selection, and the best markers were those closest to the transcription start site./p pConclusion/p pWe propose that itVvAGL11 /itis the major functional candidate gene for seedlessness, and we provide experimental evidence suggesting that the seedless phenotype might be caused by variations in its promoter region. Current knowledge of the function of its orthologous genes, its expression profile in itVitis /itvarieties and the strong association between its sequence variation and the degree of seedlessness together indicate that the D-lineage MADS-box gene itVvAGL11 /itcorresponds to the itSeed Development Inhibitor locus /itdescribed earlier as a major locus for seedlessness. These results provide new hypotheses for further investigations of the molecular mechanisms involved in seed and berry development./p

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}