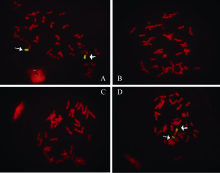
关键词:小麦; 黑麦; T1BL•1RS易位系; 赤霉病 Distribution of Wheat-Rye Translocation Line T1BL•1RS in Wheat and Its Association with Fusarium Head Blight Resistance LI Tao*, ZHENG Fei, QIN Sheng-Nan, LI Lei, GU Shi-Liang Jiangsu Provincial Key Laboratory of Crop Genetics and Physiology / Co-innovation Center for Modern Production Technology of Grain Crops / Key Laboratory of Plant Functional Genomics of Ministry of Education, Yangzhou University, Yangzhou 225009, China Fund:This study was supported by the National Major Project of Breeding for New Transgenic Organisms (2012ZX08009003-004), the National Natural Science Foundation of China (31171537), and the Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD) AbstractThe short arm of 1R chromosome (1RS) of rye carries resistant genes to stripe rust, leaf rust, stem rust, powdery mildew and aphids. To understand if 1RS also mediates resistance to wheat Fusarium head blight (FHB), we genotyped a panel of 192 wheat accessions from diverse geographic regions and a population of recombinant inbred lines (RIL) consisting of 184 lines developed from the cross of Ning 7840 and Chokwang by 1RS-specific molecular marker Xscm9 and evaluated FHB severities in three consecutive seasons in greenhouses using single floret inoculation method. The results demonstrated that 22 of 192 accessions carried a T1BL•1RS translocation, and the mean FHB severity (PSS) of varieties carrying T1BL•1RS translocation was significantly lower than that of lines without the translocation across the three experiments ( P < 0.05), indicating 1RS may have a positive effect on reducing FHB severity. 1RS-specific marker Xscm9 and Genome in situ hybridization (GISH) showed Ning 7840 carries T1BL•1RS translocation. In the population of RILs, irrespective of Fhb1 locus, the mean PSS of lines with T1BL•1RS translocation was significantly lower than that of those lines without T1BL•1RS. The effects of Fhb1 and 1RS on FHB resistance were additive and the interactions between them were not significant ( P= 0.48). The results of this study suggested that 1RS of rye most likely carries the genes resistant to FHB.
表1Fhb1和1RS组合的赤霉病扩展抗性 Table 1 Analysis of variance of 1RS and Fhb1 to type 2 resistance to Fusarium head blight
变异来源Source
SS
df
MS
F
p
Xscm9
0.857
1
0.857
19.051
1.53E-05
Xsts142
7.551
1
7.551
167.839
0
Xscm9× Xsts142
0.022
1
0.022
0.496
0.48
误差Error
23.754
528
0.045
总和Total
35.418
539
Xsts142is the closely linked marker to Fhb1 in the RIL population derived from Ning7840/Chokwang.Xscm9 is a 1RS-specific dominant marker. Xsts142是宁7840/Chokwang RIL群体中与Fhb1连锁紧密的鉴别标记; Xscm9为1RS特异分子标记。
表1Fhb1和1RS组合的赤霉病扩展抗性 Table 1 Analysis of variance of 1RS and Fhb1 to type 2 resistance to Fusarium head blight
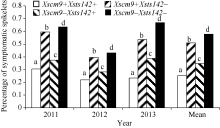
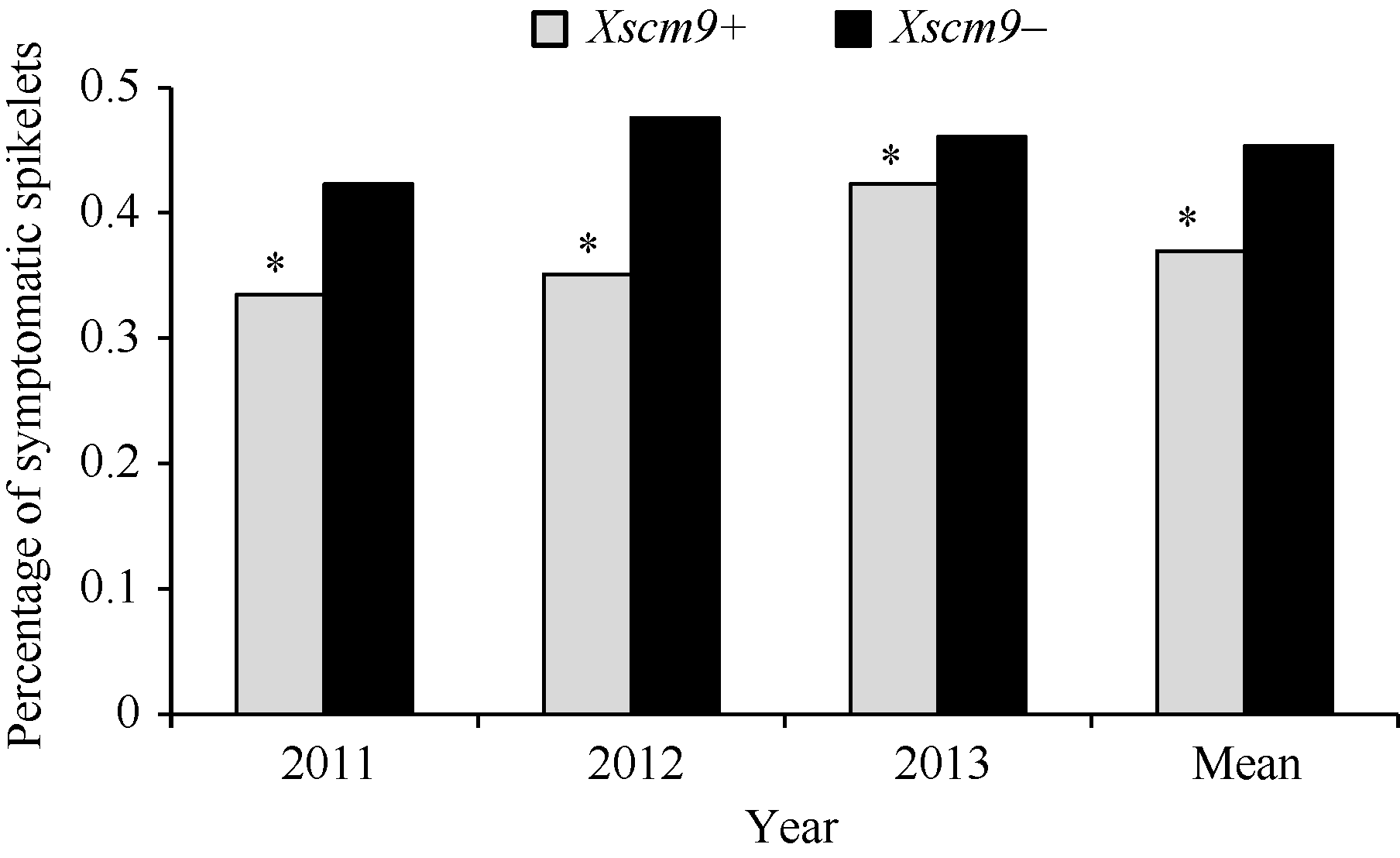
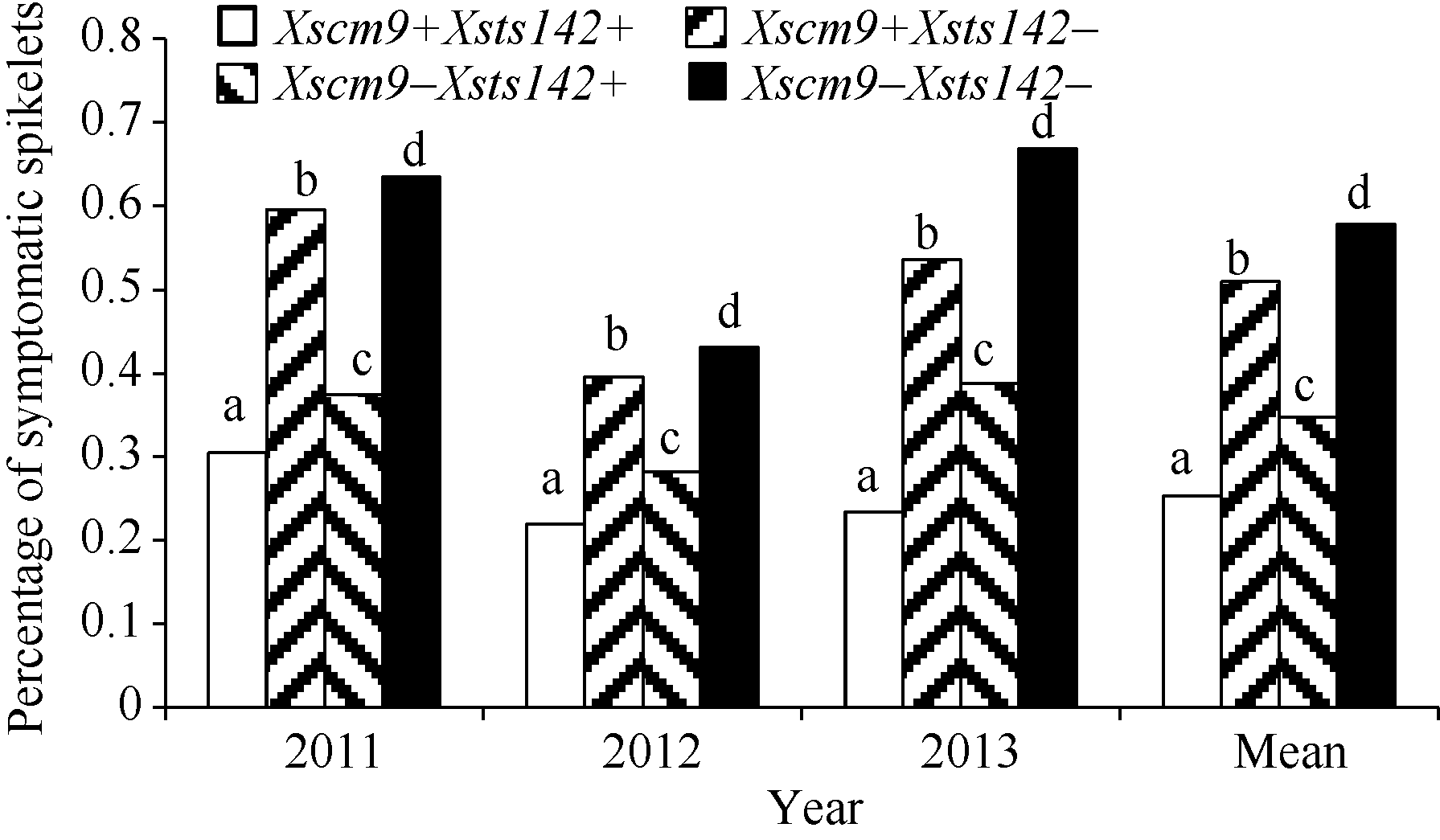
表2Fhb1、1RS不同组合型品种的赤霉病病小穗率 Table 2 Rate of symptomatic spikelets of combinations contrasting in Fhb1 and 1RS
组合 Combination
频率 Frequency
病小穗率 Rate of symptomatic spikelets
平均 Mean
2011
2012
2013
Xscm9+Xsts142+
0.194
0.305
0.220
0.234
0.253
Xscm9+Xsts142-
0.189
0.596
0.396
0.536
0.510
Xscm9-Xsts142+
0.317
0.374
0.282
0.388
0.348
Xscm9-Xsts142-
0.300
0.635
0.431
0.669
0.578
Xsts142is the closely linked marker to Fhb1 in the RIL population derived from Ning 7840/Chokwang.Xscm9 is a 1RS-specific dominant marker. Xsts142是宁7840/Chokwang RIL群体中与Fhb1连锁紧密的鉴别标记; Xscm9为1RS特异分子标记。
表2Fhb1、1RS不同组合型品种的赤霉病病小穗率 Table 2 Rate of symptomatic spikelets of combinations contrasting in Fhb1 and 1RS
RabinovichS. Importance of wheat-rye translocations for breeding modern cultivar of Triticum aestivum L. Euphytica, 1998, 100: 323-340[本文引用:1]
[2]
MagoR, MiahH, Lawrence GJ, Wellings CR, SpielmeyerW, Bariana HS, McIntosh R A, Pryor A J, Ellis J G. High-resolution mapping and mutation analysis separate the rust resistance genes Sr31, Lr26 and Yr9 on the short arm of rye chromosome 1. Theor Appl Genet, 2005, 112: 41-50[本文引用:1]
[3]
SinghN, ShepherdK, McIntosh R. Linkage mapping of genes for resistance to leaf, stem and stripe rusts and ω-secalins on the short arm of rye chromosome 1R. Theor Appl Genet, 1990, 80: 609-616[本文引用:1]
[4]
Anderson GR, PapaD, PengJ, TahirM, Lapitan NL. Genetic mapping of Dn7, a rye gene conferring resistance to the Russian wheat aphid in wheat. Theor Appl Genet, 2003, 107: 1297-1303[本文引用:1]
[5]
PengJ, WangH, Haley SD, Peairs FB, Lapitan N L V. Molecular mapping of the Russian wheat aphid resistance gene Dn2414 in wheat. Crop Sci, 2007, 47: 2418-2429[本文引用:1]
[6]
LuH, Rudd JC, Burd JD, WengY. Molecular mapping of greenbug resistance genes Gb2 and Gb6 in T1AL•1RS wheat-rye translocations. Plant Breed, 2010, 129: 472-476[本文引用:1]
[7]
MaterY, BaenzigerS, GillK, GrayboschR, WhitcherL, BakerC, SpechtJ, DweikatI. Linkage mapping of powdery mildew and greenbug resistance genes on recombinant 1RS from ‘Amigo’ and ‘Kavkaz’ wheat-rye translocations of chromosome 1RS•1AL. Genome, 2004, 47: 292-298[本文引用:1]
[8]
Maheepala DC, EhdaieB, Waines JG. Yield performance of wheat isolines with different dosages of the short arm of rye chromosome 1. J Agron Crop Sci, 2015, 201: 152-160[本文引用:1]
[9]
HowellT, HaleI, JankuloskiL, BonafedeM, GilbertM, DubcovskyJ. Mapping a region within the 1RS•1BL translocation in common wheat affecting grain yield and canopy water status. Theor Appl Genet, 2014, 127: 2695-2709[本文引用:1]
[10]
EhdaieB, Whitkus RW, Waines JG. Root biomass, water-use efficiency, and performance of wheat-rye translocations of chromosomes 1 and 2 in spring bread wheat ‘Pavon’. Crop Sci, 2003, 43: 710-717[本文引用:1]
[11]
Yang MY, Ren TH, Yan BJ, LiZ, Ren ZL. Diversity resistance to Puccinia striiformis f. sp. tritici in rye chromosome arm 1RS expressed in wheat. Genet Mol Res, 2014, 13: 8783-8793[本文引用:1]
[12]
刘建军, 肖永贵, 程敦公, 李豪圣, 刘丽, 宋健民, 刘爱峰, 赵振东, 何中虎. 利用揉面特性鉴定小麦1BL/1RS易位系. 作物学报, 2009, 35: 79-86Liu JJ, Xiao YG, Cheng DG, Li HS, LiuL, Song JM, Liu AF, Zhao ZD, He ZH. Identification of 1BL/1RS translocation based on Mixograph parameters in common wheat. Acta Agron Sin, 2009, 35: 79-86 (in Chinese with English abstract)[本文引用:2]
[13]
肖永贵, 阎俊, 何中虎, 张勇, 张晓科, 刘丽, 李天富, 曲延英, 夏先春. 1BL/1RS易位对小麦产量性状和白粉病抗性的影响及其QTL分析. 作物学报, 2006, 32: 1636-1641Xiao YG, YanJ, He ZH, ZhangY, Zhang XK, LiuL, Li TF, Qu YY, Xia XC. Effect of 1BL. 1RS translocation on yield traits and powdery mildew resistance in common wheat and QTL analysis. Acta Agron Sin, 2006, 32: 1636-1641 (in Chinese with English abstract)[本文引用:1]
[14]
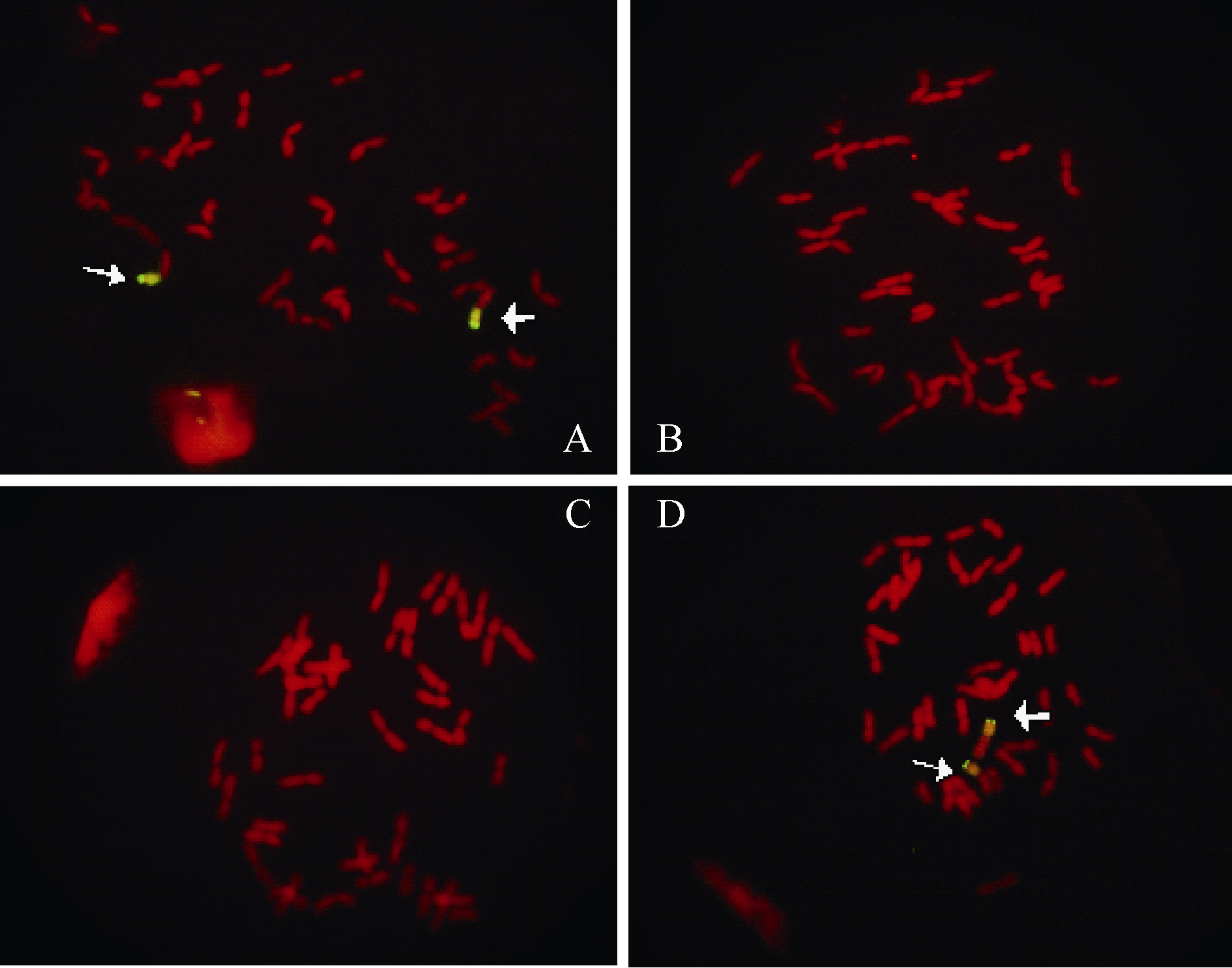
余利, 何方, 陈桂玲, 崔法, 亓晓蕾, 王洪钢, 李兴锋. 利用1RS特异标记和染色体原位杂交技术鉴定小麦1BL·1RS 易位系. 作物学报, 2011, 37: 563-569YuL, HeF, Chen GL, CuiF, Qi XL, Wang HG, Li XF. Identification of 1BL·1RS wheat-rye chromosome translocations via 1RS specific molecular markers and genomic in situ hybridization. Acta Agron Sin, 2011, 37: 563-569 (in Chinese with English abstract)[本文引用:1]
[15]
SaalB, WrickeG. Development of simple sequence repeat markers in rye (Secale cereale L. ). Genome, 1999, 42: 964-972[本文引用:2]
[16]
WengY, AzhaguvelP, Devkota RN, Rudd JC. PCR-based markers for detection of different sources of 1AL•1RS and 1BL•1RS wheat-rye translocations in wheat background. Plant Breed, 2007, 126: 482-486[本文引用:2]
[17]
BaiG, ShanerG. Management and resistance in wheat and barley to Fusarium head blight. Annu Rev Phytopathol, 2004, 42: 135-161[本文引用:1]
[18]
曾娟, 姜玉英. 2012年我国小麦赤霉病暴发原因分析及持续监控与治理对策. 中国植保导刊, 2013, 33(4): 38-41ZengJ, Jiang YY. The causal factors for the epidemics of wheat Fusarium head blight in the year of 2012 in China and the strategies for continuous monitoring and prevention. China Plant Prot, 2013, 33(4): 38-41 (in Chinese)[本文引用:1]
[19]
BuerstmayrH, BanT, Anderson JA. QTL mapping and marker-assisted selection for Fusarium head blight resistance in wheat: a review. Plant Breed, 2009, 128: 1-26[本文引用:3]
[20]
DraegerR, GosmanN, SteedA, Chand lerE, ThomsettM, SchondelmaierJ, BuerstmayrH, LemmensM, SchmolkeM, MesterhazyA. Identification of QTLs for resistance to Fusarium head blight, DON accumulation and associated traits in the winter wheat variety Arina. Theor Appl Genet, 2007, 115: 617-625[本文引用:1]
[21]
Cuthbert PA, Somers DJ, Brulé-BabelA. Mapping of Fhb2 on chromosome 6BS: a gene controlling Fusarium head blight field resistance in bread wheat (Triticum aestivum L. ). Theor Appl Genet, 2007, 114: 429-437[本文引用:1]
[22]
LiuS, ZhangX, Pumphrey MO, Stack RW, Gill BS, Anderson JA. Complex microcolinearity among wheat, rice, and barley revealed by fine mapping of the genomic region harboring a major QTL for resistance to Fusarium head blight in wheat. Funct Integr Genomic, 2006, 6: 83-89[本文引用:1]
[23]
MaH, ZhangK, GaoL, BaiG, ChenH, CaiZ, LuW. Quantitative trait loci for resistance to Fusarium head blight and deoxynivalenol accumulation in Wangshuibai wheat under field conditions. Plant Pathol, 2006, 55: 739-745[本文引用:1]
[24]
ZengJ, CaoW, HuclP, YangY, XueA, ChiD, FedakG. Molecular cytogenetic analysis of wheat-Elymus repens introgression lines with resistance to Fusarium head blight. Genome, 2013, 56: 75-82[本文引用:1]
[25]
Turner MK, DeHaan L R, Jin Y, Anderson J A. Wheatgrass- wheat partial amphiploids as a novel source of stem rust and Fusarium head blight resistance. Crop Sci, 2013, 53: 1994-2005[本文引用:1]
[26]
ZhangX, ShenX, HaoY, CaiJ, Ohm HW, KongL. A genetic map of Lophopyrum ponticum chromosome 7E, harboring resistance genes to Fusarium head blight and leaf rust. Theor Appl Genet, 2011, 122: 263-270[本文引用:1]
[27]
Yu JB, Bai GH, Cai SB, BanT. Marker-assisted characterization of Asian wheat lines for resistance to Fusarium head blight. Theor Appl Genet, 2006, 113: 308-320[本文引用:2]
[28]
PorebskiS, Bailey LG, Baum BR. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol Biol Rep, 1997, 15: 8-15[本文引用:1]
[29]
GustafsonJ, DilleJ. Chromosome location of Oryza sativa recombination linkage groups. Proc Natl Acad Sci USA, 1992, 89: 8646-8650[本文引用:1]
[30]
LiuS, ZhangX, Pumphrey MO, Stack RW, Gill BS, Anderson JA. Complex microcolinearity among wheat, rice, and barley revealed by fine mapping of the genomic region harboring a major QTL for resistance to Fusarium head blight in wheat. Funct Integr Genomic, 2006, 6: 83-89[本文引用:1]
[31]
KimW, Johnson JW, Baenziger PS, Lukaszewski AJ, Gaines CS. Agronomic effect of wheat-rye translocation carrying rye chromatin (1R) from different sources. Crop Sci, 2004, 44: 1254-1258[本文引用:1]
 , 郑飞, 秦胜男, 李磊, 顾世梁
, 郑飞, 秦胜男, 李磊, 顾世梁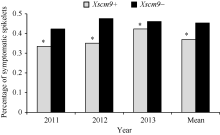

{kind=link}
{kind=link}
{kind=link}