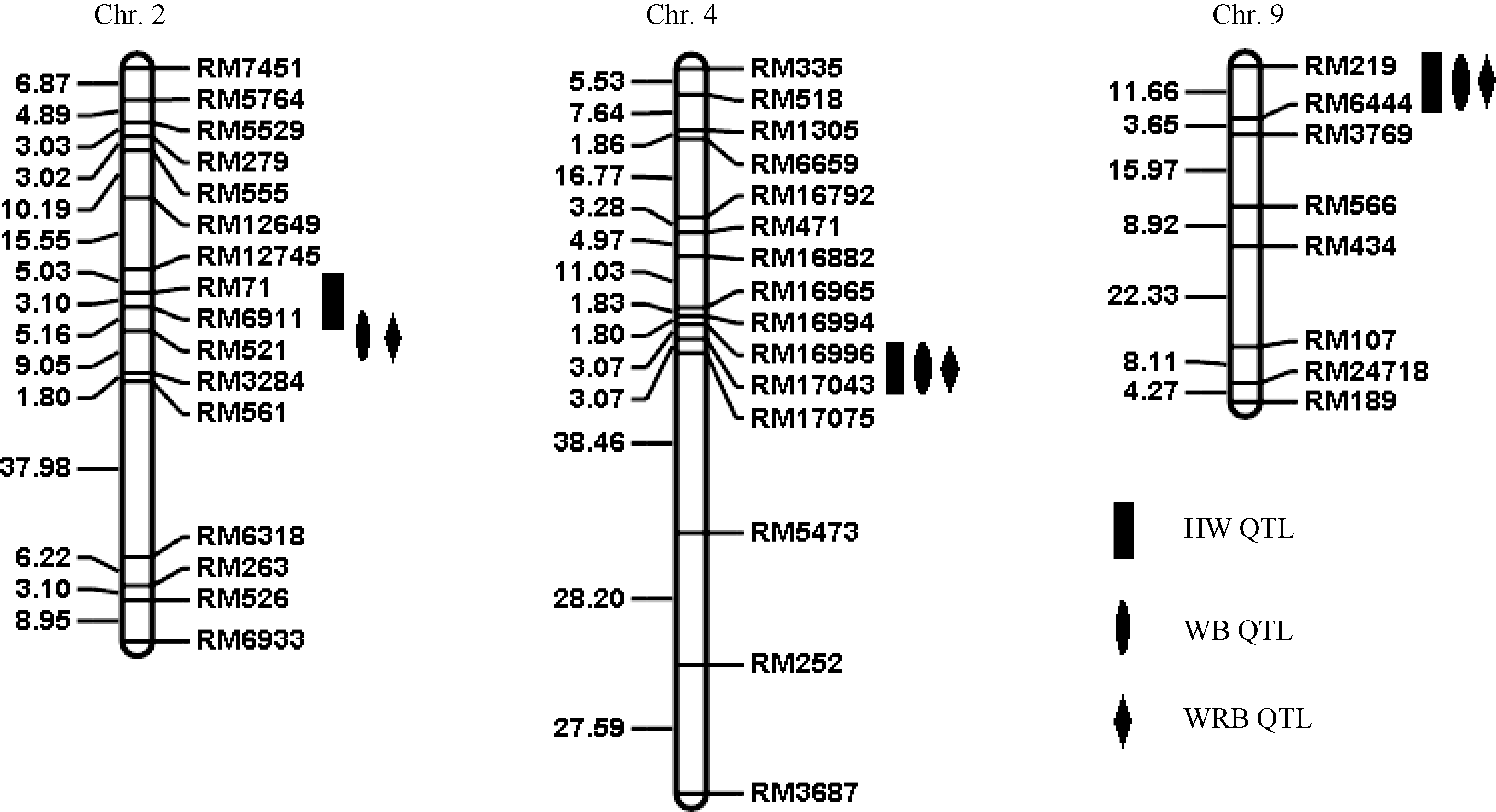
关键词:水稻; 抗性; 褐飞虱; 数量性状基因座; 遗传图谱 Analysis of QTLs for Brown Planthopper Resistance in Indica Rice WD15515 DENG Zhao**, SHI Shao-Jie**, WANG Hui-Ying, SHANG-GUAN Xin-Xin, LIU Bing-Fang, JING Sheng-Li, DU Bo, CHEN Rong-Zhi, ZHU Li-Li, HE Guang-Cun* State Key Laboratory of Hybrid Rice, College of Life Sciences, Wuhan University, Wuhan 430072, China Fund:This study was supported by the National Natural Science Foundation of China (31230060) and the National Basic Key Research Program (973 Program) (2013CBA01403) AbstractBrown planthopper (BPH) is one of the most destructive insect pests of rice. WD15515 is an indica germplasm highly resistant to BPH. An F2 population was developed from the cross between 9311 and WD15515. One hundred F2 plants were genotyped by using SSR markers and evaluated for BPH-resistance by measuring the honeydew weight secreted by BPH (HW), the body weight increment (WB) and the body weight increment ratio (WRB). A total of four QTLs for BPH resistance were identified. The qBph2-1, based on HW (honeydew weight), was detected between RM71 and RM6911 on chromosome 2, with LOD score of 3.68 and explaining the 11.08% of phenotypic variation. The qBph2-2, based on both WB and WRB, was mapped between RM6911 and RM521 on chromosome 2, with LOD score of 3.31 and 4.05 and explaining 7.81% and 9.38% of the phenotypic variation, respectively. The qBph4, based on HW, WB and WRB, was detected between RM16996 and RM17075 on chromosome 4, with LOD score of 11.11, 13.81, and 15.41 and explaining 44.38%, 45.24%, and 52.40% of the phenotypic variation, respectively. The qBph9, based on HW, WB, and WRB, was detected on chromosome 9 between RM219 and RM6444, with LOD score of 2.59, 4.04, and 3.63. This locus explained 10.91%, 12.39%, and 10.01% of the phenotypic variation in this population, respectively. qBph4is a major gene for BPH-resistance. This result provides the new resources for BPH-resistance breeding.
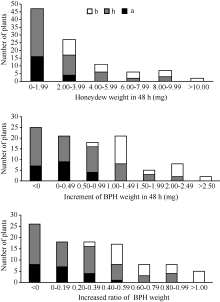
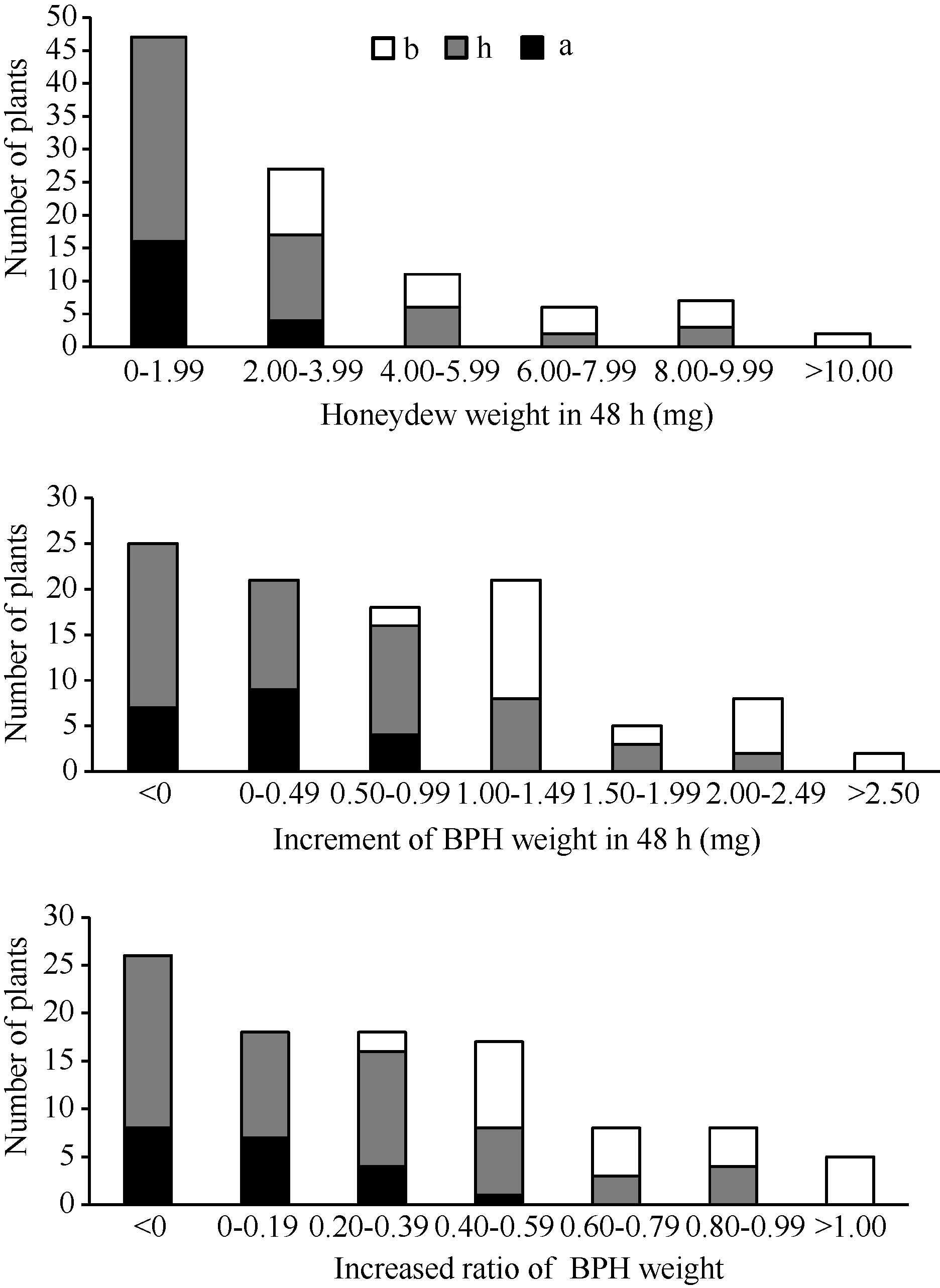
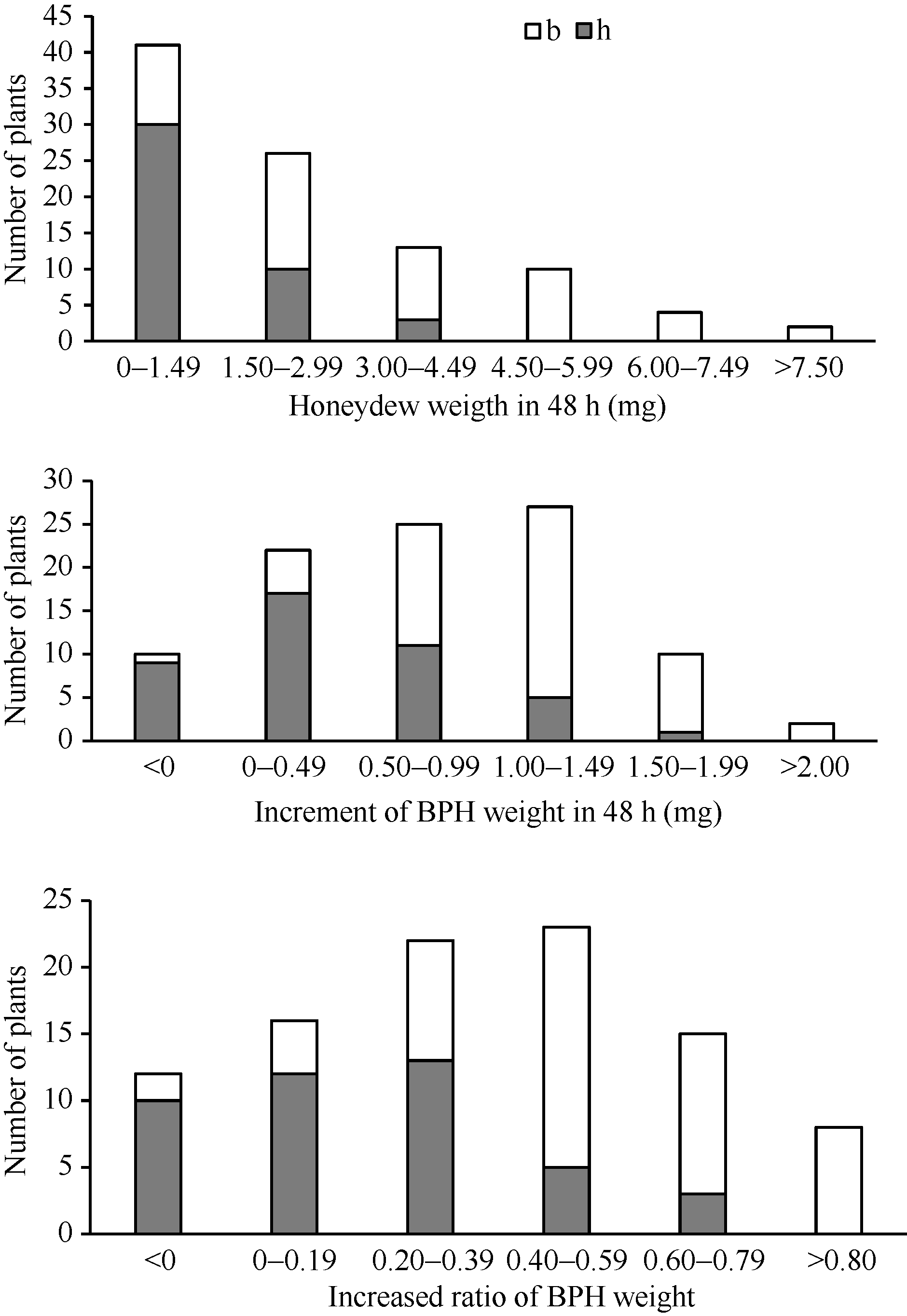
图3 9311/WD15515 F2群体的RM16996基因型和抗虫性表型频率分布图Fig. 3 Frequency distributions of BPH-resistance phenotypes and genotypes in F2population derived from the cross of 9311/WD15515
表3 Table 3 表3(Table 3)
表3 水稻9311/WD15515 F2群体中抗褐飞虱QTL检测结果 Table 3 Brown planthopper resistance QTLs detected in 9311/WD15515 F2 population
QTL
性状 Trait
染色体 Chromosome
区间 Interval
LOD值 LOD score
贡献率 PVE (%)
加性效应 ADD
显性效应 DOM
Left marker
Right marker
qBph2-1
HW
2
RM71
RM6911
3.68
11.08
-0.85
-1.12
qBph2-2
WB
2
RM6911
RM521
3.31
7.81
-0.25
-0.12
WRB
4.05
9.38
-0.13
-0.05
qBph4
HW
4
RM16996
RM17075
11.11
44.38
-2.43
-1.45
WB
13.81
45.24
-0.72
0.01
WRB
15.41
52.40
-0.33
-0.18
qBph9
HW
9
RM219
RM6444
2.59
10.91
-0.55
1.54
WB
4.04
12.39
-0.05
0.50
WRB
3.63
10.01
-0.03
0.20
HW: honeydew weight; WB: weight of BPH; WRB: weight ratio of BPH. HW: 蜜露分泌量; WB: 虫体增重; WRB: 虫体增重比。
表3 水稻9311/WD15515 F2群体中抗褐飞虱QTL检测结果 Table 3 Brown planthopper resistance QTLs detected in 9311/WD15515 F2 population
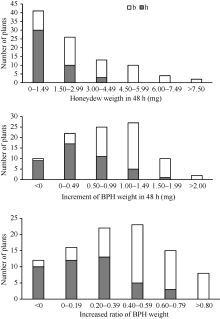
图5 WD15515/9311//9311 BC1F1群体中抗褐飞虱表型和RM16996基因型频率分布图Fig. 5 Frequency distributions of BPH-resistance phenotypes and genotypes of RM16996 in the BC1F1 population derived from the cross of WD15515/9311//9311
He GC, DuB, Chen RZ. Insect resistance. In: Zhang Q F, Wingeds R A, eds. Genetics and Genomics of Rice. New York: Springer, 2013. p 399[本文引用:1]
[2]
翟保平. 稻飞虱: 国际视野下的中国问题. 应用昆虫学报, 2011, 48: 1184-1193Zhai BP. Rice planthoppers: a China problem under the international perspectives. Chin J Appl Entomol, 2011, 48: 1184-1193 (in Chinese with English abstract). [本文引用:1]
[3]
Cheng XY, Zhu LL, He GC. Towards understand ing of molecular interactions between rice and the brown planthopper. Mol Plant, 2013, 6: 621-634[本文引用:1]
[4]
DuB, Zhang WL, Liu BF, HuJ, WeiZ, Shi ZY, He RF, Zhu LL, Chen RZ, HanB, He GC. Identification and characterization of Bph14, a gene conferring resistance to brown planthopper in rice. Proc Natl Acad Sci USA, 2009, 106: 22163-22168[本文引用:1]
[5]
TamuraY, HattoriM, YoshiokaH, YoshiokaM, TakahashiA, Wu JZ, SentokuN, YasuiH. Map-based cloning and characterization of a brown planthopper resistance gene BPH26 from Oryza sativa L. ssp. indica cultivar ADR52. Sci Rep, 2014, 4: 5872[本文引用:1]
[6]
LiuY, WuH, ChenH, WuH, ChenH, Liu YL, HeJ, Kang HY, Sun ZG, PanG, WangQ, Hu JL, ZhouF, Zhou KN, Zheng XM, Ren YL, Chen LM, Wang YH, Zhao ZG, Lin QB, Wu FQ, ZhangX, Guo XP, Cheng XN, JiangL, Wu CY, Wang HY, Wan JM. A gene cluster encoding lectin receptor kinases confers broad-spectrum and durable insect resistance in rice. Nat Biotechnol, 2015, 33: 301-305[本文引用:1]
[7]
Cohen MB, Alam SN, Medina EB, Bernal CC. Brown planthopper, Nilaparvatalugens, resistance in rice cultivar IR64: mechanism and role in successful N. lugens management in Central Luzon, Philippines. Entomol Exp Appl, 1997, 85: 221-229[本文引用:1]
[8]
HuangZ, He GC, Shu LH, Li XH, Zhang QF. Identification and mapping of two brown planthopper resistance genes in rice. Theor Appl Genet, 2001, 102: 929-934[本文引用:1]
[9]
Xu XF, Mei HW, Luo LJ, Chen XN, Li ZK. RFLP-facilitated investigation of the quantitative resistance of rice to brown planthopper (Nilaparvata lugens). Theor Appl Genet, 2002, 104: 248-253[本文引用:1]
[10]
苏昌潮, 程遐年, 翟虎渠, 万建民. 利用回交重组自交群体检测水稻抗褐飞虱数量性状基因座. 遗传学报, 2002, 29: 332-338Su CC, Cheng XN, Zhai HQ, Wan JM. Detection and analysis of QTL for resistance to the brown planthopper, Nilaparvata lugens (Stal), in rice (Oryza sativa L. ), using backcross inbred lines. Acta Genet Sin, 2002, 29: 332-338 (in Chinese with English abstract)[本文引用:1]
[11]
王建康. 数量性状基因的完备区间作图方法. 作物学报, 2009, 35: 239-245Wang JK. Inclusive composite interval mapping of quantitative trait genes. Acta Agron Sin, 2009, 35: 239-245 (in Chinese with English abstract)[本文引用:1]
[12]
李慧慧, 张鲁燕, 王建康. 数量性状基因定位研究中若干常见问题的分析与解答. 作物学报, 2010, 36: 918-931Li HH, Zhang LY, Wang JK. Analysis and answers to frequently asked questions in quantitative trait locus mapping. Acta Agron Sin, 2010, 36: 918-931 (in Chinese with English abstract). [本文引用:1]
[13]
Kosambi DD. The estimation of map distances from recombination values. Ann Eugenics, 1994, 12: 172-175[本文引用:1]
[14]
McCouch SR, Cho YG, YanoM, PaulE, BlinstrubM, MorishimaH, KinoshitaT. Report on QTL nomenclature. Rice Genet Newsl, 1997, 14(11): 11-13[本文引用:1]
[15]
McCouch SR, TeytelmanL, XuY, Lobos KB, ClareK, WaltonM, Fu BY, MaghirangR, Li ZK, Xing YZ. Development and mapping of 2240 new SSR markers for rice (Oryza sativa L. ). DNA Res, 2002, 9(6): 199-207[本文引用:1]
[16]
刘光杰, 郑宜才, 桂丽琴, 沈君辉. 水稻品种抗稻飞虱鉴定方法的比较研究. 浙江农业学报, 1999, 11: 306-310Liu GJ, Zheng YC, Gui LQ, Shen JH. A comparative study of the rice planthopper resistance identification method. Zhejiang Agric Sci, 1999, 11: 306-310 (in Chinese)[本文引用:1]
[17]
刘国庆, 颜辉煌, 傅强, 钱前, 张志涛, 翟文学, 朱立煌. 栽培稻的紧穗野生稻抗褐风虱主效基因的遗传定位. 科学通报, 2001, 46: 738-742Liu GQ, Yan HH, FuQ, QianQ, Zhang ZT, Zhai WX, Zhu LH. Mapping of a new gene for brown planthopper resistance in cultivated rice introgressed from Oryza eichingeri. Chin Sci Bull, 2001, 46: 1495-1462 (in Chinese)[本文引用:1]
[18]
Ali MP, Chowdhury TR. Tagging and mapping of genes and QTLs of Nilaparvatalugens resistance in rice. Euphytica, 2014, 195: 1-30[本文引用:1]
[19]
Qiu YF, Guo JP, Jing SL, Zhu LL, He GC. Development and characterization of japonica rice lines carrying the brown planthopper-resistance genes BPH12 and BPH6. Theor Appl Genet, 2012, 124: 485-494[本文引用:1]
[20]
Yang HY, You AQ, Yang ZF, ZhangF, He RF, Zhu LL, He GC. High-resolution genetic mapping at the Bph15 locus for brown planthopper resistance in rice (Oryza sativa L. ). Theor Appl Genet, 2004, 110: 182-191[本文引用:1]
[21]
Sun LH, Su CC, Wang CM, Zhai HQ, Wan JM. Mapping of a major resistance gene to the brown planthopper in the rice cultivar Rathu Heenati. Breed Sci, 2005, 55: 391-396[本文引用:1]
[22]
Rahman ML, JiangW, Chu SH, QiaoY, Ham TH, Woo MO, LeeJ, Khanam MS, Chin JH, Jeung JU. High-resolution mapping of two rice brown planthopper resistance genes, Bph20(t) and Bph21(t), originating from Oryza minuta. Theor Appl Genet, 2009, 119: 1237-1246[本文引用:1]
[23]
HirabayashiH, OgawaT. Identification and utilization of DNA markers linked to genes for reistance to brown planthopper (BPH) in rice. Adv Breed (Japan), 1999, 41: 71-74[本文引用:1]
[24]
Qiu YF, Guo JP, Jing SL, Zhu LL, He GC. High-resolution mapping of the brown planthopper resistance gene Bph6 in rice and characterizing its resistance in the 9311 and Nipponbare near isogenic backgrounds. Theor Appl Genet, 2010, 121: 1601-1611[本文引用:1]
[25]
Li RB, Li LS, Wei SM, Wei YP, Chen YZ, Bai DL, YangL, Huang FK, Lu WL, Zhang XJ, Li XY, Yang XQ, Wei YW. The evaluation and utilization of new genes for brown planthopper resistance in common wild rice (Oryza rufipogon Griff. ). Mol Plant Breed, 2006, 4: 365-371[本文引用:1]
[26]
HuangD, QiuY, ZhangY, HuangF, MengJ, WeiS, LiR, ChenB. Fine mapping and characterization of BPH27, a brown planthopper resistance gene from wild rice (Oryza rufipogon Griff. ). Theor Appl Genet, 2013, 126: 219-229[本文引用:1]
[27]
HeJ, LiuY, Liu YQ, Liu YL, JiangL, WuH, Kang HY, Liu SJ, Chen LM, LiuX, Cheng XN, Wan JM. High-resolution mapping of brown planthopper (BPH) resistance gene Bph27(t) in rice (Oryza sativa L. ). Mol Breed, 2013, 31: 549-557[本文引用:1]
[28]
MeiM, ZhuangC, WanR, WuJ, HuW, KochertG. Genetic analysis and tagging of gene for brown planthopper resistance in indica rice. In: Rice Genetics III. Proceedings of the Third International Rice Genetics Symposium. IRRI, Los Baños, Philippines, 1996. pp 16-20[本文引用:1]
 , 石少阶
, 石少阶




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}