 , 张溪, 张前前, 朱爽阁, 熊正琴
, 张溪, 张前前, 朱爽阁, 熊正琴
南京农业大学资源与环境科学学院, 江苏省低碳农业与温室气体减排重点实验室, 南京 210095
收稿日期: 2021-05-27; 修回日期: 2021-06-14; 录用日期: 2021-06-14
基金项目: 国家自然科学基金面上项目(No.41977078)
作者简介: 陈安枫(1997-), 男, E-mail: 2019103086@njau.edu.cn
通讯作者(责任作者): 熊正琴, E-mail: zqxiong@njau.edu.cn
摘要:氨氧化细菌(AOB)和氨氧化古菌(AOA)是驱动土壤氨氧化过程的"引擎".氨氧化过程在土壤氧化亚氮(N2O)和一氧化氮(NO)排放过程中扮演着重要角色.有机无机肥配施是实现化肥零增长和作物稳产增产的重要途径,但在有机无机肥配施下,菜地土壤AOB和AOA对氨氧化过程的相对贡献仍不清楚.本研究采用选择性抑制的方法(辛炔和乙炔)区分有机肥添加近3年后(2016年10月—2019年5月)AOB和AOA在氨氧化过程中对碱性菜地土壤N2O和NO产生的相对贡献.试验共设5种施肥处理:不施氮肥(CK)、单施尿素(N)、单施有机肥(M)、50%尿素+50%有机肥(M1N1)和80%尿素+20%有机肥(M1N4).结果表明,有机无机肥配施(M1N1和M1N4)可显著增加土壤电导率、有机碳和全氮含量.培养试验发现,与N处理相比,M和M1N1处理分别使N2O排放量增加100.7%和38.8%,NO排放量增加77.9%和42.8%,AOB基因丰度增加16.6%和10.2%,同时,AOB对N2O排放的相对贡献增加6.5%.相反,M1N4处理分别使N2O和NO排放量降低19.3%和4.8%,AOB基因丰度降低37.5%,同时,AOB对N2O及NO排放的相对贡献分别降低7.8%和7.4%.相关分析表明,土壤N2O和NO累积排放量与土壤AOB基因丰度呈显著正相关(p < 0.05),与土壤AOA基因丰度无显著相关性.有机无机肥配施下AOB是氨氧化过程的主要驱动者,适当比例的有机无机肥配施(即M1N4)措施可在一定程度上减弱AOB对碱性菜地土壤N2O及NO排放的相对贡献.
关键词:氨氧化细菌(AOB)氨氧化古菌(AOA)氧化亚氮(N2O)一氧化氮(NO)有机无机肥配施
Responses of N2O and NO emissions driven by ammonia-oxidizing bacteria and archaea to different combinations of organic and inorganic N fertilization in a vegetable field
CHEN Anfeng
, ZHANG Xi, ZHANG Qianqian, ZHU Shuangge, XIONG ZhengqinJiangsu Key Laboratory of Low Carbon Agriculture and GHGs Mitigation, College of Resources and Environmental Sciences, Nanjing Agricultural University, Nanjing 210095
Received 27 May 2021; received in revised from 14 June 2021; accepted 14 June 2021
Abstract: Ammonia-oxidizing bacteria (AOB) and ammonia-oxidizing archaea (AOA) are the engines of soil ammonia oxidation. The ammonia oxidation process plays an important role in the emissions of nitrous oxide (N2O) and nitric oxide (NO) from the soil. The combined application of organic and inorganic nitrogen (N) fertilizers is an important way to achieve zero growth of chemical N fertilizer application and stable crop yields. However, the relative contribution of AOB and AOA to ammonia oxidation in vegetable soil is still unclear. In this study, the relative contribution of AOB and AOA to N2O and NO emissions from an alkaline vegetable soil was investigated by the inhibitor technology (octyne+acetylene) after three years of combined application of organic and inorganic N fertilizers (from October 2016 to May 2019). In the field experiment, five treatments were set up as follows: no N application (CK), only urea N (N), only organic N fertilizer (M), 50% urea+50% organic N fertilizer (M1N1), and 80% urea+20% organic N fertilizer (M1N4). The combined application of organic and inorganic N fertilizers (M1N1 and M1N4) significantly increased soil EC, SOC, and TN contents. Compared with the N treatment, the M and M1N1 treatments increased N2O emissions by 100.7% and 38.8%, NO emissions by 77.9% and 42.8%, AOB gene abundance by 16.6% and 10.2%, respectively; and the average relative contribution of AOB to N2O emissions was increased by 6.5%. To the contrary, relative to the N treatment, the M1N4 treatment reduced N2O and NO emissions by 19.3% and 4.8%, respectively; reduced AOB gene abundance by 37.5%, and the relative contribution of AOB to N2O and NO emissions by 7.8% and 7.4%, respectively. Correlation analysis showed that the cumulative emissions of N2O and NO were positively correlated with the AOB gene abundance (p < 0.05), while no correlation with the AOA gene abundance. It's inferred that the AOB was the main driver of ammonia oxidation in the alkaline vegetable soil under the combined application of organic and inorganic N fertilizers, and the appropriate mixing ratio at M1N4 fertilization could reduce the contribution of AOB to N2O and NO emissions in this study.
Keywords: ammonia-oxidizing bacteria (AOB)ammonia-oxidizing archaea(AOA)nitrous oxide (N2O)nitric oxide (NO)organic and inorganic fertilization
1 引言(Introduction)目前, 中国蔬菜播种面积已突破3亿亩, 占主要农作物播种面积的13.6%, 单位面积蔬菜产量是全球的1.1倍(宁启文等, 2018).因水肥施用量大、耕作频繁及复种指数高等特点, 极易造成土壤酸化、水体富营养化和活性氮排放等环境问题(Guo et al., 2010;Zhuang et al., 2019).氧化亚氮(N2O)作为一种长效温室气体, 百年时间尺度上其增温潜势约是二氧化碳的265倍, 且会破坏大气臭氧层.相比之下, 一氧化氮(NO)虽不直接影响地球辐射强迫, 但会增加对流层产生的臭氧进而危害人体健康(Anenberg et al., 2012).通过meta分析发现, 中国菜地N2O和NO平均排放系数分别为2.4%和1.7%, 高于其他生态系统(Liu et al., 2017).
硝化过程是农业生态系统N2O和NO产生的重要路径(Mosier et al., 1998).硝化过程是指在有氧条件下, 微生物将氨(NH3)氧化, 最终生成硝态氮(NO3-)的过程.研究发现, 在高氮投入的碱性菜地土壤中, 硝化过程贡献了56%的N2O排放量(Duan et al., 2019).氨氧化过程是硝化过程的第一步, 也是限速步骤.在菜地土壤生态系统中, 氨氧化过程N2O排放量占据总排量的58%(Zhang et al., 2016).此外, 有研究发现在25 ℃和60%田间持水量条件下, 粉壤土氨氧化过程对N2O排放的贡献高达80%(G?dde et al., 1999).因此, 深入了解氨氧化过程对减少菜地活性氮排放有重要意义.
虽然最近在陆地氮循环中发现了全程硝化菌, 但通常认为氨氧化过程主要由氨氧化细菌(AOB)和氨氧化古菌(AOA)控制(Beeckman et al., 2018).AOB和AOA的丰度及群落结构受到多种环境因子的影响.在碱性高氮环境中, AOB具有更高的微生物活性, 主导氨氧化过程, 而AOA主要在酸性低氮环境下发挥功能活性(Li et al., 2018).研究发现, 施用尿素会增加AOB基因丰度, 同时提高硝化活性(Ai et al., 2013);有机肥投入会刺激AOB和AOA的生长和活化, 提高硝化活性, 从而增加农田土壤N2O排放(解开治等, 2016).相反, 有研究发现施用有机肥可降低酸性菜地AOB基因丰度, 降低硝化活性, 从而减少N2O排放(Geng et al., 2021).有机无机肥配施能够增强土壤固碳能力、改善土壤理化性质、提高肥料利用率, 同时改善生物学特性、增产增质(Charles et al., 2017; Ren et al., 2018), 有利于实现中国农业农村部于2015年提出的到2020年“一控二减三基本”的目标.尽管当前针对不同生态系统中氨氧化过程已有大量研究, 但在有机肥添加的菜地土壤中, 区分氨氧化过程中AOB和AOA对N2O和NO产生的相对贡献仍缺乏研究.
选择性抑制剂技术是区分氨氧化过程中AOB和AOA相对贡献的重要手段(Taylor et al., 2013; Zhang et al., 2019; Duan et al., 2021).有****研究发现在菌株纯培养实验中, 不同碳链长度的炔烃对AOB和AOA具有不同的抑制效果, 0.01 kPa的乙炔能完全抑制AOB和AOA的活性, 而5 μmol · L-1的辛炔可完全抑制AOB生长, 对AOA无显著抑制效果(Taylor et al., 2013).因此, 本研究采用空气、辛炔和乙炔组合选择性抑制的方法, 区分大田试验中不同比例有机、无机肥配施近3年后(2016年10月—2019年5月), AOB和AOA对碱性菜地土壤氨氧化过程中N2O和NO产生的相对贡献, 以期为有机无机肥配施实践调控菜地活性氮气体排放、实现可持续蔬菜生产提供科学依据和理论指导.
2 材料与方法(Material and methods)2.1 试验地概况试验地点位于江苏省南京市江宁区窦村(32°01′N, 118°52′E), 地处长江中下游, 属于亚热带季风气候, 年均降水量1107 mm, 年均气温15.4 ℃.试验地具有12年蔬菜种植历史, 是中国南方典型菜地, 一年种植3~5季蔬菜.试验地土壤属于淋溶土, 土壤质地为粉壤土(其中, 黏粒2.4%, 粉粒77.6%, 砂粒20.0%).土壤pH为7.90, 容重为1.3 g · cm-3, 有机碳(SOC)含量为18.8 g · kg-1, 总氮(TN含量为1.76 g · kg-1, 阳离子交换量(CEC)为25.3 cmol · kg-1, 电导率(EC)为0.17 mS · cm-1.
2.2 试验设计田间试验开始于2016年10月, 共设5个处理: 不施氮肥(CK)、单施尿素(N)、单施有机肥(M)、50%尿素+50%有机肥(M1N1)和80%尿素+20%有机肥(M1N4), 每个处理3个重复, 每个小区面积为3 m×3 m.在每季蔬菜种植前肥料作为基肥一次性施入, 其中, 4个施氮处理等氮量施肥, 均为每季作物240 kg · hm-2(以N计), 另等量施入240 kg · hm-2磷肥(P2O5)和240 kg · hm-2钾肥(K2O).不施氮处理补充相应数量磷钾肥.试验所用无机肥为尿素(含N量46.0%)、钙镁磷肥(含P2O5 14.0%)和氯化钾(含K2O 63.2%).有机肥由南京明珠有限责任公司生产, 总氮为2.0%, 有机质为30.6%, P2O5为3.2%, K2O为2.5%.田间管理采用当地农民常规措施.试验菜地一年种植3~5季蔬菜, 蔬菜种类以小青菜、苋菜、菠菜为主.土壤样品于2019年5月小青菜收获后用土钻取0~20 cm土样, 每个小区内梅花形随机采样, 一部分存储于4 ℃冰箱, 于3 d内测定理化指标, 另一部分过2 mm筛后立即进行培养试验.
参照前人研究方法(Taylor et al., 2013; Zhang et al., 2019), 选择辛炔(0.03%, 体积比)和乙炔(0.01%, 体积比)对菜地土壤进行抑制性培养, 评估氨氧化过程中AOB和AOA的相对贡献.具体操作步骤如下: 称取相当于20 g干土重的土壤样品于120 mL玻璃血清瓶, 将5种土壤样品添加灭菌去离子水, 在50% WFPS、25 ℃条件下预培养7 d, 以激活微生物活性.每种土样分别设置3个抑制剂处理: 空气(不加抑制剂)、辛炔和乙炔, 每个抑制剂处理设置3次重复.将硫酸铵(150 mg · kg-1干土)溶于灭菌去离子水, 均匀添加至土样, 调节土壤水分到60% WFPS.将辛炔(0.03%, 体积比)和乙炔(0.01%, 体积比)快速注射至橡胶塞密封的相应的血清瓶内.恒温25 ℃下培养28 d, 培养期间每隔2 d通气20 min, 并及时补充水分和相应抑制剂.
2.3 N2O和NO气体排放量测定和计算于培养1、2、3、4、5、7、10、14、21和28 d采气.采气前打开血清瓶通气20 min后, 再次用橡胶塞密封, 并添加相应抑制剂.密封6 h后用带三通阀的注射器采集30 mL气体样品测定NO, 采集8 mL气体样品测定N2O, 气体采集后重新通气并密封及添加相应抑制剂.将NO气体样品转移至含有720 mL N2的1 L密封气袋(Delin Gas Packing Co. Ltd., Dalian, China), 随后立即用氮氧化物分析仪(Thermo Environmental Instruments Inc, Franklin, MA, USA)测定.将N2O气体样品用配电子捕获检测器(ECD)的气相色谱仪(Agilent 7890A, Agilent Ltd, Shanghai, China)测定.N2O和NO产生速率为单位时间内单位质量土壤排放的气体质量(Ma et al., 2010), 计算公式见式(1).
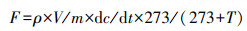 | (1) |
N2O和NO累积排放量为平均产生速率与时间的乘积, 公式见式(2).
 | (2) |
2.4 土壤样品理化性质测定在培养0、3、7、14、21和28 d进行破坏性取样, 测定土壤pH、电导率EC和无机氮含量.土壤pH和EC用精密pH计(PHS-3C)和电导率仪(FE30-K, Shanghai, China)测定, 土水比为1 ∶ 5;NH4+含量、NO2-含量和NO3-含量用2 mol · L-1 KCl浸提后分别用靛酚蓝比色法、盐酸萘乙二胺-分光光度法和双波长紫外分光光度法进行测定;土壤SOC采用外加热重铬酸钾容量法测定;全氮采用凯氏定氮法测定;CEC采用多次离心交换法测定;容重采用环刀法测定(鲁如坤, 2000).
2.5 DNA提取和定量PCR分析土壤排放N2O和NO气体于第3 d达到顶峰, 采集此时土壤样品提取DNA, 测定AOB和AOA基因丰度.利用土壤FastDNA Spin试剂盒(MP Biomedicals), 从0.5 g土样中提取微生物总DNA.利用异硫氰酸胍反复清洗DNA, 除去菜地土壤中影响分子分析的腐殖酸.使用NanoDrop ND-1000分光光度计(NanoDrop Technologies, USA)测定DNA浓度和质量.用CF96 Real-Time PCR System(Bio-Rad Laboratories Inc, USA)机器进行实时荧光定量PCR获取土壤氨氧化细菌和氨氧化古菌氨单加氧化酶基因丰度(amoA-AOB、amoA-AOA), 分别用Arch-amoAF(STA ATG GTC TGG CTT AGA CG)/Arch-amoAR(GCG GCC ATC CAT CTG TAT GT)和amoA-1f(GGG GTT TCT ACT GGT GGT)/amoA-2R(CCC CTC KGS AAA GCC TTC TTC)引物及引物序列(5′~3′)确定氨氧化微生物基因丰度(Harter et al., 2014).实时荧光定量PCR反应体系为20 μL, 包含10 μL SYBR Green(TaKaRa, Japan)、0.2 μL Rox DYEⅡ、1 μL DNA样品、8 μL灭菌水和各0.4 μL的前后引物(10 μmol · L-1).AOB的反应条件为: 94 ℃变性60 s, 60 ℃退火90 s, 72 ℃延伸90 s;AOA的反应条件为: 94 ℃变性45 s, 53 ℃退火60 s, 72 ℃延伸60 s.试验设置严格阴性对照, 并将已知拷贝数的质粒DNA进行梯度稀释, 同时进行荧光定量扩增, 得到标准曲线(3次平行), 标准曲线AOB和AOA的amoA基因扩增效率为95.0%~105.0%, R2大于0.98, 标线与样品的溶解曲线为单一峰.
2.6 统计与分析将空气和乙炔处理下N2O及NO排放量差值作为AOB和AOA对氨氧化过程的总贡献.将辛炔和乙炔处理下N2O及NO排放量差值作为AOA对氨氧化过程的贡献.AOB对氨氧化过程的贡献由总贡献与AOA贡献相减得到.采用Microsoft Excel 2016进行数据记录和计算.采用SPSS 22.0(IBM Co, Armonk, NY, USA)通过Tukey法方差分析土壤理化性质(pH、SOC、TN、EC和无机氮)和功能基因, p < 0.05具有显著性差异;对培养期间各处理N2O和NO累积排放量与土壤理化性及相关功能基因丰度进行相关性分析.利用Origin 2018(Origin Lab, USA)制作图表.
3 结果(Results)3.1 不同施肥处理菜地土壤理化性质由表 1可知, 有机无机肥配施近3年后, 施肥可显著增加菜地土壤SOC、TN、EC及无机态氮含量, 显著降低土壤pH(p < 0.05);有机肥添加处理中, 土壤NO3-含量随着尿素占比的增加而增加;M处理较N处理使土壤TN和SOC含量分别显著增加23.0%和45.1%, NO3-含量降低83.5%(p < 0.05);与N处理相比, M1N4处理显著提高土壤pH(p < 0.05), M1N1处理则与之没有显著性差异.
表 1(Table 1)
| 表 1 不同施肥处理对土壤理化性质的影响 Table 1 Effects of different fertilization treatments on soil physical and chemical properties | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 1 不同施肥处理对土壤理化性质的影响 Table 1 Effects of different fertilization treatments on soil physical and chemical properties
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
3.2 不同施肥处理菜地土壤N2O和NO排放的动态变化及累积排放量如图 1所示, 空气处理下, N2O与NO排放在培养初期迅速增加, 于第3 d达到顶峰, 随后逐渐减缓.各施肥处理间, M处理N2O和NO排放最高, 分别为317.79 ng · g-1和106.73 ng · g-1;CK处理N2O和NO排放最低, 分别为103.29 ng · g-1和45.79 ng · g-1.乙炔可显著抑制土壤N2O和NO排放(p < 0.05).有机肥添加处理中, N2O和NO累积排放量随着有机肥占比的升高而升高.与N处理相比, M和M1N1处理使土壤N2O排放量分别增加100.7%和38.8%, NO排放量分别增加77.9%和42.8%, 而M1N4处理使土壤N2O和NO排放量分别降低19.3%和4.8%.
图 1(Fig. 1)
 |
| 图 1 在抑制剂添加下(空气、辛炔和乙炔)不同施肥处理土壤N2O和NO累积排放量的动态变化 Fig. 1The dynamic changes of N2O and NO cumulative emissions from different soils under the addition of inhibitors (air, 1-octyne, and acetylene) |
3.3 不同施肥处理菜地土壤无机态氮含量变化如图 2a~2c所示, 空气处理下, 土壤NH4+含量在前7 d迅速下降, 随后趋于平缓, M处理土壤NH4+含量在第3 d下降幅度较小.辛炔较空气处理下土壤NH4+含量减少幅度降低, 于21 d降低至低水平.培养期间土壤NH4+含量在乙炔处理下无明显下降, 说明没有发生明显的氨氧化过程.
图 2(Fig. 2)
 |
| 图 2 在抑制剂添加下(空气、辛炔和乙炔)不同施肥处理土壤NH4+-N、NO2--N、NO3--N含量动态变化 Fig. 2Dynamic changes of NH4+-N、NO2--N and NO3--N in different soils with the addition of inhibitors (air, 1-octyne, and acetylene) |
由图 2d~2f可知, 土壤仅在第3 d产生NO2-累积现象, 其中, M处理土壤NO2-含量高达47.5 mg · kg-1, 是N处理的5倍.与N处理相比, M1N4处理使土壤NO2-含量降低29.2%.与空气处理相比, 添加辛炔可缓解土壤NO2-累积现象;而添加乙炔后, 土壤则没有发生NO2-累积现象.
由图 2h~2j可知, 在空气和辛炔处理下, 土壤NO3-含量呈相似的上升趋势, 并分别于第14和21 d趋于平缓.多年施肥使菜地土壤NO3-含量背景值差异显著, 导致培养期间NO3-含量曲线区分明显, 其中, M处理土壤NO3-含量与CK处理相近, 远低于其他施肥措施土壤.
3.4 不同施肥处理菜地土壤AOB和AOA基因丰度本研究中使用荧光定量PCR分析技术检测第3 d土壤AOB和AOA amoA基因丰度, 此时土壤N2O和NO排放达到高峰.由图 3a可知, 与CK处理相比, 施用氮肥可显著增加土壤AOB基因丰度(p < 0.05).各处理间, M处理AOB基因丰度最高, 为6.3×107 copies · g-1.与空气处理相比, 两种抑制剂的添加不同程度地减少AOB基因丰度.与N处理相比, M处理显著增加AOB基因丰度, M1N4处理则显著减少AOB基因丰度(p < 0.05).如图 3b所示, 与CK处理相比, N处理显著增加AOA基因丰度(p < 0.05).添加辛炔的CK、M和M1N4处理的AOA基因丰度显著高于无抑制剂添加处理.除M1N4处理外, 添加乙炔均显著抑制土壤AOA基因丰度(p < 0.05).AOA的基因丰度在各施肥处理间都高于AOB的基因丰度.
图 3(Fig. 3)
 |
| 图 3 不同施肥处理土壤氨氧化细菌(AOB-amoA)和氨氧化古菌(AOA-amoA)基因拷贝数 (柱上不同小写字母表示处理间基因丰度差异显著) Fig. 3The copy number of AOB-amoA and AOA-amoA gene in different soils |
3.5 不同施肥处理菜地土壤AOB和AOA对N2O和NO排放的相对贡献图 4显示, 施用氮肥可显著增加土壤AOB对N2O和NO排放的相对贡献(p < 0.05).施用尿素土壤AOA对N2O和NO排放的贡献都处于较低水平. 与N处理相比, M1N4处理显著降低了AOB对N2O和NO排放的相对贡献(p < 0.05).AOB对N2O和NO排放的相对贡献在各处理间都显著高于AOA(p < 0.05).
图 4(Fig. 4)
 |
| 图 4 不同施肥处理土壤氨氧化细菌(AOB)和氨氧化古菌(AOA)对N2O (a)和NO (b) 排放总量的相对贡献 Fig. 4The relative contribution of ammonia-oxidizing bacteria (AOB) and ammonia-oxidizing archaea (AOA) to the total N2O (a) and NO (b) emissions in different fertilized soils |
3.6 N2O和NO排放与AOB和AOA基因丰度及土壤理化性质的关系由图 5可以看出, 土壤N2O和NO累积排放量均与土壤AOB基因丰度呈线性关系, R2分别为0.71和0.67(p < 0.001);与土壤AOA基因丰度不具有线性关系.说明在该菜地土壤中, AOB在氨氧化过程中较AOA发挥着更重要的作用.
图 5(Fig. 5)
 |
| 图 5 N2O和NO累积排放量与土壤AOB、AOA基因丰度的线性回归分析 (阴影部分表示95%置信区间) Fig. 5Linear regression analysis of soil cumulative N2O and NO emissions with AOB and AOA gene abundances |
土壤AOB基因丰度与培养期间土壤NH4+含量和pH值呈显著负相关, 与土壤NO2-和NO3-含量呈显著正相关(p < 0.01), 土壤AOA基因丰度则与土壤无机氮含量和pH值无显著相关性(表 2).
表 2(Table 2)
| 表 2 N2O、NO累积排放量与氨氧化细菌、氨氧化古菌基因丰度及土壤理化性质间的相关关系 Table 2 Correlation coefficients among N2O cumulative emissions, NO cumulative emissions, soil physiochemical properties, ammonia-oxidizing bacteria (amoA-AOB) and ammonia-oxidizing archaea (amoA-AOA) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 2 N2O、NO累积排放量与氨氧化细菌、氨氧化古菌基因丰度及土壤理化性质间的相关关系 Table 2 Correlation coefficients among N2O cumulative emissions, NO cumulative emissions, soil physiochemical properties, ammonia-oxidizing bacteria (amoA-AOB) and ammonia-oxidizing archaea (amoA-AOA)
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
4 讨论(Discussion)本研究发现, 有机无机肥配施多年后, 菜地土壤理化性质发生显著变化, 多年输入氮肥降低了土壤pH(表 1), 大量氮素的累积导致质子H+在表层土壤富集, 进而直接或间接影响土壤pH, 该结果与中国近30年主要农田土壤pH演变结果一致(韩天富等, 2020).值得注意的是, 施入氮肥虽可显著降低土壤pH, 但该菜地仍呈碱性, 这可能是因为该菜地属于淋溶土, 具有较强的酸碱缓冲容量, 且改自水稻种植历史仅12年.有机无机肥配施能够有效减缓土壤酸化程度, 增加土壤碳氮含量(表 1).这是因为一方面有机肥中碱性物质的输入弥补了蔬菜收获后土壤碱性物质的损失;另一方面, 有机肥可增强土壤缓冲能力, 从而减缓土壤酸化(汪吉东等, 2020).
单施氮肥可显著增加土壤AOB基因丰度, 而对AOA影响不大(图 3), 该结果已得到广泛验证(Shen et al., 2008;Di et al., 2009;Zhang et al., 2019).这主要是因为NH3是AOB的主要营养物质, 施入氮肥可增加土壤NH3浓度, 从而促进AOB的生长;而AOA则主要利用土壤有机质持续矿化的低浓度NH3进行生长, 更容易受到有机氮而不是无机氮的刺激(贺纪正等, 2013).培养试验中由于所有处理新加入硫酸铵无机氮, 导致M和M1N1处理较N处理AOB基因丰度增加更多(图 3), 土壤N2O和NO排放量也相应增加(图 1).这与大田试验所得结果不一致(Zhou et al., 2019), 这与是否有作物生长有关: 在本土壤培养试验中, 60%WFPS和25 ℃的培养条件更有利于氨氧化微生物进行氨氧化过程;无机氮源的加入也刺激了有机肥处理下土壤自养微生物的生长和活性, 增加了有机肥处理中AOB的贡献(Ju et al., 2011;Zhu et al., 2013).在大田试验中, 有机肥的施用促使植物吸收利用土壤无机氮, 减少了硝化和反硝化作用底物, 提高了氮肥利用效率, 最终降低N2O排放.值得注意的是, M1N4处理土壤AOB基因丰度、N2O和NO排放显著下降(图 1和图 3), 这可能是有机无机配施比例的不同所造成的综合效应.适宜比例有机肥的施入增加了微生物间竞争关系, 使得土壤AOB基因丰度、N2O和NO排放降低(Wang et al., 2014).
相关性分析表明, 土壤N2O和NO累积排放量与土壤AOB基因丰度呈显著正相关, 但与土壤AOA基因丰度无显著性关系, 土壤AOB基因丰度则与pH呈显著负相关(表 2).表明AOB是该碱性菜地土壤氨氧化过程的主要贡献者, 且土壤pH是影响AOB基因丰度的重要因素, 不同的施肥措施可能通过改变土壤pH等性质影响AOB基因丰度, 改变土壤氨氧化过程(Lu et al., 2015).前人研究也指出在中性和碱性环境中, AOB是氨氧化过程的主要驱动者(贺纪正等, 2013).本试验中AOB基因丰度、土壤N2O和NO累积排放量与培养期间土壤NH4+含量呈显著负相关, 与土壤NO3-含量呈显著正相关.这是因为随着AOB基因丰度的上升, 氨利用效率随之上升, N2O和NO排放亦随之增加.培养期间土壤所含氨浓度越高, 表明氨氧化过程程度较低.此外, 相关性分析表明, AOA基因丰度与土壤理化性质无显著相关性关系.前人研究也发现AOB基因丰度与土壤理化性质具有相关性(Lu et al., 2020), 而AOA种群与这些环境因子间没有显著相关性, 表明AOA可能比AOB具有更强的环境适应性.本研究利用抑制剂技术发现AOA基因丰度在各土壤间都要显著高于AOB, 但对氨氧化过程的贡献却要远小于AOB(图 3和图 4), 表明微生物数量优势不一定表现为活性贡献(Taylor et al., 2013).辛炔处理下AOA基因丰度高于空气处理, 这可能是因为土壤中AOB和AOA存在竞争关系, 添加辛炔抑制了AOB生长, 从而改善了土壤AOA生长环境, 增加了AOA基因丰度.本研究培养过程中还发现NO2-峰值与N2O和NO排放高峰时间一致, 与前人研究结果相似(Duan et al., 2019), 因此, 推测硝化细菌在反硝化过程可能具有重要作用.考虑到抑制剂技术仍具有一定的局限性和有效性, 未来应采用与同位素标记等其他手段相结合的方法来探究氨氧化微生物.
5 结论(Conclusions)1) 经过3年等氮条件大田试验, 菜地土壤理化性质发生显著差异, 施肥显著增加菜地土壤SOC、TN、EC及无机态氮含量.相较于氮肥显著降低土壤pH, 较低比例有机替代(M1N4)因其输入碱性物质可减缓碱性菜地土壤酸化程度.
2) 采用选择性抑制室内培养与大田试验相结合, 结果表明, 土壤N2O和NO累积排放量与土壤AOB基因丰度呈显著正相关(p < 0.05), 与土壤AOA基因丰度无显著相关性.有机无机肥配施下AOB是碱性菜地土壤氨氧化过程的主要驱动者.
3) 等氮施加下, 与N处理相比, 较高比例有机替代(M和M1N1)使土壤N2O排放量分别增加100.7%和38.8%, NO排放量分别增加77.9%和42.8%, 而较低比例有机替代(M1N4)在一定程度上则可减缓菜地集约化生产过程中活性氮损失.不同有机无机替代比例主要通过影响AOB氨氧化过程产生差异性, 与AOA关系不紧密.
参考文献
| Ai C, Liang G Q, Sun J W, et al. 2013. Different roles of rhizosphere effect and long-term fertilization in the activity and community structure of ammonia oxidizers in a calcareous fluvo-aquic soil[J]. Soil Biology and Biochemistry, 57: 30-42. DOI:10.1016/j.soilbio.2012.08.003 |
| Anenberg S C, Schwartz J, Shindell D, et al. 2012. Global air quality and health co-benefits of mitigating near-term climate change through methane and black carbon emission controls[J]. Environmental Health Perspectives, 120(6): 831-839. DOI:10.1289/ehp.1104301 |
| Beeckman F, Motte H, Whalen J K. 2018. Nitrification in agricultural soils: impact, actors and mitigation[J]. Current Opinion in Biotechnology, 50: 166-173. DOI:10.1016/j.copbio.2018.01.014 |
| Charles A, Rochette P, Whalen J K, et al. 2017. Global nitrous oxide emission factors from agricultural soils after addition of organic amendments: A meta-analysis[J]. Agriculture, Ecosystems and Environment, 236: 88-98. DOI:10.1016/j.agee.2016.11.021 |
| Di H J, Cameron K C, Shen J P, et al. 2009. Nitrification driven by bacteria and not archaea in nitrogen-rich grassland soils[J]. Nature Geoscience, 2(9): 621-624. DOI:10.1038/ngeo613 |
| Duan P P, Fan C H, Zhang Q Q, et al. 2019. Overdose fertilization induced ammonia-oxidizing archaea producing nitrous oxide in intensive vegetable fields[J]. Science of the Total Environment, 650(2): 1787-1794. |
| Duan P P, Wu Z, Zhang Q Q, et al. 2021. Thermodynamic responses of ammonia-oxidizing archaea and bacteria explain N2O production from greenhouse vegetable soils[J]. Soil Biology and Biochemistry, 120: 37-47. |
| Duan P P, Zhou J, Feng L, et al. 2019. Pathways and controls of N2O production in greenhouse vegetable production soils[J]. Biology and Fertility of Soils, 55(3): 285-297. DOI:10.1007/s00374-019-01348-9 |
| Geng Y J, Yuan Y M, Miao Y C, et al. 2021. Decreased nitrous oxide emissions associated with functional microbial genes under bio-organic fertilizer application in vegetable fields[J]. Pedosphere, 31(2): 279-288. DOI:10.1016/S1002-0160(20)60075-3 |
| G?dde M, Conrad R. 1999. Immediate and adaptational temperature effects on nitric oxide production and nitrous oxide release from nitrification and denitrification in two soils[J]. Biology and Fertility of Soils, 30(1/2): 33-40. |
| Guo J H, Liu X J, Zhang Y, et al. 2010. Significant acidification in major Chinese croplands[J]. Science, 327(5968): 1008-1010. DOI:10.1126/science.1182570 |
| Harter J, Krause H M, Schuettler S, et al. 2014. Linking N2O emissions from biochar-amended soil to the structure and function of the N-cycling microbial community[J]. The ISME Journal, 8(3): 660-674. DOI:10.1038/ismej.2013.160 |
| 韩天富, 柳开楼, 黄晶, 等. 2020. 近30年中国主要农田土壤pH时空演变及其驱动因素[J]. 植物营养与肥料学报, 26(12): 2137-2149. |
| 贺纪正, 张丽梅. 2013. 土壤氮素转化的关键微生物过程及机制[J]. 微生物学通报, 40(1): 98-108. |
| Ju X T, Lu X, Gao Z L, et al. 2011. Processes and factors controlling N2O production in an intensively managed low carbon calcareous soil under sub-humid monsoon conditions[J]. Environmental Pollution, 159(4): 1007-1016. DOI:10.1016/j.envpol.2010.10.040 |
| Li Y Y, Chapman S J, Nicol G W, et al. 2018. Nitrification and nitrifiers in acidic soils[J]. Soil Biology and Biochemistry, 116: 290-301. DOI:10.1016/j.soilbio.2017.10.023 |
| Liu S W, Lin F, Wu S, et al. 2017. A meta-analysis of fertilizer-induced soil NO and combined NO+N2O emissions[J]. Global Change Biology, 23(6): 2520-2530. DOI:10.1111/gcb.13485 |
| 鲁如坤. 2020. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 117-332. |
| Lu X, Bottomley P J, Myrold D D. 2015. Contributions of ammonia-oxidizing archaea and bacteria to nitrification in Oregon forest soils[J]. Soil Biology and Biochemistry, 85: 54-62. DOI:10.1016/j.soilbio.2015.02.034 |
| Lu X, Taylor A E, Myrold D D, et al. 2020. Expanding perspectives of soil nitrification to include ammonia-oxidizing archaea and comammox bacteria[J]. Soil Science Society of America Journal, 84(2): 287-302. DOI:10.1002/saj2.20029 |
| Ma B L, Wu T Y, Tremblay N, et al. 2010. Nitrous oxide fluxes from corn fields: On-farm assessment of the amount and timing of nitrogen fertilizer[J]. Global Change Biology, 16(1): 156-170. |
| Mosier A, Kroeze C, Nevison C, et al. 1998. Closing the global N2O budget: nitrous oxide emissions through the agricultural nitrogen cycle[J]. Nutrient Cycling in Agroecosystems, 52(2/3): 225-248. DOI:10.1023/A:1009740530221 |
| 宁启文, 胡乐鸣. 2018. 中国农业年鉴[M]. 北京: 中国农业出版社, 11-12. |
| Ren F L, Zhang X B, Liu J, et al. 2018. A synthetic analysis of livestock manure substitution effects on organic carbon changes in China's arable topsoil[J]. Catena, 171(9): 1-10. |
| Shen J P, Zhang L M, Zhu Y G, et al. 2008. Abundance and composition of ammonia-oxidizing bacteria and ammonia-oxidizing archaea communities of an alkaline sandy loam[J]. Environmental microbiology, 10(6): 1601-1611. |
| Taylor A E, Vajrala N, Giguere A T, et al. 2013. Use of aliphatic n-alkynes to discriminate soil nitrification activities of ammonia-oxidizing thaumarchaea and bacteria[J]. Applied and Environmental Microbiology, 79(21): 6544-6551. DOI:10.1128/AEM.01928-13 |
| 汪吉东, 戚冰洁, 张永春, 等. 2012. 长期施肥对砂壤质石灰性潮土土壤酸碱缓冲体系的影响[J]. 应用生态学报, 23(4): 1031-1036. |
| Wang Y, Zhu G B, Song L Y, et al. 2014. Manure fertilization alters the population of ammonia-oxidizing bacteria rather than ammonia-oxidizing archaea in a paddy soil[J]. Journal of Basic Microbiology, 54(3): 190-197. DOI:10.1002/jobm.201200671 |
| 解开治, 徐培智, 蒋瑞萍, 等. 2016. 有机无机肥配施提升冷浸田土壤氮转化相关微生物丰度和水稻产量[J]. 植物营养与肥料学报, 22(5): 1267-1277. |
| Zhang W, Li Y Z, Li Q Z, et al. 2016. Isotope signatures of N2O emitted from vegetable soil: Ammonia oxidation drives N2O production in NH4+-fertilized soil of North China[J]. Scientific Reports, 6(1): 29257. DOI:10.1038/srep29257 |
| Zhang X, Duan P P, Wu Z, et al. 2019. Aged biochar stimulated ammonia-oxidizing archaea and bacteria-derived N2O and NO production in an acidic vegetable soil[J]. Science of the Total Environment, 687: 433-440. |
| Zhou J, Li B, Xia L L, et al. 2019. Organic-substitute strategies reduced carbon and reactive nitrogen footprints and gained net ecosystem economic benefit for intensive vegetable production[J]. Journal of Cleaner Production, 225(10): 984-994. |
| Zhu X, Burger M, Doane T A, et al. 2013. Ammonia oxidation pathways and nitrifier denitrification are significant sources of N2O and NO under low oxygen availability[J]. Proceedings of the National Academy of Sciences of the United States of America, 110(16): 6328-6333. |
| Zhuang M H, Lam S K, Zhang J, et al. 2019. Effect of full substituting compound fertilizer with different organic manure on reactive nitrogen losses and crop productivity in intensive vegetable production system of China[J]. Journal of Environmental Management, 243: 381-384. |
