 , 崔建升1,2, 剧泽佳1, 申立娜1, 张璐璐1,2
, 崔建升1,2, 剧泽佳1, 申立娜1, 张璐璐1,2 , 付雨1
, 付雨11. 河北科技大学环境科学与工程学院, 石家庄 050018;
2. 河北省污染防治生物技术实验室, 石家庄 050018
收稿日期: 2020-06-13; 修回日期: 2020-09-09; 录用日期: 2020-09-09
基金项目: 国家自然科学基金(No.41601547);河北省自然科学基金(No.D2019208152);河北科技大学研究生创新资助项目
作者简介: 秦珊(1995-), 女, E-mail: 18330820355@163.com
通讯作者(责任作者): 张璐璐, E-mail: zhanglulu19850703@163.com
摘要:底栖动物对维持和稳定湖泊生态系统结构与功能具有重要作用.为了探明长期强人为干扰条件下底栖动物群落结构特征及其主要环境影响因子,本研究分别于2009年和2018年的4月和8月采集和分析了白洋淀8个区水体、沉积物和底栖动物样品.根据人为干扰程度的不同,将白洋淀分为重度干扰区(HD)、中度干扰区(MD)和轻度干扰区(LD),同时分析了3类生境的理化参数和底栖动物群落结构与多样性指数.研究结果表明:1就水体和沉积物理化参数而言,总磷(TP)、总氮(TN)、氨氮(NH4+)、硝酸盐(NO3-)、磷酸盐(PO43-)和沉积物总氮(TNs)、总磷(TPs)等理化参数在重度干扰区呈最高值;2就底栖动物群落组成而言,在重度干扰区底栖动物群落物种丰度、生物量、密度均最低,且优势种大多以水生昆虫为主;3就底栖动物群落多样性指数而言,2009年,白洋淀底栖动物群落Margalef丰富度指数D值(0.84)和Shannon-Wiener多样性指数H'值(1.13)均在中度干扰区最高,而Pielou均匀度指数J值(0.53)在轻度干扰区最高,这些指数均与沉积物总氮(TNs)呈显著负相关;2018年,D值(2.02)和H'值(2.21)在轻度干扰区中最高,而J值(0.84)在重度干扰区中最高,D值和H'值与水深(Water depth,WD)呈正相关关系,而与硝酸盐(NO3-)和总磷(TP)等呈负相关关系;4RDA分析结果表明,白洋淀底栖动物群落组成的主要环境影响因子在2009年为WD和pH,而在2018年为沉积物中总磷(TPs).2009-2018年,白洋淀底栖动物群落(主要影响因子从WD和pH变为沉积物总磷)和多样性指数(主要影响因子从TNs变为WD、TN、TP、NO3-、TNs等)的主要环境影响因子发生了显著变化.因此,针对主要环境影响因子的时间变化,在不同时期底栖动物群落的恢复需采取不同措施,本研究结果可为白洋淀生态修复提供理论和数据基础.
关键词:人为干扰底栖动物群落结构多样性指数白洋淀
Changes of benthic invertebrate community in the Baiyangdian Lake and analysis of main environmental factors under the condition of human disturbance
QIN Shan1
, CUI Jiansheng1,2, JU Zejia1, SHEN Lina1, ZHANG Lulu1,2, FU Yu11. Academy of Environmental Science, Hebei University of Science and Technology, Shijiazhuang 050018;
2. Biotechnology Laboratory for Pollution Control in Hebei Province, Shijiazhuang 050018
Received 13 June 2020; received in revised from 9 September 2020; accepted 9 September 2020
Abstract: Benthic invertebrate communities play an important role in maintaining and stabilizing the structure and function of lake ecosystems. In order to reveal the structural characteristics of benthic invertebrate communities and the main environmental influencing factors under the condition of long-term strong human disturbance, water, sediments, and biological samples were simultaneously collected and analyzed in Baiyangdian Lake during the period of 2009 and 2018. According to the different degree of human disturbance, Baiyangdian Lake can be divided into heavy disturbance area (HD), moderate disturbance area (MD), and mild disturbance area (LD). In this study, the physical-chemical parameters and the structure and diversity index of benthic invertebrate communities were analyzed among three habitats. The results showed that: 1In terms of physical-chemical parameters of water and sediment, such as total phosphorus (TP), total nitrogen (TN), ammonia nitrogen (NH4+), nitrate (NO3-), phosphate (PO43-) and sediment total nitrogen (TNs) and total phosphorus (TPs) showed the highest values in HD; 2In terms of the composition of benthic invertebrate communities, the abundance, biomass, and density of benthic invertebrate communities exhibited the lowest values in HD, and the dominant species were aquatic insects; 3 In terms of the diversity indexes of benthic invertebrate communities, in 2009, the Margalef richness index D value (0.84) and Shannon-Wiener diversity index H' value (1.13) of the benthic community showed the highest values in MD; while the Pielou evenness index J value (0.53) showed the highest value in LD, and all of the indexes were significantly negatively correlated with sediment total nitrogen (TNs); while in 2018, the D (2.02) and H' (2.21) showed the highest values in LD, while the J (0.84) exhibited the highest value in HD; D and H' values were positively correlated with WD, while negatively correlated with nitrate (NO3-) and total phosphorus (TP); 4The results of RDA analysis showed that WD/pH and sediment total phosphorus (TPs) were the main environmental affecting factors for the benthic invertebrate communities in 2009 and 2018, respectively. From 2009 to 2018, the main environmental influencing factors of benthic invertebrate communities (the main impact factors changed from WD and pH to sediment total phosphorus) and diversity indexes (the main impact factors changed from TNs to WD, TN, TP, NO3-, TNs and other factors) were different. Therefore, according to the temporal variation of the main environmental influencing factors, different restoration measures need to be taken in different periods. The study will provide the theoretical and data basis for the restoration of Baiyangdian Lake.
Keywords: human disturbancebenthic invertebrate communitycommunity structurediversity indexBaiyangdian Lake
1 引言(Introduction)湖泊作为维持人类生存和发展的重要水生态系统(杨柳燕等, 2019), 在水源涵养、气候调节、生物多样性保护和经济发展中发挥着重要作用(Penning, 2012; 刘丹等, 2019; 相晨等, 2019; 宋晓薇等, 2019).同样, 湖泊因具有明确边界且通常位于人口密集的平原, 也极易受到严重的人为干扰(Yang et al., 2017).近年来, 随着工业化和城市化的发展, 人类活动影响加剧(卞少伟等, 2016), 导致湖泊环境问题日益凸显, 如富营养化(Wu et al., 2010)、生物多样性减少(梁红等, 2018), 以及服务功能退化(孟云飞等, 2019)等.其中, 底栖动物群落作为湖泊生态系统中的重要组成部分, 在维持生态系统的物质循环和能量流动中发挥着重要的作用(Vanni et al., 2002; 张宇航等, 2018; 刘淑丽等, 2019;王琰等, 2019).然而, 值得关注的是, 底栖动物群落对环境变化非常敏感, 并且人类生产生活产生的污染物会影响底栖动物群落结构的组成.环境变化会引起底栖动物群落的丰富度、生物量及物种组成发生改变(Marcello et al., 2017), 这些影响因素包括物理因素(如底质(Bazzanti et al., 2012)、温度(许亚红等, 2019)、水流(Pedro et al., 2008)、水深(杨青瑞等, 2010))和化学因素(如溶解氧(其木乐等, 2020)、氮(Yu et al., 2018)、磷(蔡佳亮等, 2011)).人为干扰会导致底栖动物生境条件改变, 进而影响底栖动物的群落结构和多样性.因此, 以底栖动物群落作为生物指示群落, 易于明晰人为干扰对湖泊生态系统的影响.
白洋淀作为华北地区最大的浅水草型湖泊, 在维持区域生物多样性和生态安全中发挥着重要的作用, 尤其在雄安新区成立后, 白洋淀将为构建蓝绿交织的新区提供重要生态支撑.然而, 近年来由于自然因素和人为活动的共同影响, 导致白洋淀面临着复合污染严重(汪敬忠等, 2020)、水域面积减少(Xu et al., 1998; 姚敏奇, 2019)、水生态系统不断退化和生物多样性下降(张素珍等, 2007)等生态环境问题.自2017年雄安新区设立后, 人们对白洋淀实施了多项生态恢复措施, 包括生态补水和入湖污染物削减等, 对白洋淀生态系统恢复产生了显著效果.目前, 多数研究关注白洋淀水环境改善效果(刘鑫等, 2020)及其对浮游生物的影响(李娜等, 2020), 然而对白洋淀底栖动物群落结构和多样性则较少关注.鉴于此, 本研究以白洋淀为研究对象, 分析2009年和2018年白洋淀底栖动物群落组成和多样性指数及主要水体和沉积物理化参数, 识别不同时期白洋淀底栖动物群落及其主要环境影响因子, 以期为下一步生态环境整治工程的实施提供理论基础.
2 材料与方法(Materials and methods)2.1 研究区概况白洋淀由藻苲淀、马棚淀、腰葫芦淀等143个大小不等的淀泊组成, 总面积为366 km2, 是华北地区最大的内陆浅水湖泊(朱金峰等, 2019; 闫欣等, 2019; 雷瑾等, 2019).自20世纪60年代以来, 白洋淀上游河流来水持续减少, 连续5年的干旱(1983—1988年)对湖泊生态系统产生了重大影响(Yi et al., 2018).水位和养分含量是降低湖泊生态稳定性的重要压力源, 降雨减少和自然径流减少也加速了水生生态退化(Kong et al., 2017).强人为干扰作用下使白洋淀形成了包括居民区、养殖区、旅游区在内的各种生境, 导致白洋淀水环境污染形势严峻, 从而严重影响白洋淀生态系统健康, 并造成白洋淀生物多样性下降.为了研究长期强人为干扰对白洋淀底栖动物群落组成和多样性指数的影响, 识别其主要环境影响因子, 本研究于2009年和2018年的4月和8月分别在白洋淀8个区共32个采样点(图 1), 采集底栖动物样品、水体和沉积物样品.其中, Ⅰ和Ⅱ区因其主要受到城市污水的污染, 称为重度干扰区(HD);Ⅴ、Ⅶ和Ⅷ区主要受养殖业和农村生活污水的污染, 称为中度干扰区(MD);Ⅲ、Ⅳ和Ⅵ区较少受人为干扰, 称为轻度干扰区(LD).
图 1(Fig. 1)
 |
| 图 1 白洋淀位置及监控点分布 (Ⅰ. 藻苲淀区, Ⅱ. 南刘庄区, Ⅲ. 烧车淀区, Ⅳ. 枣林庄区, Ⅴ. 淀头区, Ⅵ. 圈头区, Ⅶ. 采蒲台区, Ⅷ. 端村区) Fig. 1Sampling sites of Baiyangdian Lake |
2.2 底栖动物样品的采集与处理利用改良的1/16 m2彼德森采泥器进行定量采样, 每个采样点重复采集3~4次(陈伟明等, 2005);采集的样品用孔径为40目网筛进行筛选洗涤, 将筛选出的底栖动物用福尔马林溶液(4%)固定(Yu et al., 2018).底栖动物的种类鉴定根据分类学特征尽可能鉴定到种级水平(刘月英等, 1979; Brinkhurst et al., 1986; Morse et al., 1994), 进行计数和称重, 根据采样面积换算成密度(ind · m-2)与生物量(g · m-2)(张璐璐, 2013).
2.3 水体和沉积物样品采集与主要理化因子分析采用容量为4 L的低密度聚乙烯容器收集经BC-2300便携式自动水质采样器采集的水样, 在低温下保存并送回实验室进行分析.水深(Water depth, WD)、pH、温度(Temperature, T)和溶解氧(Dissolved oxygen, DO)等指标直接在现场用便携式水质分析仪(HYDROLAB, DS5X)进行测定(Tesgaye et al., 2006; 张欢等, 2015).参照相关国家标准方法(《水和废水分析检测方法(第4版)》)(国家环境保护总局《水和废水监测分析方法》编委会, 2006), 在实验室内进行硝酸(NO3-)、氨氮(NH4+)、总氮(TN)、总磷(TP)、磷酸盐(PO43-)等指标测定(Ma et al., 2011).
将沉积物样品用聚乙烯塑封袋封口标记, 冷藏并送至实验室分析.干燥后的样品转移至玻璃盘中, 筛选去除异物, 用聚乙烯袋保存备用(张璐璐, 2013).沉积物总氮(TNs)和总磷(TPs)按照标准方法(GB7173—1987和GB9387—1988)测定(陈伟明等, 2005);沉积物总有机碳(Total Organic Carbon, TOCs)根据GB9834—1988标准方法进行测定(鲁如坤等, 2000).
2.4 底栖动物统计分析方法底栖动物群落特征分析采用Shannon-Wiener多样性指数(H′)(Shannon-Wiener, 1949)、Margalef丰富度指数(D)(Margalef, 1958)、Pielou均匀度指数(J)(史键等, 2013)、物种优势度指数(Y)(Lampitt et al., 1993), 具体计算公式如下所示:
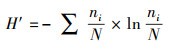 | (1) |
 | (2) |
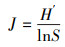 | (3) |
 | (4) |
2.5 数据分析利用CANOCO4.5软件进行多元相关分析, 主要采用非约束线性模型的主成分分析(Principal component analysis, PCA)和约束线性模型的冗余分析(Redundancy analysis, RDA).在进行生物数据(生物密度和生物参数)与环境因子多元相关分析前, 首先对生物数据进行非约束排序, 排序结果显示4个排序轴均小于3, 选用线性模型更加合理, 因此, 在约束排序中选用冗余分析(Terbraak, 1986), 并对生物密度数据进行对数转换.生物密度数据在排序前进行中心化, 进行冗余分析时对生物密度数据进行标准化处理.冗余分析中环境变量的筛选采用向前引入法(a=0.05), 基于蒙特卡洛置换检验约束模型的显著性(n=9999).水体和沉积物理化参数数据及多样性指数均在SPSS24中进行单因素方差分析.利用Origin8.6及ArcGIS10.5进行绘图.
3 结果与分析(Result and analysis)3.1 水样和沉积物样品理化特征分析就空间分布而言, 水体理化参数中除DO、pH、WD和T外, 其它理化参数均在重度干扰区呈最高值, 沉积物中TNs和TPs在重度干扰区最高, 其次为中度干扰区;就时间变化而言, 除DO、pH外, 2009年其它水质和沉积物理化参数的值高于2018年, 且TN、NH4+、NO3-和沉积物TPs呈显著差异(p < 0.05)(图 2).
图 2(Fig. 2)
 |
| 图 2 2009年和2018年水体和沉积物理化参数空间分布 (a.沉积物理化参数, b.水体理化参数, c.水深, d.温度和pH) Fig. 2Spatial distribution of physical and chemical parameters of water bodies and sediments in 2009 and 2018 |
3.2 底栖动物的群落组成时空变化就群落组成时间变化而言, 2009年共采集大型底栖动物21(属)种, 隶属于3门4纲10科;2018年共采集24(属)种大型底栖动物, 隶属于3门6纲15科.与2009年相比, 2018年底栖动物群落种类呈增加趋势.就群落组成空间分布而言, 2009年和2018年底栖动物种类在重度干扰区最少, 分别为2种(属)和14种(属);在轻度干扰区分别为15种(属)和20种(属).就优势种空间分布而言, 2018年优势种在轻度干扰区多为软体动物和甲壳类动物, 在重度和中度干扰区优势种则以水生昆虫为主.此外, 2009年白洋淀底栖动物优势种均为耐污种, 而2018年白洋淀底栖动物优势种包含清洁种和耐污种.就优势度时间变化而言, 2018年底栖动物优势种优势度比2009年增加.就物种平均丰度空间变化而言, 2009年和2018年重度干扰区丰度最低;就物种平均丰度时间变化而言, 与2009年相比, 2018年白洋淀底栖动物群落物种丰度呈增加趋势(图 3).
图 3(Fig. 3)
 |
| 图 3 2009年和2018年白洋淀底栖动物群落结构组成(a)和物种丰度(b) Fig. 3Community structure composition (a) and species abundance (b) of benthic Fauna in Baiyangdian Lake in 2009 and 2018 |
3.3 底栖动物密度和生物量的时空分布特征就底栖动物密度而言, 2009年共采集底栖动物2260个, 其密度为0.00~776.00 ind · m-2, 平均密度为282.50 ind · m-2;且在中度干扰区平均密度最高为438.67 ind · m-2, 重度干扰区平均密度最低为28.00 ind · m-2(图 4a).2018年共采集白洋淀底栖动物6584个, 其密度为354.00 ~1338.00 ind · m-2, 平均密度为823.00 ind · m-2;在轻度干扰区平均密度最高为1080.67 ind · m-2, 重度干扰区的平均密度最低为515.00 ind · m-2(图 4b).与2009年相比, 2018年白洋淀底栖动物群落平均密度呈增加趋势.
图 4(Fig. 4)
 |
| 图 4 2009年和2018年白洋淀底栖动物群落各采样点密度(a、b)和生物量分布(c、d) Fig. 4Density and biomass distributions of benthic communities in Baiyangdian in 2009 and 2018 |
就底栖动物生物量而言, 2009年白洋淀底栖动物群落生物量为315.93 g · m-2, 平均生物量为39.49 g · m-2;中度干扰区平均生物量最高为93.94 g · m-2, 重度干扰区平均最低为0.40 g · m-2(图 4c).2018年白洋淀底栖动物群落生物量为9114.34 g · m-2, 平均生物量为1139.29 g · m-2, 平均生物量最高值和最低值分别出现在中度干扰区(1954.80 g · m-2)和重度干扰区(734.23 g · m-2)(图 4d).相比于2009年, 2018年白洋淀底栖动物密度和生物量总体上呈增加趋势.
3.4 底栖动物群落结构时间分布特征2009年和2018年白洋淀底栖动物群落的H′值、D值和J值空间分异性较大(图 5).2009年, 白洋淀底栖动物群落D值、H′值和J值的均值分别为0.63、0.92和0.51;其中, D值(0.84)和H′值(1.13)均在中度干扰区最高, 而J值(0.53)在轻度干扰区最高.2018年, 白洋淀底栖动物群落D值、H′值和J值的均值分别为1.73、2.01和0.81;其中, D值(2.02)和H′值(2.21)在轻度干扰区最高, 而J值(0.84)在重度干扰区最高.与2009年相比, 2018年白洋淀底栖动物群落D、H′和J值的平均值均呈增加趋势.单因素方差分析结果表明, 仅D值(p < 0.05)呈显著差异.
图 5(Fig. 5)
 |
| 图 5 2009年和2018年白洋淀底栖动物的Shannon-Wiener多样性指数(H′)、Margalef丰富度指数(D)、Pielou均匀度指数(J)空间分布 Fig. 5Spatial distribution of Shannon-Wiener diversity index (H′), Margalef richness index (D), Pielou evenness index (J) of benthic invertebrate community in Baiyangdian Lake in 2009 and 2018 |
将各类多样性指数平均值与环境变量进行相关性分析, 结果表明, 多样性指数平均值与一个或多个环境因子存在显著相关关系(表 1).2009年白洋淀底栖动物群落的H′值(r=-0.745, p < 0.05)、D值(r=-0.714, p < 0.05)和J值(r=-0.772, p < 0.05)与沉积物TNs呈显著负相关;2018年白洋淀底栖动物群落的D值与WD(r=0.879, p < 0.01)和DO(r=0.743, p < 0.05)呈正相关, 而D值与TP(r=-0.913, p < 0.01)、TN(r=-0.794, p < 0.05)、NO3-(r=-0.848, p < 0.01)和TNs(r=-0.873, p < 0.01)呈负相关.H′值与WD(r=0.731, p < 0.05)呈正相关, 与TP(r=-0.752, p < 0.05)和NO3-(r=-0.722, p < 0.05)呈负相关.J值与环境变量无相关关系.
表 1(Table 1)
| 表 1 底栖动物群落多样性指数与环境变量的Pearson相关性 Table 1 Pearson correlation between benthic community diversity index | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 1 底栖动物群落多样性指数与环境变量的Pearson相关性 Table 1 Pearson correlation between benthic community diversity index
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
3.5 底栖动物群落与环境因子的关系运用RDA分析, 分别对2009年和2018年白洋淀底栖动物群落与环境因子的相关性进行研究.结果表明, 2009年白洋淀底栖动物群落主要与4种环境因子(WD、pH、TOCs和PO43-)呈显著相关关系, 轴1和轴2共解释了84.9%的群落结构变化, 其中, pH(p < 0.05)和WD(F=5.3, p < 0.05)对群落变异的解释度分别为54.7%和23.4%.轴1和轴2分别与pH和WD呈正相关.此外, WD与甲壳类和软体动物呈正相关, pH与环节动物和昆虫类动物呈显著正相关(图 6a).
图 6(Fig. 6)
 |
| 图 6 2009年(a)和2018年(b) 底栖动物群落物种组成与环境因子的RDA排序图 Fig. 6RDA ranking of benthic animal species composition and environmental factors in 2009 (a) and 2018 (b) |
2018年白洋淀底栖动物群落与4种关键环境因子(TPs、TOCs、WD、pH)呈显著相关关系, 共解释了61.98%的群落结构变化, 其中, TPs对群落变异的解释度为43%(图 6b).其中, 轴1与TPs呈负相关, 而与TOCs呈正相关.此外, TPs和TOCs分别与昆虫类/环节动物和甲壳类/软体动物呈正相关, 而WD与昆虫类和环节动物呈负相关.
4 讨论(Discussion)4.1 人为干扰对白洋淀水环境的影响自20世纪60年代以来, 由于人为活动(包括水库的开发、地下水开采、农业灌溉和土地利用等)和多种气候因素的影响, 导致白洋淀水量下降(Jing et al., 2019), 湖泊面积萎缩;此外, 上游工业废水、生活废水和畜禽养殖废水通过各种途径源源不断汇入淀区, 进一步加剧了白洋淀湖泊污染(Zhang et al., 2020; 王东升等, 2020), 这也导致白洋淀水生生物种类减少(Yang et al., 2014).此前的研究结果表明:1988—2017年白洋淀水质总体呈恶化趋势, 水质下降主要是由于入淀水量减少和废水的直接排放(王欢欢等, 2020).与2009年相比, 2018年多数水质和沉积物理化参数的值较低, 如:总氮和总磷2009年的平均值分别为0.28 mg · L-1和4.84 mg · L-1, 2018年分别为0.15 mg · L-1和2.68 mg · L-1, 表明白洋淀水质和沉积物质量出现好转趋势(王欢欢等, 2020).这与雄安新区成立后白洋淀流域实施多项流域综合治理措施有关(刘鑫等, 2020).例如, 引黄入冀补淀等生态调水工程, 持续的调水可以有效稀释污染物, 促进污染物的平流和扩散(Quan et al., 2020).
4.2 白洋淀底栖动物群落结构和多样性尽管已对白洋淀进行了多次调水(Tang et al., 2018), 但生态系统依然较为脆弱, 且生态功能退化(Yu et al., 2017), 如水生生物(水生高等植物(刘世存等, 2020)、鱼类和浮游生物(崔秀丽等, 1999; 李云虎等, 2013; 付显婷等, 2020)、底栖动物(Yan et al., 2020))多样性降低、景观破碎化等.
其中, 大型底栖动物因其对外界环境质量变化较为敏感, 且活动能力较差, 进而其群落组成及多样性更容易受自然和人为干扰的影响(芦康乐等, 2020).具体表现为白洋淀底栖动物种群数量减少, 生物多样性降低, 耐污种在生物群落中比例增加(Yan et al., 2020).如:底栖动物种类从1958年的53种减少到1975年的35种, 1994年进一步下降到21种(Xu et al., 1998).本研究结果表明, 白洋淀底栖动物的种类和平均密度在2018年分别为24种(属)和823 ind · m-2;与2009年相比, 其种类和生物量均呈现增加趋势.此外, 在2018年白洋淀底栖动物群落的优势种从耐污种(谢松等, 2010; 杨雨风等, 2019)向清洁种(如:中华米虾(Caridina denticulata)和日本沼虾(Macrodrachium nipponensis)等清洁种)和耐污种共存转变.就多样性指数而言, 与2009年相比, 白洋淀底栖动物群落丰富度指数值(1.73)、多样性指数值(2.01)和均匀度指数值(0.81)均有所提高(谢松等, 2010; 杨雨风等, 2019).因此, 近年来白洋淀底栖动物种类、种群丰度和均匀度呈现增加的趋势.
这可能与白洋淀实施生态补水措施有关(刘鑫等, 2020).生态补水使白洋淀水位升高, 水面面积得到一定程度恢复(杨薇等, 2020), 引起底栖物种增加.根据岛屿生态学理论, 物种数量与生境面积呈正相关, 物种数量随水面面积增加而增加, 并趋于稳定(田世民等, 2010).遥感调查结果亦可证实, 1995年白洋淀水域面积为177 km2, 而2005年仅为53 km2, 2015年“引黄济淀”后为78.5 km2(王东升等, 2020).另外, 在海珠湿地底栖动物群落的研究中, 发现水面面积较大的水体可以承载更多的物种(黄业辉等, 2020);小南湖在进行生态补水后, 湖区的水生生态环境得到改善, 底栖动物群落物种多样性也随之增加(虞左明等, 1997);这些研究与本研究结果相似.本研究也表明D和H′值与水深(WD)呈正相关, 随着水位的升高, 底栖动物丰富度和多样性均会升高.湖泊水域面积越大, 环境异质性越强, 其抗干扰能力越强, 物种多样性越高(Wagner et al., 2014).
就空间分异性而言, 2009年和2018年底栖动物群落的密度、生物量及H′值、D值均在重度干扰区呈现最低值(图 4).这与其它研究的结果类似, 如对滦河(沈月含等, 2020)和北京市(何玉晓等, 2020)河流的研究结果表明, 在人为干扰较强的区域, 底栖动物群落的多样性指数和丰富度指数值较低.由于人为干扰会影响河流的形态、流速、水质, 也会降低生境的复杂程度(Rana et al., 2017), 进而影响底栖动物的生存环境, 导致生物多样性降低(Tang et al., 2006; Strayer et al., 2010; 沈洪艳等, 2015; Solis et al., 2016; 吴红斌等, 2017).大量研究表明, 人为干扰成为影响底栖动物群落结构的主要原因(Cai et al., 2012; 李艳利等, 2015; Cai et al., 2017).
4.3 环境因子对底栖动物群落结构的影响湖泊环境理化因子与底栖动物群落的种类组成、生物量和分布有着显著相关性(王银东等, 2005).本研究结果表明, 底栖动物群落结构和多样性的主要环境影响因子呈显著时空差异.在2009年pH和WD为影响底栖动物群落结构的主要环境因子, 该研究结果与在高岚河(许亚红等, 2019)、淮河流域(刘祥等, 2016)、长荡湖(尹子龙, 2020)的研究结果类似, 在水位较深的区域底栖动物群落中的软体动物和甲壳类动物的密度较高.pH值对底栖动物群落的繁殖能力影响较大, 会导致底栖无脊椎动物多样性降低(李永刚等, 2018).而2018年底栖动物群落结构的主要环境影响因子为沉积物中的TPs, 这与杭州西溪湿地(路强等, 2013)的研究结果类似, 底泥中的总磷含量与底栖动物群落密度呈正相关.
就空间差异性而言, 研究区不同, 大型底栖动物群落的主要影响因素也不同.影响大型底栖动物群落的环境因素非常复杂, 不同研究区对同一底栖动物的主要影响因素也有所不同(朱利明等, 2019).此前研究表明水体的T(王丑明等, 2016)、DO(孟云飞等, 2019)和TP、TN等理化因子(孙枭琼等, 2019)影响着底栖动物群落的空间分布;而本研究中这些环境因子对底栖动物群落结构的影响不显著.此外, 针对杭州西溪湿地的研究表明(路强等, 2013), 随着水体深度的增加, 大型底栖动物的密度和种类数通常减少;而本研究中水深与软体动物和甲壳类动物密度呈正相关, 而昆虫类和环节动物的密度则随着水深的增加而减小, 这可能与研究区不同有关.
就时间变化而言, 底栖动物群落结构和多样性的主要环境影响因子呈现显著的时间变化趋势.本研究结果表明, 在2009年WD是白洋淀底栖动物群落结构和多样性的主要环境影响因子;而2018年白洋淀底栖动物群落分布与WD不相关, 这主要是因为经过长期的生态调水, 白洋淀水位提高, 水域面积增加.根据限制因子定律, WD已经不成为底栖动物分布的限制因子.随着雄安新区的成立, 白洋淀的生态补水趋于稳定, 形成了“引黄济淀”、“两库连通”、“南水北调”等多源补水态势(刘世存等, 2020).
5 结论(Conclusions)1) 就白洋淀底栖动物群落种类而言, 物种数量和密度均呈增加趋势.2009年采集了底栖动物大型底栖动物群落21(属)种, 而2018年采集了24(属)种;平均生物量由2009年的39.49 g · m-2上升到2018年的1139.29 g · m-2;就空间分异规律而言, 底栖动物群落的密度、生物量均在重干扰区呈最低值.
2) 白洋淀底栖动物群落多样性指数呈增加趋势.与2009年相比, 2018年白洋淀底栖动物群落Margalef丰富度指数D值(1.73)、Shannon-Wiener多样性指数H′值(2.01)和Pielou均匀度指数J值(0.81)增加的比例分别为174.60%、118.47%和58.82%.
3) 2009年和2018年白洋淀底栖动物群落组成结构主要影响因子呈现时间差异性, 在2019年主要环境影响因子为pH/WD, 而在2018年为沉积物中的TPs.
参考文献
| Bazzanti M, Mastrantuono L, Solimini A G. 2012. Selecting macroinvertebrate taxa and metrics to assess eutrophication in different depth zones of Mediterranean lakes[J]. Fundamental and Applied Limnology, 180(2): 133-143. DOI:10.1127/1863-9135/2012/0200 |
| 卞少伟, 姜伟, 梅鹏蔚, 等. 2016. 基于底栖动物指数法的湖泊生态系统健康评价研究进展[J]. 环境保护与循环经济, 36(10): 49-52. |
| Brinkhurst R O. 1986. Guide to the freshwater aquatic Microdrile Oligochaetes of North America[J]. Journal of the North American Benthological Society, 6(1): 78-79. |
| Cai Y, Gong Z, Qin B. 2012. Benthic macroinvertebrate community structure in Lake Taihu, China: Effects of trophic status, wind-induced disturbance and habitat complexity[J]. Journal of Great Lakes Research, 38(1): 39-48. DOI:10.1016/j.jglr.2011.12.009 |
| Cai Y, Xu H, Vilmi A, et al. 2017. Relative roles of spatial processes, natural factors and anthropogenic stressors in structuring a lake macroinvertebrate metacommunity[J]. Science of the Total Environmental, 601-602: 1702-1711. DOI:10.1016/j.scitotenv.2017.05.264 |
| 蔡佳亮, 苏玉, 文航, 等. 2011. 滇池流域入湖河流丰水期大型底栖动物群落特征及其与水环境因子的关系[J]. 环境科学, 32(4): 982-989. |
| 陈伟明, 黄翔飞, 周万平, 等. 2005. 湖泊生态系统观测方法[M]. 北京: 中国环境科学出版社, 17-37. |
| 崔秀丽, 候玉聊. 1999. 白洋淀生态演变的原因、趋势与保护对策[J]. 保定学院学报, (2): 86-89. |
| 付显婷, 杨薇, 赵彦伟, 等. 2020. 白洋淀浮游动物群落结构与水环境因子的关系[J]. 农业环境科学学报, 39(6): 1271-1282. |
| 国家环境保护总局《水和废水监测分析方法》编委会. 2006. 水和废水监测分析方法[M]. 第4版. 北京: 中国环境科学出版社. |
| Han Q, Tong R, Sun W C, et al. 2020. Anthropogenic influences on the water quality of the Baiyangdian Lake in North China over the last decade[J]. Science of the Total Environment, 701: 134929. DOI:10.1016/j.scitotenv.2019.134929 |
| 贺玉晓, 李珂, 任玉芬. 2020. 春季北京市河流大型底栖动物群落结构特征及影响因子分析[J]. 环境科学, 41(6): 2951-2962. |
| 黄业辉, 范存祥, 吴中奎, 等. 2020. 广州市海珠湿地大型底栖动物群落物种组成和分布初探[J]. 湿地科学, 18(2): 200-206. |
| 江为群, 刘淑丽, 简敏菲, 等. 2020. 鄱阳湖主要入湖段典型底栖动物体内微塑料累积特征[J]. 生态学杂志, 39(4): 1273-1280. |
| Jing Y, Maryna S, Carolien K, et al. 2019. Nutrient losses to surface waters in Hai He basin: A case study of Guanting reservoir and Baiyangdian lake[J]. Agricultural Water Management, 213: 62-75. DOI:10.1016/j.agwat.2018.09.022 |
| Lampitt R S, Wishner K F, Turley C M, et al. 1993. Marine snow studies in the Northeast Atlantic Ocean: Distribution, composition and role as a food source for migrating plankton[J]. Marine Biology, 116(4): 689-702. DOI:10.1007/BF00355486 |
| Kong X Z, He Q S, Yang B, et al. 2017. Hydrological regulation drives regime shifts: evidence from paleolimnology and ecosystem modeling of a large shallow Chinese lake[J]. Global Change Biology, 23(2): 737-754. DOI:10.1111/gcb.13416 |
| 雷瑾, 史小丽, 张民, 等. 2019. 白洋淀超微真核藻的空间分布特征及关键影响因子[J]. 湖泊科学, 31(6): 1559-1569. |
| 李永刚, 胡庆杰, 曲疆奇, 等. 2018. 北京密云水库底栖动物群落结构及其时空变化[J]. 水生态学杂志, 39(5): 31-38. |
| 李云虎, 刘秉良. 2013. 白洋淀湿地生态环境现状与保护对策[J]. 江苏农业科学, 41(10): 350-353. |
| 李艳利, 李艳粉, 徐宗学. 2015. 影响浑太河流域大型底栖动物群落结构的环境因子分析[J]. 环境科学, 36(1): 94-106. |
| 刘丹, 等. 2019. 水文连通性对湖泊生态环境影响的研究进展[J]. 长江流域资源与环境, 28(7): 1702-1715. |
| 梁红, 黄林培, 陈光杰, 等. 2018. 滇东湖泊水生植物和浮游生物碳、氮稳定同位素与元素组成特征[J]. 湖泊科学, 30(5): 1400-1412. |
| 李娜, 周绪申, 孙博闻, 等. 2020. 白洋淀浮游植物群落的时空变化及其与环境因子的关系[J]. 湖泊科学, 32(3): 772-783. |
| 刘世存, 王欢欢, 田凯, 等. 2020. 白洋淀生态环境变化及影响因素分析[J]. 农业环境科学学报, 39(5): 1060-1069. |
| 刘祥, 陈凯, 陈求稳, 等. 2016. 淮河流域典型河流夏秋季底栖动物群落特征及其与环境因子的关系[J]. 环境科学学报, 36(6): 1928-1938. |
| 刘鑫, 史斌, 孟晶, 等. 2020. 白洋淀水体富营养化和沉积物污染时空变化特征[J]. 环境科学, 41(5): 2127-2136. |
| 刘月英, 张文珍, 王跃先, 等. 1979. 中国经济动物志: 淡水软体动物[M]. 北京: 科学出版社. |
| 芦康乐, 杨萌尧, 武海涛, 等. 2020. 黄河三角洲芦苇湿地底栖无脊椎动物与环境因子的关系研究-以石油开采区与淡水补给区为例[J]. 生态学报, 40(5): 1637-1649. |
| 鲁如坤. 2000. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社. |
| 陆强, 陈慧丽, 邵晓阳, 等. 2013. 杭州西溪湿地大型底栖动物群落特征及与环境因子的关系[J]. 生态学报, 33(9): 2803-2815. |
| Ma M Y, Liu J L, Wang X M. 2011. Biofilms as potential indicators of macrophyte-dominated lake health[J]. Ecotoxicology, 20(5): 982-992. DOI:10.1007/s10646-011-0677-2 |
| Marcello B, Luciana M, Francesca P. 2017. Depth-related response of macroinvertebrates to the reversal of eutrophication in a Mediterranean lake: Implications for ecological assessment[J]. Science of the Total Environment, 579: 456-465. DOI:10.1016/j.scitotenv.2016.11.073 |
| Margalef D R. 1958. Information theory in ecology[J]. General Systems, 3: 36-71. |
| 孟云飞, 崔恩慧, 鲁甲, 等. 2019. 辽河流域太子河大型底栖动物群落与水环境因子关联性的量化分析[J]. 湖泊科学, 31(6): 1637-1650. |
| Morse J C, Yang L F, Tian L X. 1994. Aquatic Insects of China Useful for Monitoring Water Quality[M]. Nanjing: Hohai University Press. |
| 其木乐, 李宁, 白露超, 等. 2020. 湟水河上游底栖动物群落结构及其与环境因子间的关系[J]. 南京农业大学学报, 43(1): 72-79. |
| Pedro M, Leunda J O, Rafael M, et al. 2008. Longitudinal and seasonal variation of the benthic macroinvertebrate community and biotic indices in an undisturbed Pyrenean river.[J]. Ecological Indicators, 9(1): 52-63. |
| Penning W E.2012.Ecohydraulics in large shallow lakes: implications for management[D].Netherlands: Delft University of Technology |
| Rana J S, Semalty B, Singh P, et al. 2017. Checklist of benthic macroinvertebrate taxa along different riparian land use types in Alaknanda River Catchment of the Central Himalaya, Uttarakhand.(India)[J]. Proceedings of the Zoological Society, 11: 1-24. |
| 任海庆, 袁兴中, 刘红, 等. 2015. 环境因子对河流底栖无脊椎动物群落结构的影响[J]. 生态学报, 35(10): 3148-3156. |
| Shannon C E. 1949. The mathematical theory of communication[J]. The Mathematical Theory of Communication: 29-125. |
| 邵勇, 王洪杨, 徐蛟, 等. 2020. 滆湖入湖河流春季大型底栖动物群落结构及水质生物学评价[J]. 生态学杂志, 39(5): 1617-1628. |
| 沈洪艳, 曹志会, 刘军伟, 等. 2015. 太子河流域大型底栖动物功能摄食类群与环境要素的关系[J]. 中国环境科学, 35(2): 579-590. |
| 沈月含, 丁森, 王慧, 等. 2020. 滦河秋季大型底栖动物群落结构空间格局与关键环境影响因子识别[J]. 环境科学研究, 33(10): 2298-2307. |
| 史键, 赵文, 李赫, 等. 2013. 汤河水库和大伙房水库底栖动物群落结构、底层鱼类产量及水质生态评价[J]. 大连海洋大学学报, 28(4): 383-389. |
| Solis M, Mugni H, Hunt L, et al. 2016. Land use effect on invertebrate assemblages in Pampasic streams(Buenos Aires, Argentina)[J]. Environmental Monitoring and Assessment, 188(9): 539. DOI:10.1007/s10661-016-5545-3 |
| 宋晓薇, 赵侣璇, 谢祎敏, 等. 2019. 湖泊生态系统服务功能价值估算及保护对策研究[J]. 环境科学与管理, 44(1): 162-166. |
| Strayer D, Dudgeon D. 2010. Freshwater biodiversity conservation: recent progress and future challenges[J]. Journal of the North American Benthological Society, 29(1): 344-358. DOI:10.1899/08-171.1 |
| 孙枭琼, 李宏昌, 廖晨延, 等. 2019. 白洋淀底栖无脊椎动物调查及水环境分析[J]. 河北渔业, (8): 40-44+50. |
| Tang T, Cai Q, Liu J. 2006. Using epilithic diatom communities to assess ecological condition of Xiangxi River system[J]. Environmental Monitoring and Assessment, 112(1/3): 347-361. DOI:10.1007%2Fs10661-006-7666-6 |
| Tang C H, Yi Y J, Yang Z F, et al. 2018. Effects of ecological flow release patterns on water quality and ecological restoration of a large shallow lake[J]. Journal of Cleaner Production, 174: 577-590. DOI:10.1016/j.jclepro.2017.10.338 |
| Terbraak C J F. 1986. Canonical correspondence analysis: a new eigenvector method for multivariate direct gradient analysis[J]. Ecology, 67(5): 1167-1179. DOI:10.2307/1938672 |
| Tesgaye T, Sheppard D, Islam K, et al. 2006. Development of chemical index as a measure of instream water quality in response to land-use and land cover changes[J]. Water, Air&Soil Pollution, 174: 161-179. |
| 田世民, 王兆印, 徐梦珍, 等. 2010. 底栖动物与水生栖息地面积耦合机制研究[J]. 人民黄河, 32(11): 19-20+22+152. |
| Vanni M J. 2002. Nutrient cycling by 2animals in freshwater ecosystems[J]. Annual Review of Ecology and Systematics, 33: 341-370. DOI:10.1146/annurev.ecolsys.33.010802.150519 |
| Wagner C E, Harmon L J, Seehausen O. 2014. Cichlid species-area relationships are shaped by adaptive radiations that scale with area[J]. Ecology Letters, 17(5): 583-592. DOI:10.1111/ele.12260 |
| 汪敬忠, 刘卓, 魏浩, 等. 2020. 白洋淀表层沉积物元素的空间特征、风险评价及来源分析[J]. 环境科学, 41(1): 224-231. |
| 王丑明, 张屹, 石慧华, 等. 2016. 洞庭湖大型底栖动物群落结构和水质评价[J]. 湖泊科学, 28(2): 395-404. |
| 王东升, 门彬, 张美一. 2020. 论浅水湖泊中的水固交错带与科学清淤规划-以雄安新区白洋淀为例[J]. 环境科学学报, 40(5): 1550-1559. |
| 王欢欢, 白洁, 刘世存, 等. 2020. 白洋淀近30年水质时空变化特征[J]. 农业环境科学学报, 39(5): 1051-1059. |
| 王琰, 童春富, 汤琳, 等. 2020. 崇明东滩盐沼湿地大型底栖动物功能群分布特征及其影响因子[J]. 生态学杂志, 39(03): 880-892. |
| 王银东, 熊邦喜, 陈才保, 等. 2005. 环境因子对底栖动物生命活动的影响[J]. 浙江海洋学院学报(自然科学版), (3): 60-64+87. |
| 吴红斌, 田自强. 2017. 流域景观格局对大型底栖动物群落结构的影响研究-以太子河山区段为例[J]. 环境保护科学, 43(4): 95-101. |
| Wu Y H, Liu E F, Yao S C, et al. 2010. Recent heavy metal accumulation in Dongjiu and Xijiu Lakes, East China[J]. Journal of Paleolimnology, 43(2): 385-392. DOI:10.1007/s10933-009-9337-4 |
| 相晨, 严力蛟, 韩轶才, 等. 2019. 千岛湖生态系统服务价值评估[J]. 应用生态学报, 30(11): 3875-3884. |
| 谢松, 黄宝生, 王宏伟, 等. 2010. 白洋淀底栖动物多样性调查及水质评价[J]. 水生态学杂志, 31(1): 43-48. |
| Xu M Q, Zhu J, Huang Y Y, et al. 1998. The ecological degradation and restoration of Baiyangdian Lake, China[J]. Journal of Freshwater Ecology, 13(4): 433-446. DOI:10.1080/02705060.1998.9663640 |
| 许亚红, 郑子叶, 王建柱, 等. 2019. 高岚河大型底栖动物时空分布及影响因子研究[J]. 生物资源, 41(5): 426-433. |
| Yan S J, Xuan W, Zhang Y L, et al. 2020. A hybrid PCA-GAM model for investigating the spatiotemporal impacts of water level fluctuations on the diversity of benthic macroinvertebrates in Baiyangdian Lake, North China[J]. Ecological Indicators: 116. |
| 闫欣, 牛振国. 2019. 1990-2017年白洋淀的时空变化特征[J]. 湿地科学, 17(4): 436-444. |
| 杨柳燕, 杨欣妍, 任丽曼, 等. 2019. 太湖蓝藻水华暴发机制与控制对策[J]. 湖泊科学, 31(1): 18-27. |
| 杨青瑞, 陈求稳. 2010. 漓江大型底栖无脊椎动物及其与水环境的关系[J]. 水利水电科技进展, 30(6): 8-10+38. |
| 杨薇, 孙立鑫, 王烜, 等. 2020. 生态补水驱动下白洋淀生态系统服务演变趋势[J]. 农业环境科学学报, 39(05): 1077-1084. |
| Yang W, Yang Z F. 2014. Effects of long-term environmental flow releases on the restoration and preservation of Baiyangdian Lake, a regulated Chinese fresh[J]. Hydrobiologia, 730: 79-91. DOI:10.1007/s10750-014-1823-7 |
| Yang Y, Zhou X, Yi Y, et al. 2018. Influence of debris flows on macroinvertebrate diversity and assemblage structure[J]. Ecological Indicators Integrating Monitoring Assessment & Management, 85: 781-790. |
| 杨雨风, 易雨君, 周扬, 等. 2019. 白洋淀底栖动物群落影响因子研究[J]. 水利水电技术, 50(2): 21-27. |
| Yang Y F, Zhou X D, Yi Y J, et al. 2018. Influence of debris flows on macroinvertebrate diversity and assemblage structure[J]. Ecological Indicators, 85: 781-790. DOI:10.1016/j.ecolind.2017.11.008 |
| Yang Y F, Yi Y J, Zhou Y, et al. 2020. Spatio-temporal variations of benthic macroinvertebrates and the driving environmental variables in a shallow lake[J]. Ecological Indicators, 110: 105948. DOI:10.1016/j.ecolind.2019.105948 |
| 姚敏奇. 2019. 白洋淀流域湿地生态安全评价及生态补偿研究[D]. 保定: 河北大学 |
| 尹子龙, 吴沛沛, 胡晓东. 2020. 长荡湖底栖动物群落结构及与环境因子关系[J]. 江苏水利, (2): 10-15. |
| Yi Y, Sun J, Yang Y, et al. 2018. Habitat suitability evaluation of a benthic macroinvertebrate community in a shallow lake[J]. Ecological Indicators, 90: 451-459. DOI:10.1016/j.ecolind.2018.03.039 |
| 虞左明, 蔡飞. 1997. 西湖引水治理前后底栖动物群落的比较研究[J]. 浙江大学学报(理学版), 24(1): 93-94. |
| 张欢, 肖协文, 王玉玉, 等. 2015. 鄱阳湖流域饶河鱼类稳定同位素比值和营养级的空间变化[J]. 湖泊科学, 27(6): 1004-1010. |
| Zhang X X, Yi Y J, Yang Z F. 2020. Nitrogen and phosphorus retention budgets of a semiarid plain basin under different human activity intensity[J]. Science of the Total Environment, 703: 134813. DOI:10.1016/j.scitotenv.2019.134813 |
| 张璐璐. 2013. 基于底栖-浮游耦合食物网的湖泊生态模型研究[D]. 北京: 北京师范大学 |
| 张素珍, 田建文, 李贵宝. 2007. 白洋淀湿地面临的生态问题及生态恢复措施[J]. 水土保持通报, (3): 146-150. |
| 张宇航, 张敏, 张海萍, 等. 2018. 北京河流大型底栖动物空间格局及其环境响应研究[J]. 生态毒理学报, 13(4): 101-110. |
| 朱金峰, 周艺, 王世新, 等. 2019. 1975年-2018年白洋淀湿地变化分析[J]. 遥感学报, 23(5): 971-986. |
| 朱利明, 肖文胜, 周东, 等. 2019. 淀山湖大型底栖动物群落结构及其与环境因子的关系[J]. 水生态学杂志, 40(2): 55-65. |
