 , 陈惠鑫1, 亓畅1, 侯锋2, 曹效鑫2, 郑祥1,4
, 陈惠鑫1, 亓畅1, 侯锋2, 曹效鑫2, 郑祥1,4 , 曾凡刚1, 李振华3
, 曾凡刚1, 李振华31. 中国人民大学环境学院, 北京 100872;
2. 国投信开水环境投资有限公司, 北京 100084;
3. 北京市昌平区水务局, 北京 102200;
4. 北京市可持续城市排水系统构建与风险控制工程技术研究中心, 北京 100044
收稿日期: 2020-09-17; 修回日期: 2020-12-18; 录用日期: 2020-12-18
基金项目: 国家水体污染控制与治理科技重大专项(No.2017ZX07102-002);国家自然科学基金(No.52070192,51778618)
作者简介: 程荣(1981-), 女, E-mail:chengrong@ruc.edu.cn
通讯作者(责任作者): 郑祥, E-mail:zhengxiang@ruc.edu.cn
曾凡刚, E-mail:zengfg@ruc.edu.cn
摘要:北运河是北京市最重要的排水河道,近年来流域综合治理工作使得其生态环境逐步得到改善.为掌握北运河生态环境的改善状况,本研究于2018年5月—2019年7月基于北运河上游重要节点的沙河水库开展了水生态调查,并通过文献调研,系统对比了近年来北运河水质改善状况.实地调查结果发现,沙河库区及其上下游(以下简称为沙河)浮游植物的主要组成为绿藻门(Chlorophyta)和硅藻门(Bacillariophyta),Shannon-Wiener多样性指数表明沙河水质处于中污染状态;沙河浮游动物主要组成部分为原生动物和轮虫,浮游动物丰度的季节性差异明显.冗余分析结果显示,化学需氧量与浮游生物丰度显著相关(p < 0.01).基于文献调研的结果表明,北运河的水质经历了"污染-污染加剧-污染遏制"的过程,且北运河的污染治理工作在2015年已见成效,浮游生物的变化也佐证了这一过程.但目前沙河化学需氧量和总氮、总磷浓度仍然无法达到水功能区划的要求.因此,未来可通过加强非点源污染的治理、强化污水处理厂脱氮处理工艺等措施进一步改善北运河污染状况.
关键词:北运河浮游植物浮游动物演变特征环境因子
Characteristics of plankton community structure in Shahe Reservoir of the North Canal
CHENG Rong1
, CHEN Huixin1, QI Chang1, HOU Feng2, CAO Xiaoxin2, ZHENG Xiang1,4, ZENG Fangang1, LI Zhenhua31. School of Environment and Natural Resources, Renmin University of China, Beijing 100872;
2. SDIC Xinkai Water Environment Investment Co., Ltd, Beijing 100084;
3. Water Authority of Changping District, Beijing 102200;
4. Engineering Research Center of Sustainable Urban Sewage System Construction and Risk Control, Beijing 100044
Received 17 September 2020; received in revised from 18 December 2020; accepted 18 December 2020
Abstract: The North Canal is the most important drainage channel in Beijing. In order to understand the improvement of the ecological environment of the North Canal, an aquatic ecological survey was performed, from May 2018 to July 2019, in Shahe Reservoir, an important node in the upper North Canal. Based on the literature survey, recent improvement of water quality in the North Canal was systematically assessed. It was found that Chlorophyta and Diatom were the main components of phytoplankton in upstream, within, and downstream of Shahe Reservoir (henceforth referred to as the Shahe River). Bashed on Shannon-Wiener Diversity Index, the water quality of the Shahe River was relatively poor. The zooplankton was dominated by Protozoa and Rotifer, and the seasonality of abundance of zooplankton was markable. The results of redundancy analysis (RDA) showed a significant correlation between COD and plankton abundance. The Shahe River has experienced the process of pollution onset, pollution intensification and pollution containment, and the pollution control measures, applied in the North Canal has achieved preliminary results since 2015. Changes in plankton are also the evidence of this process. However, COD, TN and TP of the Shahe River still cannot meet the requirements of appropriate water function zoning. Therefore, the pollution control measures such as non-point source pollution control and nitrogen removal from sewage may be applied to further improve water quality in the North Canal.
Keywords: Northern Canalphytoplanktonzooplanktonevolution characteristicsenvironmental factors
1 引言(Introduction)北运河是北京市人口最集中、产业最聚集、城市化水平最高的流域, 承担着北京中心城区90%的排水任务(许晓伟等, 2009).由于流域沿线排污口多、水利基础设施滞后等因素, 2008年北运河水系达标河段长度百分比仅为17.6%, 在五大水系中居于末位(方升华等, 2010; 北京市环境保护局, 2009).2009年, 《北运河流域水系综合治理规划(2009—2015年)》全面实施, 北运河流域开展了水资源保护体系建设、水资源配置体系建设、防洪减灾体系建设;2013年, 北京市连续实施两个三年行动计划, 不断提升污水处理率, 北运河流域的水环境质量也逐步得到改善, 到2019年Ⅰ~Ⅳ类水体的河段长度占比超60%.但水质管理的目标并非是单一的水体污染物, 基于水生态系统安全的管理强调从生态系统健康的角度进行管理(孟伟等, 2015).因此, 从水生态的角度客观评价北运河水环境质量变化状况对于保障北运河生态系统健康、实现北运河流域的科学和可持续发展具有重要的意义.
浮游生物群落在流域生态系统中占有重要地位, 它们在维持水环境生态平衡中发挥着重要作用, 也是水体富营养化的重要指示生物(Natalie et al., 2016; Hansson et al., 2007).浮游植物和浮游动物作为水生态系统的初级生产者和食物链中的重要纽带, 是水生态系统物质循环和能量流动的基础(Zhang et al., 2019; 王爱爱等, 2014).国内****针对北运河水系浮游生物的多样性调查及河流健康评价已有相关报道.例如, 高彩凤等(2012)在2009—2010年对浮游植物的调查发现, 北运河水系浮游植物表现为蓝藻-绿藻型, 基于浮游植物的水质评价结果为中~重污染;王汨等(2020)在2015年对北运河水系浮游植物进行了调查, 基于浮游植物的水质评价结果表明北运河水系处于中度污染的状态;顾晓昀等(2018)在2015年针对北运河(北京段)开展的生态系统健康评价中指出, 北运河生态系统的健康状况总体较差.然而, 以上研究仅仅监测和评价了当季/当年北运河水系浮游生物及水质的变化状况, 缺乏针对北运河流域长期浮游生物变化的系统研究.在大时间尺度范围内分析北运河浮游生物的变化, 也有助于为北运河下一步治理工作提供科学依据.
因此, 本文基于实地调查, 聚焦于2018—2019年北运河沙河水库浮游植物和浮游动物的种类、密度和物种多样性的动态变化, 研究沙河典型浮游生物群落结构特征与环境因子的关系, 分析现阶段影响浮游生物群落的主要影响因素;同时, 基于文献调研, 分别对比系列工程实施前的2006年、工程实施中和实施后的2018—2019年中浮游生物的群落结构组成, 以明确北运河浮游生物的变迁, 进而为今后北运河水体的科学管理和可持续发展提供理论依据.
2 材料与方法(Materials and methods)2.1 采样区概况和采样设置沙河水库位于北京市昌平区, 总面积约1.8 km2, 水位约为36m(孙文等, 2019), 上连东沙河、北沙河、南沙河, 下连温榆河, 是北运河上游的重要节点, 也是北运河源头区的主要调蓄水库.作为典型的非常规水源补给型水体, 沙河水库再生水补给量占总入库量的70%~80%(北京市人民政府, 2016).本研究选择沙河水库及其上下游区段作为调查区域, 采样点包括沙河水库、北沙河、南沙河, 其中, S10、S4分别为南、北沙河入库口采样点.沙河采样点示意图见图 1, 采样点坐标见表 1.
图 1(Fig. 1)
 |
| 图 1 沙河采样点示意图 Fig. 1Sketch of sampling sites in Shahe River |
表 1(Table 1)
| 表 1 采样点分布 Table 1 Distribution of sampling sites | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 1 采样点分布 Table 1 Distribution of sampling sites
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
自2018年5月以来, 沙河水库开始低水位运行模式, 库区水深由约7 m下降近2 m, 水流速度增加.为掌握低水位运行状态对水生态的改善情况, 本研究设置生态监测时间为2018年5月—2019年7月, 共6次采样.季节划分按照《气候季节划分》(QX/T152—2012), 将2018年9月划分为秋季, 2018年11月19日滑动平均气温小于10 ℃, 因此, 将2018年11月23日的调查列为冬季;2018年和2019年的5月与7月分别划分为春季与夏季.
2.2 监测方法2.2.1 浮游植物取表层水和水面下0.5 m处水样处各5 L, 混合均匀后取混合水样1 L, 加鲁哥氏液现场固定.水样带回实验室, 静置48 h后虹吸除去上清液, 将每个样品浓缩至30 mL.使用光学显微镜(OLYMPUS, CX31)在0.1 mL计数室中对浓缩的样品进行计数(孟伟等, 2011).
2.2.2 浮游动物浮游动物中枝角类和桡足类定量样品的采集方法同浮游植物, 轮虫类和甲壳类定量样品为采集表层水和水面下0.5m处水样各5 L, 混合后通过25号浮游生物网(网孔0.064mm)过滤、浓缩至50mL保存, 现场固定后在显微镜下计数(孟伟等, 2011; 金相灿等, 1990).
2.2.3 水质分析将于4 ℃冰箱中保存的1 L样品用于水质参数的测定, 其中, 化学需氧量(COD)采用快速消解分光光度法测定, 水样中的总磷(TP)采用钼酸铵分光光度法测定, 总氮(TN)采用过硫酸钾氧化紫外分光光度法测定, 氨氮(NH4+-N)采用低量程水杨酸法测定, 具体测定方法参照《地表水环境质量标准(GB3838—2002) 》.
2.3 数据处理分析方法采用Shannon-Wiener多样性指数描述浮游生物群落特征并对水质状况进行评价, 其计算公式为:
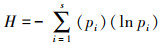 | (1) |
对浮游生物密度和理化数据进行log10(x+1)转换, 使用Canoco5对浮游生物群落与环境因子进行冗余分析(RDA), 当排序轴梯度最大值小于3时选择RDA对浮游动物与环境因子间的关系进行进一步分析.采用SPSS 22.0软件中的Pearson分析进行数据相关性分析, 使用Origin 2018软件进行数据处理和制图.
3 结果与讨论(Results and discussion)3.1 浮游植物群落变化本次调查在沙河共鉴定出浮游植物7门144种, 其中, 绿藻门(Chlorophyta)和硅藻门(Bacillariophyta)在浮游植物群落组成中占主导地位(图 2).类似的调查结果也出现在其他以再生水补给为主的河流和湖泊中(Lv et al., 2020; 陈红等, 2011).因此, 在再生水补给的水体中, 绿藻门和硅藻门是浮游植物常见的物种组成.沙河浮游植物的细胞密度呈现出季节性变化:在秋季达到最大值为8.27×103 cells·mL-1;秋季至冬季, 浮游植物的细胞密度逐渐降低, 11月达到最小值8.87×102 cells·mL-1;随着春季温度的上升, 浮游植物的细胞密度又开始增加.这可能是由于浮游植物的生长受到温度的影响, 9月的温度((24.0±1.5) ℃)处于藻类, 特别是绿藻((25.7±0.1) ℃)和硅藻((24.0±0.4) ℃)生长的最佳条件, 使得沙河9月浮游植物的密度处于最大值(Nalley et al., 2018).这一变化规律与Li等(2019)的结果一致.除季节变化外, 水位波动被认为是控制浮游植物生物量、物种多样性等的重要因素(de Emiliani, 1997).2018年5月沙河水库开始人工干预下的低水位运行, 浮游植物物种数由77种(5月)增加为84种(7月), 2019年5月达最大值106种.这可能是因为水位变化通过影响水体透明度、营养条件等间接影响浮游植物生长(Wu et al., 2019; Dembowska et al., 2017).此外, 由于水位下降, 流速增加, 破坏了蓝藻的生长环境, 使得蓝藻的细胞密度由4.96×103 cells·mL-1(2018年5月)下降为3.15×103 cells·mL-1(2018年7月), 在2019年5月蓝藻细胞密度下降为1.79×103 cells·mL-1(马健荣等, 2013).
图 2(Fig. 2)
 |
| 图 2 浮游植物群落组成和浮游植物细胞密度的季节动态 Fig. 2Seasonal dynamics of various phytoplankton community composition and total phytoplankton cell density |
低水位运行后, 沙河各采样点浮游植物Shannon-Wiener多样性指数的变化见图 3.Shannon-Wiener物种多样性指数的数值越大, 表明浮游植物群落结构越复杂, 对环境的反馈功能越强.由图 3可知, 沙河浮游植物的多样性指数在1.40~2.22之间, 表明沙河水质目前处于中污染状态.其中, 北沙河S3样点与南沙河昌平区界处S13样点的多样性指数最高, 这可能是因为在北沙河种植有荷花等大型水生植物.水生植物能够吸收和存贮营养物质, 一定数量的水生植物能够抑制浮游植物的生长, 进而提高生态系统的水生生物多样性, 使得水生态系统结构趋于稳定(李娣等, 2014).北沙河入库口处S4样点的多样性指数最小, 这可能是因为在北沙河入库口处有再生水厂的退水流入, 由于处理工艺的限制, 再生水通常具有较高的总氮(Lv et al., 2020; Borchardt, 1994).而营养物质浓度的变化会直接影响浮游植物的组成和丰度, 使得S4处水生态系统结构趋于不稳定.
图 3(Fig. 3)
 |
| 图 3 各采样点处Shannon-Wiener多样性指数 Fig. 3Shannon-Wiener biodiversity index at different sampling points |
3.2 浮游动物群落变化沙河13个监测点共鉴定出浮游动物4类81种, 其中, 原生动物的种类数最多, 共36种;轮虫次之, 共30种;枝角类和桡足类相对较少(图 4).该群落结构组成与国内一些河流湖泊的组成相似(王海邻等, 2019; 杨亮杰等, 2014).这是由于原生动物和轮虫类较枝角类和桡足类, 繁殖发育速度快, 能够快速适应水环境的变化, 因此, 在短期内能达到很高的丰度.基于密度的变化, 原生动物占浮游动物总丰度的81%以上.浮游动物密度在秋季(2018年9月)达到峰值为1.0×104 ind.·L-1, 在冬季(2018年11月)出现最小值为1.0×103 ind.·L-1, 最大值与最小值之间相差数倍, 季节性差异明显.沙河浮游动物密度在秋季达最大值, 这可能是由于浮游动物能够以较小的浮游植物等为食(Karlson et al., 2009), 2018年9月浮游植物种类多、生物量大, 进而为浮游动物的生长、繁殖提供了稳定的饵料来源, 使得9月浮游动物的密度达到最大.此外, 在沙河实施低水位运行后, 浮游动物的物种数出现增加, 由45种(2018年5月)增加至58种(2018年7月), 在2019年5月达最大值73种;而种群密度自2018年9月后出现下降, 2019年5月(5.55×103 ind.·L-1)、7月(2.95×103 ind.·L-1)的种群密度均值较2018年相应月份均出现下降.研究表明, 如果出现浮游动物种类增加、种群密度下降, 表明水体污染程度下降(杨苏文, 2015).这也表明低水位运行有助于沙河生态环境的改善.
图 4(Fig. 4)
 |
| 图 4 浮游动物群落组成和浮游动物细胞密度的季节动态 Fig. 4Seasonal dynamics of various zooplankton community composition and total phytoplankton cell density |
沙河浮游动物密度的空间变化见图 5.浮游动物密度最大值出现在南沙河S11处(均值为1.26×104 ind.·L-1), 北沙河S2处浮游动物密度最小(2.86×103 ind.·L-1), 且呈现出南沙河(8.18×103 ind.·L-1)>北沙河(6.55×103 ind.·L-1)>沙河水库(4.57×103 ind.·L-1)的特征.以浮游动物的数量作为水体营养程度的判断指标时, 当浮游动物密度>3×103 ind.·L-1时, 水体为富营养(林海等, 2019).浮游动物的监测结果表明沙河存在富营养化的问题, 且南沙河的富营养化程度高于北沙河.这可能是由于生活污水是南沙河污染的主要来源, 氮、磷含量高, 氮、磷营养盐又通过渗透和离子调节机制促进了浮游动物的生长, 使得南沙河浮游动物密度最高, 污染严重(崔驰飞等, 2019; 陈清等, 2012).
图 5(Fig. 5)
 |
| 图 5 各采样点处浮游动物数量 Fig. 5Number of zooplankton at different sampling points |
3.3 北运河水环境的演变特征由表 2可知, 北运河沙河水库及其上下游的水质经历了污染-污染加剧-污染遏制的过程.在2006年, 库区除COD外, 其余指标均超地表水Ⅴ类标准, 达不到功能区划的要求, 其中以TN污染最为严重, 超地表水Ⅴ类标准11.2倍(华振玲等, 2010).在随后的2009—2010年, 污染持续加重, 除出现TN污染外, COD、TP的污染也加剧, 入库河流呈现出南沙河的污染程度高于北沙河的状况(高彩凤等, 2012).近年来, 在开展了相关污染治理工作后, 北运河治理已初见成效.各项水质指标均呈下降趋势, 但COD和TN、TP仍无法达到功能区划的要求, TN成为北运河沙河水库上下游的主要污染因子(余昊翔等, 2020;表 2).在空间分布上, 仍呈现出南沙河污染严重于北沙河的状况.调查研究指出, 降雨地表径流、雨污河流管网溢流和污水处理厂排水是引发北运河水质不达标的主要原因(荆红卫等, 2013;杨勇等, 2012).随着相关工程的实施, 南沙河、北沙河的补水主要来源于再生水厂退水(北京市人民政府, 2016).然而由于处理工艺的限制, 再生水中通常具有较高的TN(Lv et al., 2020).朱利英等(2020)的研究也指出, 沙河库区下游温榆河的水质污染物转向了TN.因此, 除了通过加强雨污管网溢流等非点源污染的治理、恢复库岸带植被等措施缓解外, 也要强化污水处理厂脱氮处理工艺.
表 2(Table 2)
| 表 2 北运河上游水质演变 Table 2 Evolution of water environment quality in the North Canal | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 2 北运河上游水质演变 Table 2 Evolution of water environment quality in the North Canal
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
由表 3可知, 北运河上游浮游植物的物种组成经历了绿藻型为主-绿藻、蓝藻型为主-绿藻为主, 其次为硅藻-绿藻门和硅藻门为主的变化过程.在2006年, 沙河水库绿藻门的优势种群小球藻(Chlorella)、蓝藻门的颤藻(Oscillatoria)、硅藻门的舟形藻(Navicula)均为富营养化水体的指示种(华振玲等, 2010);在2009—2010年, 北运河蓝藻门和绿藻门共占物种组成的34.1%, 硅藻门占17.1%, 且蓝藻主要出现在夏季, 其余季节均以绿藻为主(高彩凤等, 2012);2015年春、夏、秋季中, 硅藻门成为仅次于绿藻门的物种组成(王汨等, 2020);2018—2019年, 硅藻门和绿藻门二者的物种组成达60%以上.且在秋季, 沙河硅藻门的物种组成(22种)超过绿藻门(19种).已有研究表明, 在未受污染的河流中, 硅藻为主要的浮游植物种群;在河流受到污染后, 从以硅藻占优势转变为以丝状绿藻占优势(冯天翼等, 2011);硅藻门物种比例较高的往往是污染较轻的河段, 而蓝、绿藻种类比较高的则为污染较重的河段(王爱爱等, 2014).因此, 从浮游植物物种组成反映出北运河流域污染状况已得到减轻.除物种组成的变化外, 藻类的平均密度也出现了下降, 由2006年的4767.44×104 cells·L-1下降为525.9×104 cells·L-1(本研究, 2018—2019年).Shannon-Wiener多样性指数表明, 2009—2010年, 大部分断面为中污染, 部分为重污染;2015年的调查结果表明北运河水体处于中度污染(除冬季);本研究的结果也表明, 近年来北运河水质处于中污染状态.北运河浮游动物物种数也出现增加:2009—2010年北运河浮游动物的调查结果显示(李学军等, 2014), 北运河水系共有浮游动物79种, 且主要由一些耐污种组成;而2018—2019年仅沙河段物种数就有81种.此外, 基于浮游生物的变化过程发现, 北运河的水质状况在相关工程实施的2009—2010年已有所改善, 在2015—2019年, 浮游植物的物种多样性增加, 污染得到缓解.
表 3(Table 3)
| 表 3 北运河上游浮游生物演变 Table 3 Evolution of plankton in the North Canal | |||||||||||||||||||||||||
表 3 北运河上游浮游生物演变 Table 3 Evolution of plankton in the North Canal
| |||||||||||||||||||||||||
3.4 环境因子对浮游生物群落的影响由污染引起的环境因子变化是河湖生态系统中浮游生物群落变化的主要驱动因素, 但与浮游生物变化相关的关键环境因子在不同水体中有所不同(Xiong et al., 2019; Peng et al., 2018; 姜会超等, 2015).RDA排序图结果显示(图 5), 前两个轴的特征值分别为0.524和0.062, 共解释了浮游生物群落变异程度的61.1%.其中, COD与浮游生物的细胞密度显著正相关(r=0.414, p=0.006), 说明水体中的有机物会促进浮游生物的生长繁殖.COD整体上反映了水体中有机质的含量水平, 有机质也可以为浮游生物提供更多的食物来源(林青等, 2014);水体中有机物会促进浮游植物的生长繁殖, 同时浮游植物自己会通过光合作用产生大量有机物, 使水体COD升高(刘洋等, 2018).此外, 当浮游植物数量增加时, 浮游动物摄食量也增加, 排泄物增多, 使得水体中有机质增多, COD增加(黄良民等, 1994).王汨等(2020)在2015年的调查中指出, TP、TN和CODMn是影响北运河浮游植物的环境因子;郭飞飞等(2016)对湖北金沙河水库的研究表明, PO43--P和COD是影响浮游动物群落结构的主要环境因子.此外, TP与硅藻密度也具有显著相关性(r=0.664, p < 0.05).这是因为PO43--P易被藻类利用, 从而有利于硅藻的生长繁殖(刘霞等, 2012).因此, 库区除了控制有机物外, 也要加强营养盐的控制.
图 6(Fig. 6)
 |
| 图 6 沙河主要浮游生物群落和环境因子的冗余分析 Fig. 6Redundancy analysis of main plankton communities and environmental factors in Shahe River |
4 结论(Conclusions)1) 北运河流域沙河水库及其上下游浮游植物的组成以绿藻门和硅藻门为主, 基于浮游植物多样性指数的评价结果表明, 沙河水库目前处于中污染状态;浮游动物以原生动物和轮虫为主, 浮游动物的细胞密度季节差异明显, 冬季最低.
2) 北运河流域沙河水库及其上下游的水质经历了污染-污染加剧-污染遏制的过程;浮游植物的物种组成经历了绿藻型为主-绿藻、蓝藻型为主-绿藻为主, 其次为硅藻-绿藻门和硅藻门为主的变化过程.尽管北运河的水质状况在2015年得到缓解, 但沙河水质仍无法达到功能区划的要求.
3) RDA分析结果显示, COD和浮游生物的细胞密度显著正相关, 表明水体中的有机物会促进浮游生物的生长繁殖.
参考文献
| 北京市环境保护局.2009.2008北京市环境状况报告[R].北京: 北京市环境保护局 |
| 北京市人民政府.2016.北京市"十三五"时期水务发展规划[R].北京: 北京市人民政府 |
| Borchardt M A. 1994. Effects of flowing water on nitrogen and phosphorus-limited photosynthesis and optimum N:P ratios by Spirogyra Fluviatilis[J]. Journal of Phycology, 30(3): 418-430. DOI:10.1111/j.0022-3646.1994.00418.x |
| 陈红, 魏彤宇, 张良运, 等. 2011. 北方城市景观河道浮游藻类群落结构特征[J]. 湖北农业科学, 50(12): 2435-2437. DOI:10.3969/j.issn.0439-8114.2011.12.018 |
| 陈清, 白龙. 2012. 北京市南沙河海淀段水环境现状分析与评价[J]. 北京水务, (5): 37-39. DOI:10.3969/j.issn.1673-4637.2012.05.014 |
| 崔驰飞, 祁诗月, 郝明, 等. 2019. 城市黑臭水体治理技术研究-以北京市海淀区南沙河流域为例[J]. 北京水务, (2): 12-16. |
| de Emiliani M O G. 1997. Effects of water level fluctuations on phytoplankton in a river-floodplain lake system (Paraná River, Argentina)[J]. Hydrobiologia, 357(1): 1-15. |
| Dembowska E A, Kubiak-Wójcicka K. 2017. Influence of water level fluctuations on phytoplankton communities in an oxbow lake[J]. Fundamental and Applied Limnology/Archiv für Hydrobiologie, 190(3): 221-233. DOI:10.1127/fal/2017/0998 |
| 方升华, 白文荣, 刘泽娟, 等. 2010. 北运河流域管理体制与机制探讨[J]. 北京水务, (5): 22-24. DOI:10.3969/j.issn.1673-4637.2010.05.008 |
| 冯天翼, 宋超, 陈家长. 2011. 水生藻类的环境指示作用[J]. 中国农学学报, 27(32): 257-265. |
| 高彩凤, 李学军, 毛战坡. 2012. 北运河浮游植物调查及水质评价[J]. 水生态学杂志, (2): 85-90. |
| 顾晓昀, 徐宗学, 刘麟菲, 等. 2018. 北京北运河河流生态系统健康评价[J]. 环境科学, 39(6): 2576-2587. |
| 郭飞飞, 张云, 赵广, 等. 2016. 金沙河水库浮游动物群落结构及其与环境因子的关系[J]. 生态学杂志, 35(8): 2208-2216. |
| Hansson L, Gustafsson S, Rengefors K, et al. 2007. Cyanobacterial chemical warfare affects zooplankton community composition[J]. Freshwater Biology, 52(7): 1290-1301. DOI:10.1111/j.1365-2427.2007.01765.x |
| 华振玲, 杜桂森, 武佃卫, 等. 2010. 北京温榆河浮游藻类与水质分析[J]. 世界科技研究与发展, 32(2): 213-215. DOI:10.3969/j.issn.1006-6055.2010.02.028 |
| 黄良民, 钱宏林, 李锦蓉. 1994. 大鹏湾赤潮多发区的叶绿素a与环境关系初探[J]. 海洋与湖沼, 25(2): 197-205. DOI:10.3321/j.issn:0029-814X.1994.02.015 |
| 姜会超, 陈海刚, 宋秀凯, 等. 2015. 莱州湾金城海域浮游动物群落结构及与环境因子的关系[J]. 生态学报, 35(22): 7308-7319. |
| 金相灿, 屠清瑛. 1990. 湖泊富营养化调查规范[M]. 北京: 中国环境科学出版社, 239-246. |
| 荆红卫, 张志刚, 郭婧. 2013. 北京北运河水系水质污染特征及污染来源分析[J]. 中国环境科学, 33(2): 319-327. DOI:10.3969/j.issn.1000-6923.2013.02.019 |
| Karlson A M L, Viitasalo-Frosen S. 2009. Assimilation of 14C-labelled zooplankton benthic eggs by macrobenthos[J]. Journal of Plankton Research, 31(4): 459-463. DOI:10.1093/plankt/fbn131 |
| Li C, Feng W, Chen H, et al. 2019. Temporal variation in zooplankton and phytoplankton community species composition and the affecting factors in Lake Taihu-a large freshwater lake in China[J]. Environmental Pollution, 245: 1050-1057. DOI:10.1016/j.envpol.2018.11.007 |
| 李娣, 李旭文, 牛志春, 等. 2014. 太湖浮游植物群落结构及其与水质指标间的关系[J]. 生态环境学报, 23(11): 1814-1820. DOI:10.3969/j.issn.1674-5906.2014.11.014 |
| 林海, 王源, 李冰. 2019. 北京市妫水河浮游动物群落结构与水质评价[J]. 生态学报, 39(20): 7583-7591. |
| 林青, 由文辉, 徐凤洁, 等. 2014. 滴水湖浮游动物群落结构及其与环境因子的关系[J]. 生态学报, 34(23): 6918-6929. |
| 刘霞, 陆晓华, 陈宇炜. 2012. 太湖浮游硅藻时空演化与环境因子的关系[J]. 环境科学学报, 32(4): 821-827. |
| 刘洋, 吕俊平, 刘琪, 等. 2018. 太原汾河蓄水区浮游植物细胞密度及其与营养元素的关系[J]. 生态学报, 38(3): 991-1002. |
| Lv X, Zhang J, Liang P, et al. 2020. Phytoplankton in an urban river replenished by reclaimed water:Features, influential factors and simulation[J]. Ecological Indicators, 112: 106090. DOI:10.1016/j.ecolind.2020.106090 |
| Nalley J O, O'Donnell D R, Litchman E. 2018. Temperature effects on growth rates and fatty acid content in freshwater algae and cyanobacteria[J]. Algal Research, 35: 500-507. DOI:10.1016/j.algal.2018.09.018 |
| 马健荣, 邓建明, 秦伯强, 等. 2013. 湖泊蓝藻水华发生机理研究进展[J]. 生态学报, 33(10): 3020-3030. |
| 孟伟, 范俊韬, 张远. 2015. 流域水生态系统健康与生态文明建设[J]. 环境科学研究, 28(10): 1495-1500. |
| 孟伟, 张远, 渠晓东, 等. 2011. 河流生态调查技术方法[M]. 北京: 科学出版社, 92-97. |
| Natalie L, Sarah C W, Brandon J C, et al. 2016. Nitrogen sources and net growth efficiency of zooplankton in three Amazon River plume food webs[J]. Limnology and Oceanography, 61: 460-468. DOI:10.1002/lno.10227 |
| Peng H, Xiong W, Zhan A. 2018. Fine-scale environmental gradients formed by local pollutants largely impact zooplankton communities in running water ecosystems[J]. Aquatic Biology, 27: 43-53. DOI:10.3354/ab00695 |
| 孙文, 王理明, 刘吉宝, 等. 2019. 北运河沙河水库沉积物营养盐分布特征及其溯源分析[J]. 环境科学学报, 39(5): 1581-1589. |
| 王爱爱, 冯佳, 谢树莲. 2014. 汾河中下游浮游藻类群落特征及水质分析[J]. 环境科学, 35(3): 915-923. |
| 王海邻, 刘玉飞, 任玉芬, 等. 2019. 北京市河流秋季浮游动物群落特征分析[J]. 环境科学, 40(8): 3568-3576. |
| 王汨, 杨柏贺, 马思琦, 等. 2020. 北运河水系浮游植物功能群与水体环境因子的关系[J]. 水产学杂志, 33(2): 70-76. |
| Wu Z, Liu J, Huang J, et al. 2019. Do the key factors determining phytoplankton growth change with water level in China's largest freshwater lake?[J]. Ecological Indicators, 107: 105675. DOI:10.1016/j.ecolind.2019.105675 |
| Xiong W, Ni P, Chen Y, et al. 2019. Biological consequences of environmental pollution in running water ecosystems:A case study in zooplankton[J]. Environmental Pollution, 252: 1483-1490. DOI:10.1016/j.envpol.2019.06.055 |
| 许晓伟, 刘德文, 车洪军, 等. 2009. 北运河水环境调查与评价[J]. 海河水利, (2): 14-15. DOI:10.3969/j.issn.1004-7328.2009.02.007 |
| 杨亮杰, 吕光汉, 竺俊全, 等. 2014. 横山水库浮游动物群落结构特征及水质评价[J]. 水生生物学报, 38(4): 720-728. |
| 杨苏文. 2015. 滇池、洱海浮游动植物环境图谱[M]. 北京: 科学出版社, 352-394. |
| 杨勇, 魏源送, 郑祥, 等. 2012. 北京温榆河流域微生物污染调查研究[J]. 环境科学学报, 32(1): 9-18. |
| 余昊翔, 杨东翰, 谢梦飞, 等. 2020. 北运河流域沙河水库水质时空变化特征[J]. 环境化学, 39(10): 2849-2857. DOI:10.7524/j.issn.0254-6108.2019101804 |
| 朱利英, 陈媛媛, 刘静, 等. 2020. 温榆河水环境质量与浮游植物群落结构的时空变化及其相互关系[J]. 环境科学, 41(2): 702-712. |
