 , 陈诚2
, 陈诚2 , 董建玮2, 王智源2, 毛媛媛3, 陈求稳1,2
, 董建玮2, 王智源2, 毛媛媛3, 陈求稳1,21. 重庆交通大学河海学院, 重庆 400074;
2. 南京水利科学研究院生态环境研究中心, 南京 210029;
3. 江苏省水利工程规划办公室, 南京 210029
收稿日期: 2019-03-11; 修回日期: 2019-05-05; 录用日期: 2019-05-08
基金项目: 国家自然科学基金(No.51709179);江苏省水利科技项目(No.2017012);中央级公益性科研院所基金资助项目(No.Y918011)
作者简介: 何欣霞(1997-), 女, E-mail:hexinxia_xx@163.com
通讯作者(责任作者): 陈诚, E-mail:chencheng@nhri.cn
陈求稳, E-mail:qwchen@nhri.cn
摘要: 以江苏省里下河腹部地区38个典型湖泊湖荡为研究对象,分析了春季浮游植物群落结构特征及其与环境因子的关系,运用了基于理化指标的综合营养状态指数(TLI)评价方法和基于浮游植物群落结构分析的生物指标(Shannon-Wiener多样性指数H'和Pielou均匀度指数J)评价方法,评价了湖泊湖荡营养状态.结果显示共采集到里下河腹部地区湖泊湖荡浮游植物7门50属89种,主要隶属于绿藻门(Chlorophyta)和硅藻门(Bacillariophyta);浮游植物细胞丰度介于2.3×105~3.7×107 cells·L-1,生物量介于0.05~15.02 mg·L-1,绿藻门与隐藻门(Cryptophyta)在细胞丰度上占优势,隐藻门在生物量上占优势;梅尼小环藻(Cyclotella meneghiniana)、啮蚀隐藻(Cryptomonas erosa)、尖尾蓝隐藻(Chroomonas acuta)、四尾栅藻(Scenedesmus guadricauda)为主要优势种;生物指标评价方法的结果显示湖泊湖荡总体为轻度富营养,与TLI评价方法的结果有较好一致性,表明两类评价方法在里下河腹部地区具有较强内在关联性.
关键词:里下河腹部地区湖泊湖荡群落结构环境因子营养状态
Phytoplankton community structure and trophic status of lakes in the Lixia River hinterland of Jiangsu Province in spring
HE Xinxia1,2
, CHEN Cheng2 , DONG Jianwei2, WANG Zhiyuan2, MAO Yuanyuan3, CHEN Qiuwen1,2 1. College of Hehai, Chongqing Jiaotong University, Chongqing 400074;
2. Center for Eco-Environmental Research, Nanjing Hydraulic Research Institute, Nanjing 210029;
3. Planning Office for Hydraulic Engineering, Jiangsu Province, Nanjing 210029
Received 11 March 2019; received in revised from 5 May 2019; accepted 8 May 2019
Abstract: This study selected 38 typical lakes in the Lixia River hinterland of Jiangsu Province to analyze the phytoplankton community structure in spring and its relationship to the hydro-environmental factors. Meanwhile, the trophic status of the lakes was assessed through the trophic level index (TLI) method based on physicochemical indicators as well as the biological indicators method based on the Shannon-Wiener diversity index (H') and the Pielou evenness index (J). A total of 89 phytoplankton species were identified in all lakes, which belongs to 50 different genera and 7 phyla. Chlorophyta and Bacillariophyta were two dominated phyla in the communities. The cell abundance of phytoplankton ranged from 2.3×105 to 3.7×107 cells·L-1, in which Chlorophyta and Cryptophyta were dominant. The biomass of phytoplankton ranged from 0.05 to 15.02 mg·L-1, in which Cryptophyta was dominant. The dominant species included Cyclotella meneghiniana, Cryptomonas erosa, Chroomonas acuta and Scenedesmus guadricauda. The evaluation results based on biological indicators demonstrate that the lakes are in light eutrophication status, which is consistent with the results analyzed with TLI method. It reveals that there is a strong inherent relevance between two evaluation methods for the lakes of the Lixia River hinterland.
Keywords: the Lixia River hinterlandlakescommunity structureenvironmental factorstrophic status
1 引言(Introduction)湿地是分布于陆生生态系统与水生生态系统之间的自然生态系统, 对维持生态平衡、保护物种多样性和调节气候等有不可代替的生态作用.湖泊湖荡作为湿地的一种典型类型, 具有洪水调蓄、水源涵养、水质净化和生境维持等生态功能.随着社会经济的快速发展和人类活动的不断加剧, 湖泊湖荡面临自由水面面积减小、生物多样性降低和富营养化程度加剧等环境问题.湖泊湖荡水生态健康研究已成为全球关注热点之一(邱阳凌等, 2018), 而科学地评价湖泊湖荡水生态健康是水生态治理的重要前提.
水体营养状态是评价水生态健康的重要部分.目前, 对于湖泊湖荡营养状态的评价, 国内外研究者多采用综合营养状态指数(TLI)评价方法(Burns et al., 1999; 张思冲等, 2003), 由于地域差异, 其评价结果具有一定局限性(Xu et al., 2010; 张蕊等, 2013).浮游植物作为水生态系统重要的初级生产者, 能有效表征水体营养状态(梁伟林, 2017).研究表明, 浮游植物群落结构(细胞丰度、生物量、Shannon-Wiener多样性指数(H′)和Pielou均匀度指数(J))可作为生物指标评价水体营养状态(王雨等, 2010; 梁伟林, 2017; 邱阳凌等, 2018), 然而其评价结果与理化指标评价结果的一致性尚有待检验(徐成斌, 2006; 邱阳凌等, 2018), 亟需揭示并建立湖泊湖荡营养状态理化指标和生物指标评价结果的内在关系, 从而支撑湖泊湖荡生态修复.
本文以江苏省苏北平原里下河腹部地区38个典型湖泊湖荡为研究对象, 开展水质监测和浮游植物调查, 分析春季浮游植物群落结构特征及其与环境因子的关系, 运用基于理化指标的TLI评价方法和基于浮游植物群落结构分析的生物指标评价方法, 分别评价湖泊湖荡营养状态, 并分析两者之间的一致性, 为平原河网地区水生态健康评价和水生态治理提供科学支撑.
2 材料与方法(Materials and methods)2.1 研究区域与采样点布设里下河腹部地区位于江苏省中部, 地处32° ~34° N, 119°~120° E(图 1), 东临通榆河, 西贴里运河, 北起苏北灌溉总渠, 南至新通扬运河.地势四周高、中间低, 呈蝶形洼地, 水系复杂, 湖泊湖荡众多且分散, 属复杂平原河网地区(毛媛媛等, 2015).年平均气温为14~15 ℃, 年降雨量为500~1400 mm, 汛期为6—9月, 非汛期为10月—次年5月, 属亚热带温润季风气候(张贤芳, 2012).该地区由射阳湖、绿草荡、大凹子圩等41个零星分散的湖泊湖荡与圩区组成, 总面积为695 km2.
图 1(Fig. 1)
 |
| 图 1 里下河腹部地区湖泊湖荡及采样点分布 Fig. 1Locations of the sampling sites and lakes in the Lixia River hinterland |
本次调查于2018年4月22—4月28日进行38个典型湖泊湖荡水样与浮游植物样品采集, 采样点布设严格参照《水环境检测规范》(SL219—2013), 遵循湖泊湖荡全覆盖的原则, 同时根据各湖泊湖荡面积、形态、污染程度及开发利用等情况增设采样点, 最终共布设48个采样点(图 1).
2.2 样品采集与测定方法2.2.1 样品采集水样采集遵照《地表水和污水检测技术规范》(HJ/T91-2002), 并优先选择自由水面进行采集.湖水荡洗采水器与水样容器2~3次, 利用采水器汲取水面下0.5 m处水体(王瑜等, 2011), 装入水样容器中, 并加入固定剂和贴好瓶身标签, 保存于冰盒, 带回实验室检测.每个采样点需设置1~2个平行样以避免单次采样的偶然性.
浮游植物定量样品采集依照《水环境检测规范》(SL219—2013), 使用2.5 L有机玻璃采水器, 分别采集表、中、底层水样于塑料桶中(邱阳凌等, 2018), 混合均匀后取1 L水加15 mL鲁戈氏液固定, 经筒型分液漏斗静置沉淀48 h, 吸取上清液, 保留30 mL浓缩样品待检.
2.2.2 测定方法利用便携式YSI-6600型多参数水质检测仪现场测定pH值、电导率、溶解氧、温度, 其余指标依据《地表水和污水检测技术规范》(HJ/T91—2002)进行测定.
浮游植物种类测定时, 吸取0.1 mL水样于载玻片, 制成临时装片于显微镜下观察, 每瓶标本至少计数两片.根据形态结构归入大门类, 再依据《浮游植物手册》(Hasle et al., 1978)和《中国淡水藻类—系统、分类及生态》(胡鸿钧等, 2006)进行属种鉴定.浮游植物细胞丰度采用样方法, 浮游植物生物量采用细胞体积法(Hasle et al., 1978).
2.3 评价方法计算优势度(Y)并判断浮游植物优势种;由于Shannon-Wiener多样性指数(H′)和Pielou均匀度指数(J)均能反映湖泊湖荡营养状态(成芳等, 2010), 两者数值越大, 水质越好, 水体富营养化程度越低(程军明等, 2015), 所以将H′、J作为湖泊湖荡营养状态评价的生物指标;选取叶绿素a(Chla)、总氮(TN)、总磷(TP)、高锰酸盐指数(CODMn)、透明度(SD)共5个理化指标, 分别运用生物指标评价方法和TLI评价方法, 依据湖泊湖荡营养状态等级划分的标准(表 1)(成芳等, 2010; 梁伟林, 2017; 邱阳凌等, 2018), 评价湖泊湖荡营养状态, 并进行一致性检验;采用R语言corrplot函数程序包对6种浮游植物群落指标(细胞丰度、生物量及主要优势种群啮蚀隐藻、尖尾蓝隐藻、四尾栅藻和梅尼小环藻)与TN、TP、磷酸根(PO43-)、溶解氧(DO)、Chla、CODMn及SD共7个环境因子进行相关性分析.本文均以湖泊湖荡为基本评价单元, 具体计算公式如下(张蕊等, 2013; 孟顺龙等, 2015):
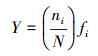 | (1) |
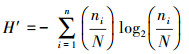 | (2) |
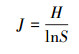 | (3) |
 | (4) |
| 表 1 湖泊湖荡营养状态等级划分 Table 1 Trophic status classification of lakes | ||||||||||||||||||||||||
表 1 湖泊湖荡营养状态等级划分 Table 1 Trophic status classification of lakes
| ||||||||||||||||||||||||
式中, ni为湖泊湖荡中第i个种群的个数, N为同一湖泊湖荡中所有种群的总个数, S为该湖泊湖荡中种类的总数, fi为同一湖泊湖荡中第i个种群出现的频率, 当Y≥0.02时确定其为优势种(邱阳凌等, 2018).TLI(∑)为综合营养状态指数, Wj为第j种参数的营养状态指数相关权重, TLI(j)为第j种参数的营养状态指数.
3 结果分析(Results)3.1 浮游植物群落结构特征分析3.1.1 浮游植物种类组成里下河腹部地区共采集到浮游植物7门50属89种.由图 2所示里下河腹部地区湖泊湖荡春季浮游植物种类组成可知, 浮游植物种类组成特征整体为绿藻门(Chlorophyta)>硅藻门(Bacillariophyta)>裸藻门(Euglenophyta)>蓝藻门(Cyanophyta)>隐藻门(Cryptophyta)>甲藻门(Pyrrophyta)>金藻门(Chrysophyta), 绝大多数湖泊湖荡浮游植物以绿藻门和硅藻门为主.
图 2(Fig. 2)
 |
| 图 2 里下河腹部地区湖泊湖荡春季浮游植物种类组成 Fig. 2Species composition of phytoplankton for lakes in the Lixia River hinterland in spring |
3.1.2 浮游植物细胞丰度及生物量图 3为里下河腹部地区各湖泊湖荡春季浮游植物细胞丰度与生物量, 图 4为里下河腹部地区湖泊湖荡春季浮游植物相对细胞丰度和生物量.浮游植物细胞丰度介于2.3×105~3.7×107 cells·L-1, 均值为2.6×106 cells·L-1, 生物量介于0.05~15.02 mg·L-1, 均值为1.32 mg·L-1, 各湖泊湖荡细胞丰度和生物量空间差异显著.绿藻门与隐藻门在细胞丰度上占显著优势, 且相对细胞丰度占比共达80%以上;甲藻门相对生物量最大, 但其生物量极大值仅在个别湖泊湖荡出现, 隐藻门相对生物量仅次于甲藻门, 且分布于绝大多数湖泊湖荡, 因此隐藻门在生物量上占主要优势.
图 3(Fig. 3)
 |
| 图 3 里下河腹部地区各湖泊湖荡春季浮游植物细胞丰度与生物量 Fig. 3Cell abundance and biomass of phytoplankton for lakes in the Lixia River hinterland in spring |
图 4(Fig. 4)
 |
| 图 4 里下河腹部地区湖泊湖荡春季浮游植物相对细胞丰度与生物量 Fig. 4Relative cell abundance and biomass of phytoplankton for lakes in the Lixia River hinterland in spring |
3.1.3 浮游植物优势种表 2为里下河腹部地区各湖泊湖荡优势种及其优势度.本次调查共发现10种优势种, 分别为硅藻门中的变异直链藻(Melosira varians)、谷皮菱形藻(Nitzchia palea)、异极藻(Gomphonema)、梅尼小环藻;绿藻门中的小球藻(Chlorella)、二形栅藻(Scenedesmus dimorphus)、四尾栅藻;隐藻门中的啮蚀隐藻、尖尾蓝隐藻;甲藻门中的裸甲藻(Gymnodinium aerucyinosum Stein), 其中隐藻门、绿藻门、硅藻门、甲藻门为优势种的湖泊湖荡数目分别为21个、11个、4个和1个, 东荡、王庄荡、大纵湖、郭正湖未发现优势种.依据优势种在湖泊湖荡的分布情况可知, 绿藻门四尾栅藻、隐藻门啮蚀隐藻和尖尾蓝隐藻为主要优势种, 而主要浮游植物硅藻门表现为1种优势种对应1个湖泊湖荡, 其中梅尼小环藻的优势度最高(Y=0.081), 因此梅尼小环藻同样为主要优势种.
表 2(Table 2)
| 表 2 里下河腹部地区各湖泊湖荡优势种及优势度 Table 2 Distribution and dominance of dominant species in lakes of the Lixia River hinterland | |||||||||||||||||||||||||||||||||
表 2 里下河腹部地区各湖泊湖荡优势种及优势度 Table 2 Distribution and dominance of dominant species in lakes of the Lixia River hinterland
| |||||||||||||||||||||||||||||||||
3.2 浮游植物群落多样性分析图 5为里下河腹部地区各湖泊湖荡春季浮游植物生物多样性的分析结果, 多样性指数(H′)介于0.75~3.77, 均值为2.64;均匀度指数(J)介于0.24~0.88, 均值为0.67.总体而言, H′与J空间分布较为一致, 表现为H′较高的湖泊湖荡, 其J也较高.
图 5(Fig. 5)
 |
| 图 5 里下河腹部地区湖泊湖荡春季浮游植物多样性指数与均匀度指数 Fig. 5The diversity index and evenness index of phytoplankton for lakes in the Lixia River hinterland in spring |
3.3 湖泊湖荡营养状态理化指标与生物指标评价图 6为里下河腹部地区各湖泊湖荡营养状态评价的空间分布结果, 其中图 6a为基于理化指标的TLI方法评价结果, 其结果显示湖泊湖荡整体为轻度-重度富营养, 绝大多数为轻度富营养;图 6b和6c分别为运用生物指标H′和J的评价结果, 其结果显示湖泊湖荡整体为中营养-重度富营养, 绝大多数为轻度富营养.运用TLI和H′评价营养状态时有29个湖泊湖荡营养状态评价结果一致(图 6a和6b);运用TLI和J评价营养状态时有25个湖泊湖荡营养状态评价结果一致(图 6a和6c).同时H′和J评价结果的空间分布较为一致, 表现为H′评价显示营养状态较高的湖泊湖荡, J评价显示其营养状态也较高, 但J评价结果较TLI与H′评价结果而言个别湖泊湖荡营养状态略低.总体而言, 66%以上的湖泊湖荡营养状态理化指标和生物指标评价结果完全一致, 且均呈轻度富营养.此外, 对TLI评价结果与生物指标评价结果进行相关性分析, 其结果显示TLI与H′和J的相关性系数分别为-0.26和-0.32, 具有一定相关性, 统计分析结果也表明理化指标和生物指标的评价结果具有较好一致性.
图 6(Fig. 6)
 |
| 图 6 里下河腹部地区各湖泊湖荡营养状态评价 Fig. 6The trophic status evaluation of lakes in the Lixia River hinterland |
3.4 浮游植物群落指标与环境因子的相关性分析图 7为浮游植物群落指标和环境因子的相关性结果(蓝色为正相关, 红色为负相关, 圆圈越大颜色越深表示相关系数越大).由图 7可知, TN(r=0.38)、TP(r=0.49)与梅尼小环藻呈显著正相关, 与H′和J均呈负相关;CODMn与细胞丰度呈显著正相关(r=0.41), 与H′(r=-0.26)和J(r=-0.30)均呈负相关;Chla与浮游植物生物量相关系数高达0.95, 与细胞丰度呈显著正相关(r=0.42), 与H′(r=-0.35)和J(r=-0.38)均呈显著负相关;SD与生物量相关性尤为显著(r=-0.95), 与Chla呈显著正相关(r=0.42).
图 7(Fig. 7)
 |
| 图 7 浮游植物群落指标与环境因子的Pearson相关系数 Fig. 7The pearson correlation coefficient between phytoplankton community index and environmental factors |
4 讨论(Discussion)4.1 浮游植物群落结构生物多样性是群落结构的重要表征指标, 可用H′与J反映生物多样性特征(陈春亮等, 2016).Ke等(2011)研究表明, H′与J越大, 生物多样性越丰富, 群落结构越复杂稳定.通常以H′大于1.5和J大于0.3作为浮游植物生物多样性较好的标准(孟顺龙等, 2015).本次调查里下河腹部地区湖泊湖荡浮游植物H′均值为2.64, J均值为0.67, 其中92%的湖泊湖荡H′超过1.5, 97%的湖泊湖荡J超过0.3, 说明该地区生物多样性整体较好, 浮游植物群落结构整体较为复杂与稳定.
浮游植物细胞丰度与生物量的差异对生物多样性有一定影响(童琰, 2012; Wang et al., 2014).结合本次浮游植物调查结果与明珠湖4月(细胞丰度均值为2.5×107 cells·L-1, 生物量均值为2.1 mg·L-1, H′均值为2.33, J均值为0.61)(陈立婧等, 2011)浮游植物研究结果对比分析发现, 个别湖泊湖荡浮游植物细胞丰度与生物量较高, 但其对应湖泊湖荡生物多样性较低, 原因可能为在细胞丰度或生物量高值区, 浮游植物种类组成较为单一, 生物多样性反而降低, 例如大凹子圩中硅藻门、绿洋湖中绿藻门的细胞丰度与生物量占比极大, 花粉荡、蜈蚣湖、喜鹊湖中甲藻门的生物量占比极大(图 3), 导致物种组成单一, 对应湖泊湖荡生物多样性下降(图 5).
4.2 湖泊湖荡营养状态环境因子和浮游植物群落结构均能反映湖泊湖荡营养状态(孟顺龙等, 2015).研究表明, 氮磷过于丰富, 会引起某些浮游植物过量繁殖, 数量增加, 进而出现“富营养化”现象(况琪军等, 2005).本次调查湖泊湖荡TN浓度均值为(2.62±0.81) mg·L-1, TP浓度均值为(0.15±0.13) mg·L-1, 采用基于理化指标的TLI方法评价湖泊湖荡营养状态, TN和TP的TLI均值分别高达61.19±8.47和70.00±5.49, 对评价结果影响显著, 表明营养盐过于丰富, TLI方法评价结果显示84%的湖泊湖荡呈轻度富营养, 整体为轻度-重度富营养状态.浮游植物群落结构亦是评价水体营养状态的一种重要手段(程军明等, 2015).依据况琪军等(2005)和和王瑜等(2011)对优势种与营养状态关系的研究表明, 梅尼小环藻、啮蚀隐藻和尖尾蓝隐藻可作为富营养化水体的指示物种, 栅藻适应性强, 能在多种营养类型水体中采集到, 但常在富营养型水体中占优势.本次里下河腹部地区湖泊湖荡浮游植物调查结果显示, 梅尼小环藻、啮蚀隐藻、尖尾蓝隐藻和四尾栅藻为主要优势种, 细胞丰度占比共达66%, 生物量占比共达59%, 在一定程度上限制了生物的多样性.基于浮游植物群落结构分析的生物指标评价(H′和J)结果显示里下河腹部地区76%的湖泊湖荡呈轻度富营养, 且H′和J较里下河腹部地区2014年9月结果(H′均值为3.29, J均值为0.76)偏低(孟顺龙等, 2015), 湖泊湖荡营养状态向富营养化转变.TLI和生物指标两类评价方法显示66%以上的湖泊湖荡营养状态相同且均呈轻度富营养(图 6), 总体具有良好一致性, 且湖泊湖荡营养状态向富营养化转变.
4.3 浮游植物群落结构与环境因子的关系营养盐较高的水体环境能促进浮游植物数量的快速增长(童琰, 2012).由图 7的相关性分析结果可知, 优势种中梅尼小环藻与TN、TP和PO43-具有显著正相关性, 其原因为硅藻门对营养盐含量十分敏感, 在高营养盐水体中, 能快速大量繁殖从而形成优势(Tilman et al., 1986; Sarthou et al., 2005).其余浮游植物与营养盐相关性不显著, 表明对于其余浮游植物而言营养盐含量充足, 不是生长限制条件(Hlaili et al., 2006), 这与本次水质监测结果一致(TN和TP营养盐含量丰富).Chla与浮游植物数量呈显著正相关, CODMn、DO均与细胞丰度呈显著正相关, SD与生物量负相关性尤为显著, 表明随着浮游植物数量的增长, Chla含量增加, 光合作用增强, 水体DO含量增加, 同时死亡腐败的浮游植物数量增加, 水体有机悬浮物含量增加, SD降低, 衡量有机物含量的CODMn也随之增大.
当TN、TP、Chla和CODMn的浓度越高, 而SD越低时, TLI数值越大, 会引起适宜这一水体环境的浮游植物快速生长, 不适宜这一水体环境的浮游植物数量减少, 从而导致生物多样性降低, 富营养化程度增大(程军明等, 2015).图 7的相关性分析结果显示, 理化指标TN、TP、CODMn、Chla与生物指标H′、J均呈负相关, 表明TLI数值越大, 生物多样性越低, 从而使TLI和生物指标方法具有较好内在关联性.本次水质监测结果显示, 里下河腹部地区92%的湖泊湖荡TN浓度和79%的湖泊湖荡TP浓度均超过地表水Ⅴ类标准, 95%的湖泊湖荡CODMn浓度超过地表水Ⅲ类标准(均值为(8.63±4.05) mg·L-1), Chla浓度均值较高((8.90±7.80) μg·L-1), SD均值较低(仅为(0.11±0.07) m), 同时结合表 1湖泊营养状态等级划分结果可知, 其H′和J也相对较低, 表明基于浮游植物的生物评价方法与基于理化指标的TLI评价方法结果一致性较高, 两类评价方法存在显著内在关联性, 研究成果可为湖泊湖荡水生态健康评价提供技术支撑.
对于某一湖泊湖荡而言, 采用不同方法评价营养状态的结果略有差异, 其原因可能为在实际评价中, TLI方法理化指标的选取固定, 未考虑地域差异;生物指标可能受计数方式、鉴定种类与种间竞争关系的影响(邱阳凌等, 2018).同时考虑4月是冬季刚过渡为春季的典型月份, 此时水体营养盐含量丰富, 水温回升, 适宜浮游植物生长, 所以本文重点分析了典型季节湖泊湖荡群浮游植物群落结构和营养状态的空间分布, 后续研究中将对这38个湖泊湖荡其它季节水样和浮游植物的调查数据进行详细分析, 从而全面了解里下河腹部地区湖泊湖荡群的浮游植物群落结构和营养状态时空分布特征.
5 结论(Conclusions)1) 里下河腹部地区湖泊湖荡春季浮游植物生物多样性整体较好, 表明研究区春季浮游植物群落结构整体具有复杂性和稳定性.
2) TLI评价方法和生物指标评价方法显示里下河腹部地区2/3的湖泊湖荡营养状态均为轻度富营养, 两类方法的评价结果具有较好一致性, 湖泊湖荡营养程度呈加剧趋势.
3) 里下河腹部地区湖泊湖荡春季营养盐充足, CODMn、Chla、SD与浮游植物数量相关性显著, 且环境因子与H′、J呈负相关, 表明基于理化指标的TLI评价方法和基于群落结构分析的生物指标评价方法具有较强内在关联性.
参考文献
| Burns N M, Rutherford J C, Clayton J S. 1999. A Monitoring and Classification System for New Zealand Lakes and Reservoirs[J]. Lake and Reservoir Management, 15(4): 255–271.DOI:10.1080/07438149909354122 |
| 陈春亮, 张才学. 2016. 雷州湾浮游植物群落结构特征及其环境影响分析[J]. 应用海洋学学报, 2016, 35(2): 174–182.DOI:10.3969/J.ISSN.2095-4972.2016.02.004 |
| 陈立婧, 吴竹臣, 胡忠军, 等. 2011. 上海崇明岛明珠湖浮游植物群落结构[J]. 应用生态学报, 2011, 22(6): 1599–1605. |
| 成芳, 凌去非, 徐海军, 等. 2010. 太湖水质现状与主要污染物分析[J]. 上海海洋大学学报, 2010, 19(1): 105–110. |
| 程军明, 张曼, 王良炎, 等. 2015. 三种浮游植物指示法在景观性河流水体营养状态评价中的应用[J]. 生态科学, 2015, 34(3): 37–43.DOI:10.3969/j.issn.1671-6361.2015.03.007 |
| Hasle G, Sournia A. 1978. Phytoplankton Manual[M]. Paris: UNESCO. |
| Hlaili A S, Chikhaoui M A, Grami B E, et al. 2006. Effects of N and P supply on phytoplankton in Bizerte Lagoon (western Mediterranean)[J]. Journal of Experimental Marine Biology & Ecology, 333: 79–96. |
| 胡鸿钧, 魏印心. 2006. 中国淡水藻类-系统、分类及生态[M]. 北京: 科学出版社. |
| 况琪军, 马沛明, 胡征宇, 等. 2005. 湖泊富营养化的藻类生物学评价与治理研究进展[J]. 安全与环境学报, 2005, 5(2): 87–91.DOI:10.3969/j.issn.1009-6094.2005.02.024 |
| Ke Z, Huang L, Tan Y, et al. 2011. Plankton community structure and diversity in coral reefs area of Sanya Bay, Hainan Province, China[J]. Biodiversity Science, 19(6): 696–701. |
| 梁伟林. 2017.湖泊富营养化评价方法研究及其系统设计[D].成都: 电子科技大学 |
| 孟顺龙, 肖代, 陈小丽, 等. 2015. 里下河腹地典型水体秋季浮游植物生态学特征[J]. 浙江农业学报, 2015, 27(11): 1998–2005.DOI:10.3969/j.issn.1004-1524.2015.11.22 |
| 毛媛媛, 兰林, 张颖, 等. 2015. 里下河地区河湖水生态保护与修复措施研究[J]. 水论坛, 2015, 3: 1–5. |
| 邱阳凌, 林育青, 刘俊杰, 等. 2018. 淮河干流及主要支流夏季浮游植物群落生物多样性评价[J]. 环境科学学报, 2018, 38(4): 1665–1672. |
| Sarthou G, Timmermans K R, Blain S, et al. 2005. Growth physiology and fate of diatoms in the ocean:a review[J]. Journal of Sea Research, 53(1/2): 25–42. |
| 童琰. 2012.滴水湖浮游植物群落结构动态及其与环境因子关系的研究[D].上海: 华东师范大学 |
| Tilman D, Kiesling R, Sterner R, et al. 1986. Green, blue, green and diatom algae:taxonomic differences in competitive ability for phosphorus, silicon and nitrogen[J]. Archiv für Hydrobiologie, 106(4): 473–485. |
| 王雨, 卢昌义, 谭凤仪, 等. 2010. 深圳红树林水体浮游植物多样性与营养状态评价[J]. 海洋环境科学, 2010, 29(1): 17–22.DOI:10.3969/j.issn.1007-6336.2010.01.004 |
| 王瑜, 刘录三, 舒俭民, 等. 2011. 白洋淀浮游植物群落结构与水质评价[J]. 湖泊科学, 2011, 23(4): 575–580. |
| Wang C Y, Zhou B, Ding X D, et al. 2014. Effects of reclamation on phytoplankton community structure and biodiversity in Jiaojiang Estuary[J]. Journal of East China Normal University, 4: 141–153. |
| 徐成斌. 2006.辽河流域河流水质生物评价研究[D].辽宁: 辽宁大学 |
| Xu Q J, Zheng B H, Zhu Y Z, et al. 2010. Nutrient status evaluation for tributary of Three Gorges Reservoir[J]. China Environmental Science, 30(4): 453–457. |
| 张蕊, 高良敏, 席北斗, 等. 2013. 改进的TLI指数法及其在巢湖营养状态评价中的应用[J]. 环境工程学报, 2013, 7(6): 2127–2133. |
| 张思冲, 张雪萍, 廖永丰. 2003. 营养度指数法在寒地湖泊富营养化评价中的应用[J]. 哈尔滨工业大学学报, 2003, 35(4): 416–419.DOI:10.3321/j.issn:0367-6234.2003.04.009 |
| 张贤芳. 2012.苏北里下河地区末次冰期中晚期气候与环境演变研究[D].南京: 南京师范大学 |
| 中国环境监测总站. 2013. HJ/T91-2002地表水和污水检测技术规范[S].北京: 中国环境科学出版社 |
| 中华人民共和国水利部. 2013.SL219-2013水环境检测规范[S].北京: 中国水利水电出版社 |
