 , 杨飞2, 张毅敏2
, 杨飞2, 张毅敏2 , 朱月明2, 高月香2, 李定龙1
, 朱月明2, 高月香2, 李定龙11. 常州大学环境与安全工程学院, 常州 213164;
2. 环境保护部南京环境科学研究所, 南京 210042
收稿日期: 2017-06-01; 修回日期: 2017-09-15; 录用日期: 2017-09-30
基金项目: 国家水体污染控制与治理科技重大专项(No.2012ZX07101-007)
作者简介: 尹杰(1993—), 男, E-mail: 978815120@qq.com
通讯作者(责任作者): 张毅敏, 女, 研究员, 主要研究流域水污染控制与水体生态修复. E-mail: zym7127@163.com
摘要: 以两种沉水植物马来眼子菜(Potamogeton malainus)和狐尾藻(Myriophyllum spicatum)为研究对象,通过原位模拟试验,比较在相同水位变化幅度下不同高水位持续胁迫水平(分别以T0、T1、T2、T3表示恒静止、持续4 d、持续8 d和持续12 d)对沉水植物的生物量、植株伸长、具体形态指标和光合荧光特性的影响,并同步分析滆湖大洪港内两处风浪差异区(湖岸区M1和湖心区M2)两种沉水植物相应的适应性和形态变化的特征.结果表明,马来眼子菜和狐尾藻都表现出了较好的环境适应性,T0时任意两处的两种植物生物量和株高均最大;在风浪扰动较小区域(M2),持续高水位减弱了两种植物的伸长和生长(p < 0.01),并显著限制其在地上生物量的投资和茎干横向的分支(p < 0.05).在风浪扰动较大区域(M1),持续高水位对两种沉水植物产生了相同的形态变化,并显著降低各梯度下其Fv/Fm和快速光响应曲线(p < 0.05),但M1处两种沉水植物的光合能力均显著大于M2处.狐尾藻对风浪扰动和长时间水位胁迫能表现出更强抗弱光性和光合作用能力.因此,持续高水位胁迫下(小于8 d)两种沉水植物对一定程度水动力仍有较好的形态和生长适应性,对特定水文波动特征进行合理应对以保持沉水植物活力和维持湖泊生态系统具有指导作用.
关键词:马来眼子菜狐尾藻高水位胁迫风浪光合特性
Study on adaptability of two submerged macrophytes for wind-induced wave and high water level stress
YIN Jie1,2
, YANG Fei2, ZHANG Yimin2 , ZHU Yueming2, GAO Yuexiang2, LI Dinglong1 1. School of Environment & Safety Engineering, Changzhou University, Changzhou 213164;
2. Nanjing Institute of Environmental Sciences, Ministry of Environmental Protection of the People's Republic of China, Nanjing 210042
Received 1 June 2017; received in revised from 15 September 2017; accepted 30 September 2017
Supported by the National Major Science and Technology for Water Pollution Control and Treatment(No.2012ZX07101-007)
Biography: YIN Jie(1993—), male, E-mail: 978815120@qq.com
*Corresponding author: ZHANG Yimin, E-mail: zym7127@163.com
Abstract: The variation of two dominant submerged macrophytes, Potamogeton malainus and Myriophyllum spicatum, were studied in-situ under different time of high water level stress (T0, T1, T2 and T3 respectively referred to the initial static, 4 days, 8 days and 12 days). The biomass, elongation, specific morphological index and photosynthetic fluorescence characteristics of two plants had been compared. The characteristics of two plants' corresponding adaptations and morphological changes were synchronically researched at two difference wind-induced wave sites (M1, M2 respectively referred to the lake shore and lake center) in Dahonggang of Gehu Lake. The results showed that the high survival rate of Potamogeton malaianus and Myriophyllum spicatum presented their good adaptability to the surrounding, and the biomass and height of two plants were the largest at either place (M1 and M2) in T0 group. Around area where was disturbed by wind-induced wave slightly (M2), continuous high water level weakened plant growth (p < 0.01) and significantly limited their biomass investment on the ground and stem-lateral branching (p < 0.05). Around area where was disturbed by wind-induced wave dramatically (M1), continuous high water level produced the same morphological changes for both two kinds of submerged macrophytes and significantly decreased their Fv/Fm and rapid light response curve values under each depth gradient (p < 0.05). However, the photosynthetic capacity of two plants at M1 was significantly greater than that of M2. Meanwhile, Myriophyllum spicatum presented more strongly low light adaptability and photosynthetic ability. Therefore, two plants under continuous high water level stress (less than 8 days) showed good adaptability of morphology and growth to certain hydrodynamic conditions. It was instructive to make reasonable response to specific hydrological fluctuation characteristics in order to maintain the vitality of submerged macrophytes and the ecosystem of the lake.
Key words: Potamogeton malainusMyriophyllum spicatumhigh water level stresswind-induced wavephotosynthetic fluorescence characteristics
1 引言(Introduction)沉水植物不仅作为浅水湖泊生态系统的初级生产者, 而且还具有吸收水体营养盐、分泌化感物质、抑制藻类生长繁殖和为其他水生生物提供多样性生境等作用(Zhang et al., 2009;陈开宁等, 2006;刘永等, 2006).同时其生长和分布受多种环境要素的影响, 其中水下光强、营养盐浓度、悬浮物和底质均为重要限制因子(王素梅, 2010;宋玉芝等, 2015).风浪是产生湖泊水动力的主要方式, 它会通过风涌流直接扰动水表以下环境, 也可通过透明度间接影响沉水植物的生长和繁殖(王震等, 2016;王超等, 2014).目前研究结果表明, 水位变动改变了沉水植物光补偿点高度并决定了其在水下可分布的最大深度、光合产量及竞争能力(Xiao et al., 2010;O'Farrell et al., 2011;欧克芳等, 2008;李亚芳, 2015), 且多集中于固定水位梯度、固定水位抬升幅度和固定频率的水位上下波动这几种变化形式上(吴晓东等, 2011;李威等, 2014;Coops and Beklioglu, 2003).以往不少****已经开展了水位对沉水植物生长影响的研究, 但对滆湖实际夏季高水位变化模式的研究较少, 仅有在滆湖现场模拟水位快速抬升对沉水植物生长的影响(高汾等, 2017).此外, 在室内系统模拟研究水动力对沉水植物形态学响应的工作占多数, 这很难反映沉水植物在真实大水体的生长特征.本研究选择生态修复中常见种——马来眼子菜和狐尾藻为研究对象, 在滆湖开展原位实验, 探究水位和风浪的作用对其生理生长和光合荧光特性的影响, 为滆湖及类似湖泊水生植被的恢复及生境条件的控制提供参考.
2 材料与方法(Materials and methods)2.1 研究区概况滆湖位于太湖流域西部, 是苏南太湖湖群的重要组成部分, 水面面积为164 km2, 其地处北亚热带季风气候区, 夏季雨量充沛, 伴随着滆湖富营养化程度的日益加重, 水生植物开始大量消退, 且近年来因夏季连续暴雨导致的水位骤升和持续高水位情况加剧了水生植被的退化.原位试验区位于滆湖东北部大洪港, 该区域平水期常水深为1.6 m(吴淞水位3.2 m), 光照条件好, 并且受鱼类牧食作用小.
通过对该区域2016年夏季(6—9月)的风向频率进行统计再结合历史风向数据, 得到大洪港风向频率玫瑰图, 见图 1, 夏季基本以西南风向为主, 湖面波浪基本顺风聚集到滆湖东北部, 因而在大洪港近岸处会有较多的叠加波且较湖心处会有更明显湖面波动, 所以在大洪港设置2个区域, 近岸处和离岸距离约为1 km的湖心处作为两个风浪差异区进行试验(图 2).
图 1(Fig. 1)
 |
| 图 1 滆湖大洪港风向频率玫瑰图(注:单圈标示该风向出现50频次) Fig. 1Wind frequency rose diagram in Dahonggang |
图 2(Fig. 2)
 |
| 图 2 试验点位和原位试验设计装置图 Fig. 2Experimental site and design of the in-situ experiment |
2.2 试验设计原位试验于M1和M2处同步进行, 试验期基本水环境指标见表 1.马来眼子菜和狐尾藻常被作为富营养湖泊生态修复的主要先锋物种, 且在滆湖仍有分布.因此, 两种沉水植物就近采自滆湖, 择取长势良好的断枝(10 cm长, 顶芽未开花且根部未发育)分别单独移栽到含有15 cm滆湖沉积物基质的塑料桶中(高=30 cm, 直径=25 cm), 每盆15株, 先用塑料桶将植物定植于光照好、水深较浅且不受鱼类牧食的池塘.7 d后将栽有固定生根且萌发一致沉水植物的塑料桶通过尼龙绳以初始0.8 m水深条件(桶上端距水面距离——吴淞水位2.4 m)悬挂固定在可漂浮的种植框架上(3.0 m ×2.5 m, 24盆植物/框架), 同时在框架周围围网以防止鱼类牧食干扰(图 2).以上调绳长降水位(下调绳长升水位)方式调节绳长, 分别设置0.8 m水深恒定不动(T0)、0.6 m/3 d涨水2次, 2 m水深维持4 d, 再以0.6 m/4 d退水2次(T1)、0.6 m/3 d涨水2次, 2 m水深维持8 d, 再以0.6 m/4 d退水2次(T2)、0.6 m/3 d涨水2次, 2 m水深维持12 d, 再以0.6 m/4 d退水2次后(T3), 稳定后以0.8 m一直维持到35 d的4组高水位胁迫梯度(图 3), 每组重复3次, 定期划船于原位调节绳长.M1和M2处采用同样的高水位胁迫试验设计以作为对照.
表 1(Table 1)
| 表 1 试验点基本水环境指标 Table 1 Basic parameters in the water of the test sites | |||||||||||||||||||||||||||||||||
表 1 试验点基本水环境指标 Table 1 Basic parameters in the water of the test sites
| |||||||||||||||||||||||||||||||||
图 3(Fig. 3)
 |
| 图 3 试验中水位波动的幅度 Fig. 3The amplitudes of water-level fluctuations in this experiment |
2.3 参数测定2.3.1 植物生长指标试验于2016年8月19日—9月23日进行, 该时段为全年中水深较高(平均2.2 m)且较为稳定的阶段, 整个试验皆在自然光下进行.试验结束后, 采集两处植物并用水小心清洗去除粘附在根部的泥土后带回实验室.每个梯度选取9株直接测量计数其株高和分枝数并取平均值;植物的地上生物量和地下生物量用其地上部分和地下部分在85 ℃下烘干杀青36 h后的恒重表示, 并计算相对伸长率(RER)和相对生长率(RGR);若最终植株生物量小于试验初始平均值、株高降低(小于10 cm)或地上茎叶基本消失则被认为是未成活组并且不进入生长指标分析中.
2.3.2 植物光合荧光指标叶绿素荧光参数采用Diving-Pam荧光仪(德国WALZ公司)和数据采集软件Wincontrol进行测定.在野外观测现场, 每个梯度上的植物随机选择3片成熟叶片检测荧光强度.于每天早晨6:00左右, 将叶片暗适应5 min后, 打开测量光得到初始荧光值F0, 随后启动饱和脉冲得到最大荧光值Fm, 可得出光系统Ⅱ的最大量子产率(马梦洁等, 2016):
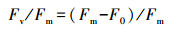 | (1) |
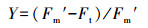 | (2) |
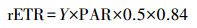 | (3) |
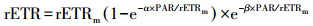 | (4) |
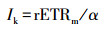 | (5) |
2.4 数据分析通过空白和多重样品平行测定的方法来获得准确数据, 且采用Excel 2007和SPSS 19.0对所得数据进行计算和统计分析处理, 对狐尾藻和马来眼子菜的各指标进行Pearson相关性分析, 图形的绘制则在origin 8.5上完成.
3 结果与分析(Results and analysis)3.1 试验区风浪特征湖泊风浪由两个因素组成, 风生流产生了湖流, 而浪亦代表着波浪, 湖流和波浪是湖泊水动力的主要形式, 通过监测湖泊的风场与波浪要素可以较为直观的表征湖面的风浪特征.由于波浪要素监测于试验前期进行, 为保证风场与波浪要素数据的稳定性和一致性, 本文选用2016年8月11—18日滆湖野外研究基地记录的风速和风向数据来表征试验期滆湖大洪港的风场情况.试验前期监测风速风向, 得到该区域此阶段以西南风为主, 且M1处平均风速为2.18 m·s-1, M2处平均风速2.02 m·s-1, 在风力表(风力等级, GB/T 28591—2012)中均属2级轻风.
波浪数据同样以试验前期波浪的特征值表征试验期波浪要素情况.两处实测风速与波浪特征关系如图 4所示, 可以发现:M1处, 随着风速值上下幅度的变化, 波高值也跟着发生极显著的相关变化(p<0.01), 出现瞬时较剧烈风的同时也会产生较大的波高, 受风速的影响明显;M2处, 风速和波高还是显著相关的(p=0.034), 但风场风速值在8月15日出现过5 m·s-1左右的阵风, 其波高并没有发生显著的上升;两处风速和波高的变化幅度相似, 但M1处的波高随风速变化的显著相关性大于M2处, 差异明显(p<0.05).试验前期两处最大波高值且为0.039 m, 在海况表(海洋调查规范, GB 12763.2-91)属于1级微浪.
图 4(Fig. 4)
 |
| 图 4 M1(a)和M2(b)处实测波高与风速 Fig. 4Measured wave height and wind speed at M1(a) and M2(b) |
3.2 持续高水位胁迫及风浪对沉水植物生长的影响两处沉水植物存活率都很高, 烘后单株干重总生物量都随着高水位胁迫时间的增加而显著减少(p<0.01).一方面, 由图 5a可以得出, M1处的马来眼子菜, 随着水位胁迫时间的增加, T2和T3组相对伸长率和相对生长率均显著下降(表 2, p<0.01), 初始T0时马来眼子菜的相对生长率大于M2处.而M2处, 马来眼子菜均随着高水位胁迫时间的增加, 其相对伸长率和相对生长率均显著减小(p<0.01);另一方面, 由图 5b可以看出, M1处的狐尾藻相对伸长率随胁迫持续开始下降, 但T0、T1和T2胁迫下的差异性不显著(p>0.05), 胁迫持续到第12 d后其相对伸长率才显著减少(p=0.021), 且位于M2处狐尾藻的相对伸长率变化幅度与之相同, 但值均小于M1处.狐尾藻的相对生长率在两处都显著减小, 说明狐尾藻的相对生长率受高水位胁迫有显著影响(p=0.018, p =0.026), 但位于M1处的相对生长率始终大于M2处, 分别为它的1.08、1.01、1.24和1.03倍.
图 5(Fig. 5)
 |
| 图 5 马来眼子菜(a)和狐尾藻(b)相对伸长率和生长率 Fig. 5RER and RGR of Potamogeton malainus (a) and Myriophyllum spicatum (b) |
表 2(Table 2)
| 表 2 不同点位、高水位胁迫及其交互作用对两种沉水植物(马来眼子菜和狐尾藻)的形态特征影响方差分析结果 Table 2 Results of ANOVA of effect of different sites, high water level stress, and their interaction on the morphological parameters of two submerged macrophytes(Potamogeton malainus and Myriophyllum spicatum) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 2 不同点位、高水位胁迫及其交互作用对两种沉水植物(马来眼子菜和狐尾藻)的形态特征影响方差分析结果 Table 2 Results of ANOVA of effect of different sites, high water level stress, and their interaction on the morphological parameters of two submerged macrophytes(Potamogeton malainus and Myriophyllum spicatum)
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
同时受到风浪和持续高水位胁迫后, 观察沉水植物各组织形态的差异性变化特征.从植株种间差异上看, 较试验开始时, 两处试验结束后各胁迫处理组的马来眼子菜和狐尾藻的株高均增加明显(死亡不计数的处理组除外).如表 2所示, 马来眼子菜在水位胁迫下的株高差异显著(F=27.101, p=0.004), 而茎端分枝数之间的差异不显著(F=0.882, p=0.540), 同样地下生物量之间的差异也不显著(F=2.097, p=0.279);而狐尾藻在不同胁迫组下的株高生长量间的差异依旧显著(F=20.898, p=0.007), 茎端分枝数之间的差异与其地下生物量之间的差异也均显著(分别为p =0.004和p=0.003).这一结果说明, 持续高水位胁迫对狐尾藻分枝数和地下生物量的影响比对马来眼子菜其相应组分的影响大.另外, M1、M2处风浪扰动有差异, 分析点位差异和高水位胁迫下其耦合交互作用仅对马来眼子菜分枝数之间有显著差异(p=0.020), 由此可见大洪港实际两处风浪条件对两种沉水植物的形态特征影响有较明显的作用.
3.3 持续高水位胁迫及风浪对沉水植物光合荧光特性的影响试验结束后, 持续高水位胁迫梯度对M1处的马来眼子菜和狐尾藻最大量子产率Fv/Fm值无显著的影响(p>0.05), 其值变化范围仅在0.732~0.823之间(图 6a).而在M2处, 除T0外各梯度不同沉水植物类型对最大光量子产率Fv/Fm影响显著(p<0.05), 且与狐尾藻相比, 马来眼子菜的Fv/Fm的值下降了7.6%~44.1%(图 6b), 其Fv/Fm值也随着胁迫持续时间的增长而减小.单对狐尾藻而言, T3水位梯度的Fv/Fm值分别为T0、T1和T2水位梯度的84.9%、83.7%和83.2%.
图 6(Fig. 6)
 |
| 图 6 M1(a)和M2(b)处马来眼子菜和狐尾藻的最大量子产率 Fig. 6The Fv/Fm of P. malainus and M. spicatum at M1(a) and M2(b) |
如图 7所示, 任意两处两种沉水植物的rETRm、α、Ik值随着深水胁迫梯度的增加均显著减小(p<0.05), 单就马来眼子菜而言, 各胁迫处理组下M1处的rETRm值分别为M2处的1.44、1.08、1.41和1.34倍, 而对狐尾藻其值分别为M2的1.35、1.22、1.97和2.42倍.并且从图中可以看出, 整个试验中位于M1处的狐尾藻在各水位胁迫下都同期拥有最高的rETRm、α和Ik值, 说明与马来眼子菜相比, 在一定风浪条件下(M1处)的狐尾藻无论是否收到高水位持续胁迫作用, 都能保持较好的光合效率和进行光合产物的积累过程.
图 7(Fig. 7)
 |
| 图 7 不同点位处两种沉水植物的快速光响应曲线 Fig. 7The RLCS of different sites of two submerged macrophytes |
4 讨论(Discussion)室内研究沉水植物对单因子胁迫(水质、水位、光照和营养盐)的响应研究较多, 而在野外真实的大湖泊水体下进行原位模拟更具有研究价值和实际借鉴意义, 因为湖泊湖区点位的差异和水动力的构成均会受到水质、风速、波浪、水深(高敏等, 2015)等多种因素的影响, 因此, 考虑讨论在特定风浪和对应实际高水位胁迫耦合作用下, 不同沉水植物所表现出的形态适应性和光合作用能力的差异.
4.1 沉水植物生长对持续高水位和风浪的响应对风场和波浪条件相互作用关系的研究表明, 区域内两者具有高度一致相关性(遐祯等, 2013).在本研究中, 试验区以风速为1.98 m·s-1的西南风为主导, 试验区的风速和波浪分别为2级轻风和1级微浪.同时, 无论风浪大小, 持续高水位对植物的生长影响显著, 其影响表现在两种沉水植物生物量、相对生长率和相对伸长率的减少, 且在水位稳定不变下其株高最高.这个结果与****所得出的(吴晓东, 2012;Zhu et al., 2012), 随着初始水位的增加, 沉水植物把分配各组织器官能量减少并将更多的生物量倾向于株高, 导致相对生长率减少而相对伸长率增加的研究成果并不矛盾, 因为水位的变动已经促使植物的株高伸长, 但由于持续高水位导致的弱光环境使得植物始终满足不了自身光补偿, 从而影响茎继续的伸长和分支, 转而加快对地下生物量的投资.其次, 风浪扰动会通过水动力改变湖泊无机悬浮物影响水体光衰减系数和真光层深度, 这对于沉水植物而言相当重要(何尚卫等, 2014;赵巧华等, 2015).在本研究中, 与风浪较小处相比, 较大的风浪加快减小了马来眼子菜的相对伸长率和相对生长率, 但对狐尾藻的作用不明显.这个结果表明, 狐尾藻对于风浪扰动的抗逆性强于马来眼子菜, 因为马来眼子菜的生态幅较小、易受环境要素影响且常当做清洁种, 而狐尾藻更适氮磷等水质胁迫且耐负荷大(王钦等, 2012;宋玉芝等, 2015;张艳娜等, 2014).
4.2 沉水植物光合荧光特性对持续高水位和风浪的响应在本研究中, 持续高水位胁迫对马来眼子菜的最大量子产量Fv/Fm值显著减小, 其值越小就代表植物的光能转化效率越低, 并且在正常生理状态下绝大多数的C3植物的Fv/Fm在0.80~0.85之间(Song et al., 2015), 而对狐尾藻的影响作用较小则表明其拥有更高的光能利用效率, 所以在水生植被重建的起始阶段, 可以r对策的狐尾藻和菹草为主(徐伟伟等, 2015).同时, 大部分沉水植物会因受到外部作用力后发生机械损伤和生物量的减少(姜义帅, 2013), 但试验结果显示在风浪扰动较大的区域, 两种沉水植物在每个水位胁迫梯度下的Fv/Fm值均要同期大于位于风浪较小的区域, 这是因为试验区的起始风速和波浪水平不高, 仅当为4 m·s-1临界风速以上才以风浪为主导, 对传递到湖面下层的波浪切应力和沉积物再悬浮才有明显影响(耿頔等, 2014;逄勇等, 2008).值得注意的是, 两地沉水植物的快速光响应曲线RLCs及其rETRm、α和Ik值也表现出了与Fv/Fm值相同的变化特征, 如表 3所示, 整体较低的风浪值, 使得位于风浪扰动较大区域其水动力依旧较为明显, 而位于风浪扰动较小区域其近乎等于静水态, 有研究表明当在0~0.01 m·s-1的低流速范围内沉水植物的光合作用率与流速量成正比, 而当流速超过这个阈值或者水流相对静置时光合作用就会受到抑制作用(王华等, 2008), 因此考虑在该种风浪水平下, 风浪的扰动更有助于沉水植物叶片的充分舒展和加大吸收光照的机会, 促进植物的捕光能力和光合效率, 更适合沉水植物生长以及光合作用的进行.
表 3(Table 3)
| 表 3 两种沉水植物的快速光响应曲线参数拟合的结果 Table 3 Parameter values of RLCS of Potamogeton malainus and Myriophyllum spicatum | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 3 两种沉水植物的快速光响应曲线参数拟合的结果 Table 3 Parameter values of RLCS of Potamogeton malainus and Myriophyllum spicatum
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
5 结论(Conclusions)1) 无论风浪程度大小, 持续高水位胁迫均对马来眼子菜和狐尾藻的伸长及生长造成明显的抑制作用, 并且持续时间越长其效果愈明显.并且, 持续弱光环境会使两种沉水植物趋向于减弱顶端生物量的分配和茎端的延伸, 以增加根部的定植恢复能力.
2) 原位风浪扰动的差异会影响不同沉水植物的生长和茎的拉升性能, 风浪扰动较大区域会加快减小马来眼子菜的相对伸长率和相对生长率, 更有甚者, 若同时处于持续高水位胁迫(大于8 d)的弱光环境下会使其衰败死亡.然而风浪差异对狐尾藻的形态作用不明显.
3) 持续高水位胁迫会显著导致两种沉水植物的光合作用效率与能力降低.进一步研究表明处于整体风浪水平低的环境下, 风浪扰动明显区域的水动力条件促进植物对光照的吸收机会, 更有利于沉水植物在弱光环境下对光的捕捉和进行光合作用.同时, 狐尾藻体现出了比马来眼子菜更强光能利用效率.
综上所述, 狐尾藻在形态学响应和光合能力特性上更适合持续高水位的弱光环境及风浪扰动的水动力环境.在定植初期, 选择合适物种适应区域特定的水文和光照条件, 有利于沉水植被在生态修复工程运用上起到更好效果以来恢复退化的浅水湖泊生态系统.
参考文献
| 陈开宁, 包先明, 史龙新, 等. 2006. 太湖五里湖生态重建示范工程——大型围隔试验[J]. 湖泊科学, 2006, 18(2): 139–149. |
| Coops H, Beklioglu M, Crisman T L. 2003. The role of water-level fluctuation in shallow lake cosystems-workshop conclusion[J]. Hydrobiologia, 506-509(1/3): 23–27. |
| 高汾, 张毅敏, 杨飞, 等. 2017. 水位抬升对4种沉水植物生长及光合特性的影响[J]. 生态与农村环境学报, 2017, 33(4): 341–348. |
| 高敏, 刘鑫, 邓建才, 等. 2015. 不同水质对沉水植物马来眼子菜主要生理指标的影响研究[J]. 生态环境学报, 2015, 24(11): 1886–1892. |
| 耿頔, 杨芬, 韦朝阳, 等. 2015. 风浪扰动对太湖水体中砷在水相-悬浮物相之间分配的影响[J]. 环境科学学报, 2015, 35(5): 1358–1365. |
| 何尚卫, 李勇, 赵海光, 等. 2014. 滆湖水体光学性质初步研究[J]. 湖泊科学, 2014, 26(5): 707–712. |
| 姜义帅, 陈灏, 马作敏, 等. 2013. 利用沉水植物生长期收割进行富营养化水体生态管理的实地研究[J]. 环境工程学报, 2013, 7(4): 1351–1358. |
| 李威, 何亮, 朱天顺, 等. 2014. 洱海苦草(Vallisneria natans)水深分布和叶片C、N、P化学计量学对不同水深的响应[J]. 湖泊科学, 2014, 26(4): 585–592. |
| 李亚芳, 陈心胜, 项文化, 等. 2015. 不同高程短尖苔草对水位变化的生长及繁殖响应[J]. 生态学报, 2015, 36(7): 1959–1966. |
| 刘永, 郭怀成, 周丰, 等. 2006. 湖泊水位变动对水生植被的影响机理及其调控方法[J]. 生态学报, 2006, 26(9): 3117–3126. |
| 马梦洁, 张毅敏, 杨飞, 等. 2016. 沉水植物黑藻的拓殖能力和光合荧光特性[J]. 应用与环境生物学报, 2016, 22(5): 0752–0758. |
| O'Farrell, Izaguirre I, Chaparro G, et al. 2011. Water level as the main driver of the alternation between a free-floating plant and a phytoplankton dominated state:a long-term study in a floodplain lake[J]. Aquatic Science, 73(2): 275–287.DOI:10.1007/s00027-010-0175-2 |
| 欧克芳, 林鸿, 陈桂桥, 等. 2008. 沉水植物的特点及其应用[J]. 安徽农业科学, 2008, 36(17): 7210–7221. |
| Platt T, Gallegos C L, Harrison W G. 1980. Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton[J]. Journal of Marine Research (USA), 38: 687–701. |
| 逄勇, 庄巍, 韩涛, 等. 2008. 风浪扰动下的太湖悬浮物实验与模拟[J]. 环境科学, 2008, 29(10): 2743–2748. |
| Ralph PJ, Gademann R. 2005. Rapid light curves:a powerful tool to assess photosynthetic activity[J]. Aquatic Botany, 82: 222–237.DOI:10.1016/j.aquabot.2005.02.006 |
| Ritchie R J, Mekjinda N. 2016. Arsenic toxicity in the water weed Wolffia arrhiza measured using Pulse Amplitude Modulation Fluorometry (PAM) measurements of photosynthesis[J]. Ecotoxicology and Environmental Safety, 132: 178–185.DOI:10.1016/j.ecoenv.2016.06.004 |
| Song X, Zhou G, Xu Z, et al. 2015. Detection of photosynthetic performance of stipa bungeana seedlings under climatic change using chlorophyll fluorescence imaging[J]. Frontiers in Plant Science, 6: 1254–1258. |
| 宋玉芝, 孔繁璠, 王敏, 等. 2015. 光照强度及附植藻类对狐尾藻生理指标的影响[J]. 农业环境科学学报, 2015, 34(2): 233–239. |
| 宋玉芝, 王宇佳, 王锦旗, 等. 2016. 水体氮磷浓度对两种沉水植物上附着藻类的影响[J]. 环境科学学报, 2016, 36(9): 3208–3212. |
| 王超, 张微敏, 王沛芳, 等. 2014. 风浪扰动条件下沉水植物对水流结构及底泥再悬浮的影响[J]. 安全与环境学报, 2014, 14(2): 107–111. |
| 王华, 逄勇, 刘申宝, 等. 2008. 沉水植物生长影响因子研究进展[J]. 生态学报, 2008, 28(8): 3958–3968. |
| 王钦, 何萍, 徐杰, 等. 2012. 北京市河流沉水植物水环境适应性研究[J]. 环境科学学报, 2012, 32(1): 30–36. |
| 王素梅. 2010. 光照和氮、磷的复合作用对苦草生长和生理的影响[D]. 广州: 华南理工大学. 9-10 |
| 王震, 吴挺峰, 邹华, 等. 2016. 太湖不同湖区风浪的季节变化特征[J]. 湖泊科学, 2016, 28(1): 217–224. |
| 吴晓东, 王国祥, 陈正勇, 等. 2011. 水深梯度对黑藻生长的影响[J]. 生态与农村环境学报, 2011, 27(4): 40–45. |
| 吴晓东. 2012. 水位和收割对沉水植物生长的影响研究[D]. 江苏: 南京师范大学. 32-36 |
| Xiao C, Wang X, Xia J, et al. 2010. The effect of temperature, water level and burial depth on seed germination of Myriophyllum spicatum and Potamogeton malaianus[J]. Aquatic Botany, 92(1): 28–32.DOI:10.1016/j.aquabot.2009.09.004 |
| 遐祯, 陶蓉茵, 赵巧华, 等. 2013. 大型浅水湖泊太湖波浪特征及其对风场的敏感性分析[J]. 湖泊科学, 2013, 13(1): 55–64. |
| 徐伟伟, 胡维平, 邓建才, 等. 2015. 菹草生物量控制对群落中沉水植物生长及水质的影响[J]. 生态环境学报, 2015, 24(7): 1222–1227. |
| 张艳娜, 宋玉芝, 王敏, 等. 2015. 附植藻类对沉水植物狐尾藻生长及生理的影响[J]. 环境科学学报, 2015, 35(7): 2282–2288. |
| 赵巧华, 王玲, 杭蓉蓉. 2016. 太湖真光层深度谱的分布特征及其介质影响研究[J]. 环境科学学报, 2016, 36(3): 778–784. |
| Zhang W, Rao P, Zhang H, et al. 2009. The role of diatomite particles in the activated sludge system for treating coal gasification wastewater[J]. Chinese Journal of Chemical Engineering, 17(1): 167–170.DOI:10.1016/S1004-9541(09)60050-1 |
| Zhu G R, Li W, Zhang M, et al. 2012. Adaptation of submerged macrophytes to both water depth and flood intensity as revealed by their mechanical resistance[J]. Hydrobiologia, 696: 77–93.DOI:10.1007/s10750-012-1185-y |
