 ,1,2,3,*, 崔宁1,2
,1,2,3,*, 崔宁1,2Five new species of Arvicolinae and Myospalacinae from the Late Pliocene-Early Pleistocene of Nihewan Basin
ZHENG Shao-Hua1,2, ZHANG Ying-Qi,1,2,3,*, CUI Ning1,2通讯作者: *zhangyingqi@ivpp.ac.cn
收稿日期:2019-05-31网络出版日期:2019-10-20
| 基金资助: |
Corresponding authors: *zhangyingqi@ivpp.ac.cn
Received:2019-05-31Online:2019-10-20

摘要
泥河湾盆地因产出丰富的哺乳动物化石及众多的旧石器地点而久负盛名。为弄清盆地地层的年代范围,近年开展了包括生物地层学、磁性地层学、沉积学、地球化学、孢粉学等多学科的综合研究。目前主要分歧在于对泥河湾层的年代学认识,表现在生物地层学与磁性地层学研究结果不一致。新种化石材料即是从甄选的典型剖面中采集的。?亚科和鼢鼠亚科由于演化趋势明显并且容易量化、演化速率快,对晚新生代生物地层学的研究具有十分突出的意义。鉴于此,对产自泥河湾盆地的?亚科三个新种(泥河湾模鼠Mimomys nihewanensis sp. nov.、简齿松田鼠Pitymys simplicidens sp. nov.、前中华波尔索地鼠Borsodia prechinensis sp. nov.)及鼢鼠亚科两个新种(稻地上新鼢鼠Pliosiphneus daodiensis sp. nov.、铺路上新鼢鼠P. puluensis sp. nov.)进行了简要记述,以期对泥河湾盆地生物地层学框架的建立有所帮助,并从生物进化的角度为泥河湾层的年代学认识提供新证据。
关键词:
Abstract
The Nihewan Basin has been well known for its Cezonoic fossiliferous fluvio-lacustrine deposits and paleolithic sites for almost a century. There have been considerable research efforts devoted to the understanding of the geology, chronology, and stratigraphy of this complex of sedimentary body. The current fundamental problem lies in the chronological aspect of the Nihewan Beds. Arvicolinae and Myospalacinae are two groups of rodents characterized by rapid evolutionary rates and quantifiable evolutionary trends, and hence particularly helpful in Late Cenozoic biostratigraphic correlations. Therefore, we briefly describe three new species of Arvicolinae and two new species of Myospalacinae that mainly came from the selected sections in the Nihewan Basin. Hopefully, they would contribute towards the establishment of the biostratigraphic framework of the Nihewan Basin and provide new evidence on the chronological understanding of the Nihewan Beds from an evolutionary point of view.
Keywords:
PDF (2042KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
郑绍华, 张颖奇, 崔宁. 记泥河湾盆地晚上新世—早更新世?亚科(Arvicolinae)和鼢鼠亚科(Myospalacinae)五新种. 古脊椎动物学报[J], 2019, 57(4): 308-324 DOI:10.19615/j.cnki.1000-3118.190725
ZHENG Shao-Hua, ZHANG Ying-Qi, CUI Ning.
1 Introduction
The Nihewan Basin has been well known for its Late Cenozoic fossiliferous fluvio-lacustrine deposits and paleolithic sites for almost a century. There have been considerable research efforts devoted to the understanding of the geology, chronology, and stratigraphy of this complex of sedimentary body. The fundamental problem that currently hinders our understanding of this important region lies in the chronological aspect of the Nihewan Beds. There are consistently conflicts between the biostratigraphic correlations and magnetostragraphical dating results, and consequently the chronological restriction and subdivision of the lithologic body are still open to further investigation. We have been working towards establishing a biostratigraphic framework of the Nihewan Beds by collecting micromammal fossils from selected sections in the basin for over three decades (Zheng and Cai, 1991; Zhang et al., 2003; Cai et al., 2004, 2007, 2013; Zheng et al., 2006; Li et al., 2008). Five new species of Arvicolinae and Myospalacinae are briefly described here while taking into consideration their importance in the biostratigraphic correlation. The materials of the new species were collected by Zheng and his colleagues from ten localities; eight of which, including all the type localities, belong to the selected sections mentioned above in the Nihewan Basin. These are Hougou (Li et al., 2008), Laowogou (Zhang et al., 2003; Cai et al., 2004), Nangou (Li et al., 2008), Yuanzigou (Li et al., 2008), Donggou (Zheng et al., 2006), Niutoushan (Cai et al., 2007), Jiangjungou (Li et al., 2008), and Danangou (Zheng and Cai, 1991; Cai et al., 2004). Some referred materials of Borsodia prechinensis sp. nov. are from Wenwanggou, Lingtai, Gansu Province (Zhang et al., 2011). One referred partial cranium of Pliosiphneus puluensis sp. nov. is from the Yushe Basin, but further details about its locality and horizon are unknown.The descriptive terminology and measuring methods for the dental morphology of the Arvicolinae are based on previously published literatures (occlusal morphology and measuring methods: Tesakov, 2004; enumeration and measuring methods of isthmuses (Is): Kawamura, 1988; SDQ (Schmelzband Differenzierungs Quotient): Heinrich, 1990; parameter E, Ea, and Eb: van de Weerd, 1976; sinuous line, HH-Index, PAA-Index, and PA-Index: Carls and Rabeder, 1988) and those for the dental and cranial morphology of Myospalacinae are after Liu et al. (2014) and Zheng (1994).
2 Systematic description
Cricetidae Rochebrune, 1883Arvicolinae Gray, 1821
Mimomys Forsyth Major, 1902
Mimomys nihewanensis sp. nov.
(Figs. 1, 2)
Fig. 1
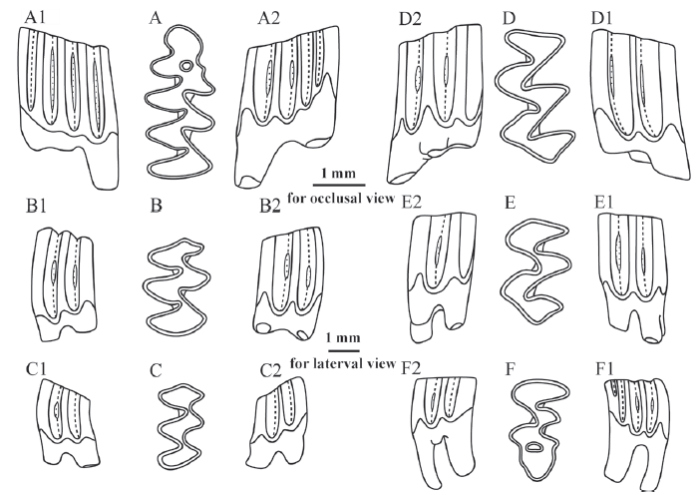 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1Selected molars of Mimomys nihewanensis sp. nov. from Hougou section near Qijiazhuang village, Yangyuan County, Hebei Province
A-A2. right m1 (IVPP V 23157.9, holotype), B-B2. right m2 (V 23157.25), C-C2. right m3 (V 23157.32), D-D2. left M1 (V 23157.38), E-E2. left M2 (V 23157.61), F-F2. left M3 (V 23157.76);A-F. occlusal view, A1-F1. buccal view, A2-F2. lingual view
Fig. 2
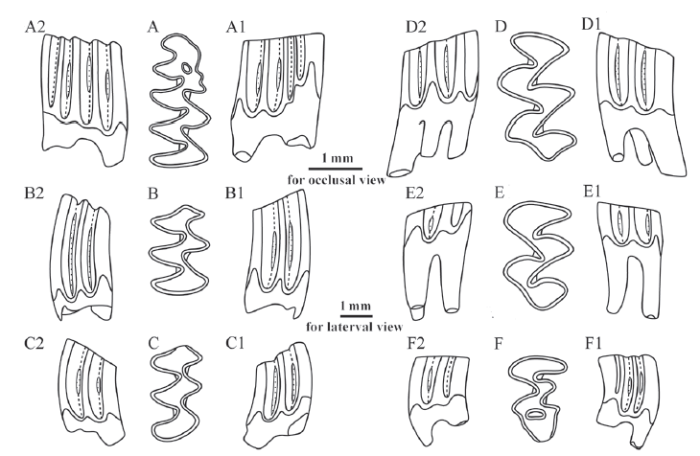 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Selected molars of Mimomys nihewanensis sp. nov. from Laowogou section near Daodi village, Yangyuan County, Hebei Province
A-A2. right m1 (IVPP V 23160.2), B-B2. right m2 (V 23160.16), C-C2. right m3 (V 23160.24),D-D2. left M1 (V 23160.28), E-E2. left M2 (V 23160.36), F-F2. left M3 (V 23160.50); A-F. occlusal view,A1-F1. buccal view, A2-F2. lingual view
Mimomys stehlini, Mimomys orientalis Cai et al., 2004, table 2
Mimomys sp., Mimomys sp. 2 Li et al., 2008, tables 1, 4-6
Mimomys sp. Zheng et al., 2006, table 2; Cai et al., 2007, table 1; Cai et al., 2013, figs. 8.2, 8.4-6
Mimomys sp. 2 Cai et al., 2013, fig. 8.2
Holotype A right m1 (IVPP V 23157.9).
Paratypes A left and a right mandible with m1, six left and ten right m1, six left and six right m2, a left and five right m3, nine left and 15 right M1, six left and nine right M2, five left and six right M3 (V 23157.1-8, 10-87).
Type locality and horizon Layer 4 of Hougou section near Qijiazhuang village, Xinbao Town, Yangyuan County, Hebei Province; Daodi Formation of Upper Pliocene (correlated to the Mazegou Formation in Yushe Basin).
Etymology The name refers to the Nihewan Basin, where the fossil materials representing the new species were collected.
Referred localities and materials Laowogou section near Daodi village, Xinao Town, Yangyuan County, Hebei Province: Layer 3 (~3.08 Ma, Cai et al., 2013), seven right m1, two left and two right m2, three left and four right m3, three left and four right M1, five left and two right M2, four left and six right M3 (V 23155.1-42); Layer 9 (~3.04 Ma, Cai et al., 2013), a left mandible with m1-2, a left mandible with m1-3, six left and four right m1, two left and five right m2, three left and two right m3, a left and four right M1, a left and four right M2, a left and two right M3 (V 23155.43-79); Layer 11 (~2.89 Ma, Cai et al., 2013), a left and eight right m1, six left and three right m2, five left and four right m3, three left and five right M1, eight left and six right M2, seven left and seven right M3 (V 23160.1-63). Nangou section near Hongya village, Xinbao Town, Yangyuan County, Hebei Province: Layer 1, two left and three right m1, three left and a right m2, five left and three right m3, four left and a right M1, three left and eight right M2, six left and three right M3 (V 23162.1-42); Layer 4, a left and a right mandible with m1-2, a right mandible with m1, eight left and six right m1, four left and three right m2, two left and five right m3, seven left and seven right M1, fifteen left and ten right M2, twelve left and ten right M3 (V 23162.43-134); Layer 6, a left and two right m1, a right m2, three left and three right M1, three right M2 (V 23162.135-147). Hougou section near Qijiazhuang village, Xinbao Town, Yangyuan County, Hebei Province: Layer 2, a left m1 (V 23157.98); Layer 5, a right m1, two left m2, a left and a right M1, a right M2, four right M3 (V 23157.88-97). Yuanzigou village, Xinbao Town, Yangyuan County, Hebei Province: Layer 2, a left and a right m1, a right m2, a left and a right M1, a right M2 (V 23158.1-6); Layer 4, a left and two right M1, a left M2 (V 23158.7-10). Donggou section near Qianjiashawa village, Huashaoying Town, Yangyuan County, Hebei Province: Layer 2, three left and nine right m1, five left and seven right m2, four left and eight right m3, six left and seven right M1, four left and nine right M2, ten left and five right M3 (V 23159.3-79); Layer 4 (>2.6 Ma, Cai et al., 2013), a left m3, a right M3 (V 23159.1-2). Niutoushan section near Pulu village, Baishuiquan Town, Yuxian County, Hebei Province: Layer 6, a left and two right m2, two right m3, two left M1, two right M2, two left and a right M3 (V 23161.1-12) and two left and seven right m1, a left and seven right m2, a left and four right m3, three left and four right M1, four left and three right M2, two left and two right M3 (V 23161.13-52, collected by Baoquan Cai in 1980s); Layer 9, a left mandible with m1, a right mandible with m2-3, two left and two right m1, a right m2, two right M1, three left and three right M2 (V 23156.1-15) and four left and three right m1, a left and four right m2, three left and two right m3, four left and two right M1, six right M2, three right M3 (V 23156.16-47, collected by Baoquan Cai in 1980s). Jiangjungou section, Beishuiquan Town, Yuxian County, Hebei Province: Layer 1, a left mandible with m2, a right m1, two left and three right m2, a right m3, two left and a right M1, three left M2, a left and two right M3 (V 23153.1-17).
Age of referred localities Late Pliocene (~3.08-2.89 Ma, Cai et al., 2013).
Measurements See Tables 1-3.
Table 1
Table 1Average width and rate of closedness of isthmuses and SDQ on m1 of related arvicoline species1) from China
| Species | average width (mm)|rate of closedness (%)2) | min.|mean|max., n | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| n | Is 1 | Is 2 | Is 3 | Is 4 | Is 5 | Is 6 | Is 7 | SDQ | ||
| Borsodia chinensis | 26 | 0.04|100 | 0.05|81.0 | 0.04|100 | 0.05|100 | 0.21|0 | 0.22|0 | 0.41|0 | 53|67|85, 21 | |
| B. prechinensis sp. nov. | 1 | 0.07|100 | 0.08|100 | 0.07|100 | 0.07|100 | 0.17|0 | 0.29|0 | na | 91|91|91, 1 | |
| B. mengensis | 5 | 0.05|100 | 0.10|80.0 | 0.07|100 | 0.07|80.0 | 0.25|0 | 0.31|0 | 0.40|0 | 101|115|122, 5 | |
| Mimomys nihewanensis sp. nov. | 58|55 | 0.06|92.5 | 0.15|0 | 0.09|61.7 | 0.08|63.6 | 0.36|0 | 0.47|0 | na | 112|138|168, 43 | |
| M. orientalis | 41|19 | 0.06|100 | 0.13|26.3 | 0.10|47.4 | 0.07|94.7 | 0.30|0 | 0.41|5.3 | 0.45|0 | 113|132|157, 21 | |
| Pitymys simplicidens sp. nov. | 1 | 0.03|100 | 0.03|100 | 0.03|100 | 0.03|100 | 0.16|0 | 0.08|100 | 0.22|0 | 62|62|62, 1 | |
新窗口打开|下载CSV
Table 2
Table 2E, Ea, Eb, HH-Index, PAA-Index, PA-Index, and length of molars of related Mimomys, Borsodia and Pitymys species* of China (min.|mean|max.,n) (mm)
| Species | m1 | m2 | m3 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| L | E | Ea | Eb | HH-Index | L | HH-Index | L | HH-Index | |||
| M. banchiaonicus | 3.90|3.90|3.90, 1 | na|>3.76|na, 1 | |||||||||
| M. bilikeensis | 2.26|2.46|2.66, 30 | 0.32|0.70|1.00, 31 | 0.24|0.50|0.76, 31 | 0.08|0.19|0.36, 31 | -0.08|0.10|0.25, 31 | 1.60|1.71|1.84, 16 | -0.04|0.10|0.22, 16 | 1.28|1.44|1.60, 10 | -0.09|-0.16|-0.20, 10 | ||
| M. gansunicus | 2.59|2.74|2.92, 15 | na|>3.33|na, 1 | na|>3.20|na, 1 | na>3.17|na,1 | na|>5.18|na, 1 | ||||||
| M. orientalis (Gaotege) | 2.44|2.87|3.20, 14 | 2.00|2.52|2.84, 5 | 1.84|2.45|2.72, 5 | 0.96|1.27|1.48, 14 | 0.97|1.27|1.52, 14 | 1.60|1.73|1.92, 12 | 0.75|0.97|1.32, 12 | 1.44|1.53|1.56, 7 | 0.32|0.43|0.54, 7 | ||
| M. orientalis (Danangou) | 2.60|2.73|2.88, 26 | 1.00|2.05|2.48, 15 | 1.40|1.87|2.28, 15 | 0.80|1.01|1.24, 15 | 0.78|1.16|1.63, 26 | 1.68|1.78|1.88, 17 | 0.72|0.94|1.40, 17 | 1.40|1.55|1.72, 12 | 0.12|0.37|0.57, 12 | ||
| M. orientalis (combined) | 2.44|2.78|3.20, 40 | 1.00|2.17|2.84, 20 | 1.40|2.02|2.72, 20 | 0.80|1.14|1.48, 29 | 0.78|1.20|1.63, 40 | 1.60|1.76|1.92, 29 | 0.72|0.95|1.40, 29 | 1.40|1.54|1.72, 19 | 0.12|0.39|0.57, 19 | ||
| M. nihewanensis sp. nov. | 2.44|2.80|3.12, 51 | 1.32|1.81|2.40, 23 | 1.16|1.63|2.20, 23 | 0.08|0.78|1.40, 54 | 0.56|0.81|1.16, 51 | 1.52|1.82|2.12, 61 | 0.14|0.67|1.22, 61 | 1.24|1.58|1.76, 38 | 0.07|0.27|0.57, 38 | ||
| M. peii | 3.47|3.64|4.01, 7 | na|>5.30|na,1 | 4.0|4.32|4.63, 2 | 2.25|2.25|2.25, 2 | na|>6.51|na, 2 | ||||||
| M. teilhardi | 2.17|2.55|2.85, 102 | 0.40|0.99|1.64, 60 | 0.28|0.92|1.64, 60 | 0.14|0.49|1.12, 60 | 0.20|0.46|1.00, 62 | 1.52|1.71|1.88, 62 | 0.20|0.40|0.65, 62 | 1.28|1.43|1.56, 39 | 0.00|0.12|0.31, 39 | ||
| M. youhenicus | 2.61|2.81|3.00, 2 | 3.30|3.30|3.30, 1 | 2.95|2.95|2.95, 1 | 2.17|2.35|2.53, 2 | 3.06|3.25|3.44, 2 | ||||||
| P. simplicidens sp. nov. | 2.36|2.36|2.36, 1 | ||||||||||
| B. mengensis | 2.20|2.38|2.60, 5 | 0.50|1.18|1.61, 4 | 1.50|1.60|1.73, 4 | 0.58|0.77|0.89, 4 | 1.23|1.27|1.30, 2 | 0.26|0.40|0.53, 2 | |||||
| B. chinensis | 2.32|2.54|2.80, 21 | >4.74|>5.35|>6.63, 12 | 1.50|1.51|1.73, 8 | >1.84|>4.15|>4.83, 8 | 1.37|1.47|1.57, 17 | >2.11|>3.08|>3.84, 17 | |||||
| B. prechinensis sp. nov. | 2.63|2.63|2.63, 1 | na|≥3.67|na, 1 | 1.60|1.60|1.60, 1 | na|>3.62|na, 1 | 1.84|1.93|2.01, 2 | ||||||
| Species | M1 | M2 | M3 | ||||||||
| L | PAA-Index | PA-Index | L | PA-Index | L | PA-Index | |||||
| M. orientalis (Gaotege) | 2.00|2.38|2.68, 24 | 0.79|1.45|2.09, 24 | 0.78|1.38|2.04, 24 | 1.52|1.87|2.12, 19 | 0.36|0.73|1.10, 19 | 1.64|1.82|2.08, 9 | 0.23|0.36|0.63, 9 | ||||
| M. orientalis (Danangou) | 2.24|2.45|2.88, 40 | 1.09|1.51|1.84, 40 | 1.07|1.43|1.78, 40 | 1.72|1.84|2.04, 30 | 0.54|0.86|1.17, 30 | 1.56|1.69|1.88, 9 | 0.26|0.50|0.71, 9 | ||||
| M. orientalis (combined) | 2.00|2.42|2.88, 64 | 0.79|1.49|2.09, 64 | 0.78|1.41|2.04, 64 | 1.52|1.85|2.12, 49 | 0.36|0.81|1.17, 49 | 1.56|1.76|2.08, 18 | 0.23|0.43|0.71, 18 | ||||
| M. nihewanensis sp. nov. | 1.88|2.45|2.88, 69 | 0.60|1.20|1.84, 69 | 0.72|1.17|1.80, 69 | 1.56|1.90|2.08, 91 | 0.24|0.65|1.20, 91 | 1.52|1.81|2.12, 76 | 0.11|0.39|0.71, 79 | ||||
| B. mengensis | 1.90|2.04|2.07, 4 | 0.69|0.90|1.15, 4 | 0.61|0.67|0.89, 4 | 1.73|1.81|1.90, 3 | 0.57|0.67|0.77, 2 | 1.37|1.51|1.83, 8 | 0.37|0.55|0.81, 8 | ||||
| B. chinensis | 1.98|2.16|2.37, 20 | >3.23|>5.04|>5.66, 20 | >3.85|>6.05|>6.83, 20 | 1.63|1.79|1.87, 16 | >2.69|>4.90|>6.25, 16 | 1.30|1.46|1.67, 8 | >2.12|>3.75|>4.79, 8 | ||||
| B. prechinensis sp. nov. | 2.03|2.10|2.17, 2 | 5.32|5.42|5.52, 2 | 4.57|4.62|4.67, 2 | 1.90|1.90|1.90, 1 | 1.67|1.67|1.67, 1 | 3.03|3.03|3.03, 1 | |||||
新窗口打开|下载CSV
Table 3
Table 3Proportions of 2- and 3-rooted M2 and M3 in four Mimomys species from China
| Species | Locality | M2 (n|%) | M3 (n|%) | |||
|---|---|---|---|---|---|---|
| 3-rooted | 2-rooted | 3-rooted | 2-rooted | |||
| M. bilikeensis | Bilike | 15|100 | 0|0 | 11|68.8 | 5|31.3 | |
| M. teilhardi | Gaotege | 64|100 | 0|0 | 2|5.9 | 32|94.1 | |
| M. orientalis | Gaotege | 12|75 | 4|25 | 1|14.3 | 6|85.7 | |
| Danangou | 16|61.5 | 10|38.5 | 5|83.3 | 1|16.7 | ||
| combined | 28|66.7 | 14|33.3 | 6|46.2 | 7|53.9 | ||
| M. nihewanensis sp. nov. | Laowogou etc | 75|96.2 | 3|3.9 | 64|84.2 | 12|15.8 | |
新窗口打开|下载CSV
Diagnosis In terms of average length of m1, Mimomys nihewanensis sp. nov. (2.80 mm) is similar in size to M. orientalis (2.78 mm), but its Is 1-4 are less closed (completely closed on 92.5%, 0%, 61.7%, and 63.6% of specimens, respectively) than the latter (completely closed on 100%, 26.3%, 47.4%, and 94.7% of specimens, respectively) (Table 1); SDQp is 138, greater than that of the latter (132) (Table 1); the average HH-index is 0.81, less than that of the latter (1.20); means of parameter E, Ea and Eb are 1.81, 1.63, and 0.78, respectively, which are also smaller than those of the latter (2.17, 2.02, 1.14, respectively) (Table 2); the percentage of 3-rooted M2s is 96.2%, higher than the latter (66.7%); the percentage of 3-rooted M3 (84.2%) is also higher than the latter (46.2%) (Table 3); means of PAA-Index and PA-Index are 1.20 and 1.17, respectively, smaller than those of M. orientalis (1.49 and 1.41, respectively) (Table 2). Little cementum is deposited in re-entrant angles, differing from M. bilikeensis and M. teilhardi.
Description Medium-sized. Molars relatively low crowned, with little cementum deposited in re-entrant angles.
The average length of m1 is 2.80 mm (Table 2). Anterior to PL, there exist three alternating triangles and a complicated ACC composed of AC2, PF, MR, IF, and EI. They, as a whole, form five LSAs, four LRAs, four BSAs and three BRAs altogether (Figs. 1A-A2, 2A-A2). An oval-shaped EI is usually formed during the early wear of the tooth and lasts for a long time. MR and PF are distinct and extend to the base of the crown. Is 1, Is 3 and Is 4 are closed on 92.5%, 61.7% and 63.6% of all the examined m1s, respectively (Table 1). SDQp is 138 (Table 1). The average HH-Index is 0.81, and the average parameter E, Ea, and Eb are 1.81, 1.63, and 0.78, respectively (Table 2).
The average length of m2 is 1.82 mm. Anterior to PL, there exist two pairs of slightly alternating triangles. Is 1 and Is 3 are generally closed, whereas Is 2 and Is 4 are always open (Figs. 1B-B2, 2B-B2). The average HH-Index is 0.67 (Table 2).
The average length of m3 is 1.58 mm. The morphology of its occlusal surface is generally similar to that of m2, but markedly shrinks in length and width (Figs. 1C-C2, 2C-C2). The average HH-Index is 0.27 (Table 2).
The average length of M1 is 2.45 mm. Posterior to AL, there exist four alternating triangles. It has three roots, with the third root situated lingually between the anterior and posterior root (Figs. 1D-D2, 2D-D2). The average PAA-Index is 1.20, and the average PA-Index is 1.17 (Table 2).
The average length of M2 is 1.90 mm. The morphology of the occlusal surface is generally similar to the part of M1 posterior to AL (Figs. 1E-E2, 2E-E2). 96.2% (75 out of 78 specimens) of all the examined M2s have 3 roots. The average PA-Index is 0.65 (Table 2).
The average length of M3 is 1.81 mm. The morphology of its occlusal surface is more complicated than that of M2, with three salient angles and two re-entrant angles on each side (Figs. 1F-F2, 2F-F2). The lingual posterior re-entrant angle (LRA3) is very deep, and a circular EI is usually formed after wear. 84.2% of M3s (64 out of 76 specimens) have three roots, and 15.8% of M3s have two roots. The average PA-Index is 0.39 (Table 2).
Comparisons and discussion In terms of m1 average length, Mimomys nihewanensis sp. nov. (2.80 mm) is strikingly smaller than M. banchiaonicus (3.90 mm), and M. peii (3.64 mm), but larger than M. bilikeensis (2.46 mm), and M. teilhardi (2.55 mm), while most similar to M. orientalis (Gaotege: 2.87 mm; Danangou: 2.73 mm) and M. youhenicus (2.81 mm) (Table 2). The average values of E, Ea, and Eb of m1 of M. nihewanensis (1.81, 1.63, and 0.78, respectively) are greater than those of M. bilikeensis (0.70, 0.50, 0.19), and M. teilhardi (0.99, 0.92, 0.49), but smaller than those of M. youhenicus (3.30, 2.95, 2.35), M. gansunicus (>3.33, >3.20, >3.17), and M. orientalis (Gaotege: 2.52, 2.45, 1.27; Danangou: 2.05, 1.87, 1.01). The average m1 HH-Index of the new species (0.81) lies between M. teilhardi (0.46) and M. orientalis (Gaotege: 1.27; Danangou: 1.16). The proportion of 3-rooted M2 for M. nihewanensis is 96.2%, closer to that of M. orientalis from Gaotege (75.0%), much higher than that of M. orientalis from Danangou (61.5%). The proportion of 3-rooted M3 for the new species is 84.2%, similar to that of M. orientalis from Danangou (83.3%), much higher than that of M. orientalis from Gaotege (14.3%) and that of M. teilhardi from Gaotege (5.9%) (Table 3). The reason that M. teilhardi has a very low proportion of 3-rooted M3 is probably because of the misidentification of quite a number of M3s that may actually belong to Germanomys.
Mimomys nihewanensis is smaller in size than M. stehlini Kormos, 1931 from Europe (length of m1: 3.07 mm), and more primitive than the latter because of smaller parameters of enamel free area on buccal side of m1, but more derived than the latter because of the cementum deposited in re-entrant angles of its molars. The contradiction probably results from the lack of a form at the similar evolutionary stage to M. nihewanensis in Europe.
All the materials referred to the new species indicate that little cementum is deposited in the re-entrant angles of the molars, which is consistent with the occurrence of Mimomys orientalis in the overlying strata of the same basin, so M. nihewanensis is most likely directly ancestral to M. orientalis. Most of the Gaotege specimens referred to M. orientalis don’t have cementum (Qiu and Li, 2016), which seems inconsistent with the general evolutionary trend of increasing deposition of cementum from none to abundant along different lineages. This phenomenon may be explained by the different ecology of the two areas: the altitude and latitude of the Nihewan Basin are relatively low, so the vegetation is better developed because of the higher temperatures and humidity, which also caused better development of cementum in M. orientalis; in contrast, the altitude and latitude of Gaotege are relatively high, so the vegetation is less developed because of the lower temperatures and humidity, which caused less developed cementum in M. orientalis.
All the other localities of Mimomys nihewanensis are probably older than Laowogou (3.08 Ma) but should be no older than the horizon of Gaotege yielding M. teilhardi (4.2 Ma).
Pitymys McMurtrie, 1831
Pitymys simplicidens sp. nov.
(Fig. 3)
Fig. 3
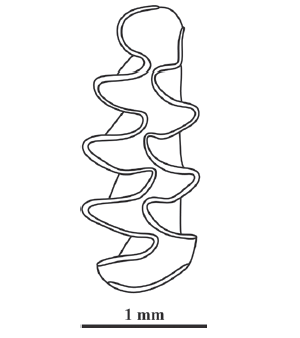 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3Pitymys simplicidens sp. nov. (right m1, GMC V 2056.1, holotype) from Danangou section near Dongyaozitou village, Yuxian County, Hebei Province (After Zheng and Cai, 1991)
Pitymys cf. hintoni Zheng and Cai, 1991, figs. 4, 4-5; Cai et al., 2004, table 2
Pitymys hintoniMin et al., 2006, p. 104
Holotype A right mandible with i1 and m1 (Geological Museum of China, GMC V 2056.1).
Paratype A right m1 lacking PL (GMC V 2056.2).
Type locality and horizon Layer 13 (Equivalent to Do-6 of Zheng and Cai, 1991) of Danangou section near Dongyaozitou village, Baishuiquan Town, Yuxian County, Hebei Province, Lower Pleistocene (~1.44 Ma, Cai et al., 2013).
Etymology Derived from Latin words simplex and dens, meaning “simple” and “tooth”, respectively, to indicate the simplified morphology of AC2 on m1 of the new species, without trace of additional RA (or SA).
Referred localities and materials Danangou section near Dongyaozitou village, Baishuiquan Town, Yuxian County, Hebei Province: Layer 12 (~1.49 Ma, Cai et al., 2013), a left m1 (IVPP V 23224).
Age of referred localities Early Pleistocene (~1.49 Ma, Cai et al., 2013).
Measurements See Tables 1-2.
Diagnosis Small-sized (length of m1: 2.36 mm); AC2 of m1 short and simple, without trace of LRA5 and BRA4; Is 1-4 and Is 6 are closed, whereas Is 5 and Is7 are open; the value of SDQI is 62, so the differentiation of enamel is negative type.
Description T1-T3 of m1 are separated from each other, while T4 and T5 are confluent with each other, and form a rhombus; AC2 is simple and oval-shaped, without trace of extra RA or SA; Is 1-4 and Is 6 are closed, whereas Is 5 and Is 7 are not; the value of SDQI is 62, indicative of negative differentiation; m1 has five LSAs and four BSAs, and four LRAs and three BRAs; cementum is abundant in all re-entrant angles.
Comparisons The rhombus-forming confluent T4 and T5 and completely closed Is 6 are consistent with the dental morphology of Pitymys. P. hintoni Kretzoi, 1941 from the locality Betfia-5 of Biharian age, Romania also has rhombus-forming confluent T4 and T5, and closed Is 6, but LRA5 is distinctly developed on the lingual side of AC2. P. gregaloides Hinton, 1923 from the Upper Freshwater Bed of West Runton, England also has a faint BRA4 developed besides LRA5, so AC2 is elongated. The extant Blyth’s mountain vole P. leucurus (Blyth, 1863), Sikkim mountain vole P. sikimensis (Hodgson, 1849), Irene’s mountain vole P. irene (Thomas, 1911), and Juniper mountain vole P. juldaschi (Severzov, 1879) are radically different from the new species by having common or similar characters on m1, that BRA3 is broader and shallower than LRA5, and relatively posteriorly located, and that Is 6 is not closed. In addition, the new species is clearly smaller than the extant species. Based on the measurements of m1-3 length of Luo et al. (2000), except for P. irene, the m1s of all the other extant species mentioned above are estimated to be equal to or greater than 3 mm.
Discussion With the exception of the fact that the dentine area between AC2 and T5 is closed, the occlusal morphology of Pitymys simplicidens sp. nov. is remarkably similar to Allophaiomys deucalion, which may prove that the new species is the direct descendant of the latter. However, P. simplicidens. is unlikely the ancestor of the extant species of Pitymys, because the closed Is 6 on the m1 of the new species indicates that it lies at the end of an evolutionary lineage, and it has not been found anywhere else other than in the Nihewan Basin.
The “Pitymys hintoni” reported by Min et al. (2006) from Taiergou west section near Xiaodukou village, Huashaoying Town, Yangyuan County reported by Min et al. (2006) should be a synonym of the new species, so it could be taken as evidence that the species was widely distributed in the Nihewan Basin.
Borsodia Jánossy & van der Meulen, 1975
Borsodia prechinensis sp. nov.
(Fig. 4)
Fig. 4
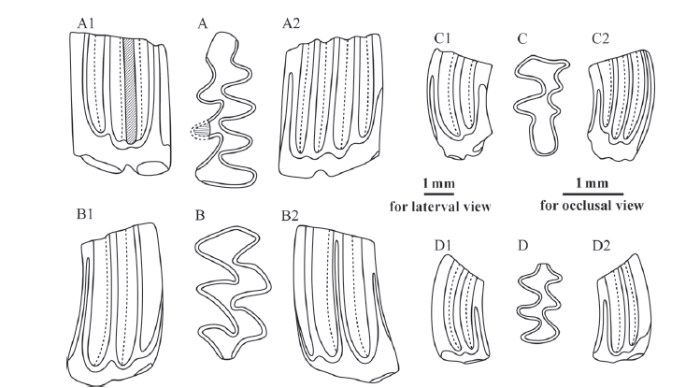 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 4Selected molars of Borsodia prechinensis sp. nov. from Donggou section near Qianjiashawa village, Yangyuan County, Hebei Province
A-A2. right m1 (IVPP V 23164.1, holotype), B-B2. right M1 (V 23164.4), C-C2. left M3 (V 23164.6),D-D2. right m3 (V 23164.3); A-D. occlusal view, A1-D1. buccal view, A2-D2. lingual view
Borsodia sp., B. chinensis (partim) Zheng et al., 2006, table 2
Borsodia n. sp. Zheng and Zhang, 2000, fig. 2; Zheng and Zhang, 2001, fig. 3
Borsodia sp. Zhang et al., 2011, fig. 3A-F
Arvicolinae gen. et sp. indet. Zhang et al., 2011, fig. 3K-L
Holotype A left m1 (IVPP V 23164.1).
Paratypes A left m1, a right m3, two right M1, a right M3 (V 23164.2-6).
Type locality and horizon Layer 11 of Donggou section near Qianjiashawa village, Huashaoying Town, Yangyuan County, Hebei Province, Lower Pleistocene (<2.6 Ma, Cai et al., 2013).
Etymology pre-, Latin, meaning "before", to indicate the new species chronologically predates Borsodia chinensis.
Referred locality and materials Donggou section near Qianjiashawa village, Huashaoying Town, Yangyuan County, Hebei Province: Layer 4 (>2.6 Ma, Cai et al., 2013), a right m2 (IVPP V 23147.1); Layer 7 (>2.6 Ma), a right m1, a left m3, two right M1, a right M2 (V 23147.2-6). Niutoushan section near Pulu village, Beishuiquan Town, Yuxian County, Hebei Province: Layer 15, a left and three right m1, a left and a right m2, two left M1, three left and a right M2 (V 23147.39-50). Wenwanggou section near Leijiahe village, Lingtai County, Gansu Province: Layer WL11-7 (~3.5-3.41 Ma, Zheng and Zhang, 2000), a right M3 (V 18079.1); Layer WL11-5, a left m1 (V 18081.1); Layer WL11-3 (~3.45 Ma, Zheng and Zhang, 2000), a right M3 (V 18079.2); Layer WL11-1, a right m3 (V 18079.3); Layer WL10-11, a broken left M3 (V 18079.4); Layer WL10-10 (~3.33 Ma, Zheng and Zhang, 2000), a left M3 (V 18079.5); Layer WL10-8, a broken right M3 (V 18079.6); Layer WL10 (~3.41-3.25 Ma, Zheng and Zhang, 2000), two left M3, two right m2, a broken left m3 (V 18079.7-11); Layer WL 8 (~3.15 Ma, Zheng and Zhang, 2000), a right M3, a broken right m1, a broken right m3 (V 18079.12-14), a left m1 (V 18081.2); Layer WL 3 (~2.16 Ma, Zheng and Zhang, 2000), a broken right m1, a broken left m2 (V 18079.15-16).
Age of referred localities Late Pliocene-Early Pleistocene (~3.5-2.2 Ma, Cai et al., 2013; Zheng and Zhang, 2000).
Measurements See Tables 1-2.
Diagnosis Similar to Borsodia chinensis in size, but LRA2 on upper molars is narrow and not Lagurus-type. AC of m1 is short, and bends buccally to a lesser extent, and MR is developed on some specimens. HH-index of m1 (≥3.67) and PAA-index (5.42) and PA-index (4.62) of M1 are distinctly smaller than B. chinensis (>5.35, >5.04, and >6.05, respectively), but greater than B. mengensis (1.18, 0.90, and 0.67, respectively) (Table 2). Is 1-4 of m1 are 100% closed, the same as B. chinensis, but higher than B. mengensis; SDQI (91) indicative of the same negative differentiation as B. chinensis. Is 1 and Is 3 of M1 (average length: 2.10 mm) are closed, and Is 2 and Is 4 are not, which is similar to B. mengensis, but different from B. chinensis with closed Is 1-4.
Description Root development of molars occurs relatively early, and no cementum deposited in the re-entrant angles.
On m1, there exist three closed and alternating triangles and 1 three-lobed ACC anterior to PL. Is 1-4 are closed, while Is 5-6 are open. There are five LSAs and four BSAs, and four LRAs and three BRAs. On some specimens, the vestige of MR exists on the anterior wall of BSA3. AC is relatively short, and does not bend buccally, and the front of which lacks enamel. The posterior edge of the tooth is flat or slightly concave anteriorly. HH-Index≥3.67 (Hsd = 2.27, Hsld≥2.87).
On m2, four triangles exist anterior to PL. Is 1 and Is 3 are closed, whereas Is 2 and Is 4 are open, resulting in two rhombic transverse dentine zones. HH-Index > 3.62 (Hsd > 2.3, Hsld > 2.80).
The m3 is similar to m2 in occlusal morphology, but shorter and narrower. HH-Index averaged 1.93 (Hsd = 1.50, Hsld = 1.33).
M1 (average length: 2.10 mm) has four triangles posterior to AL. Is 1 and Is 3 are more closed than Is 2 and Is 4. 3-rooted. PAA-index = 5.42 (Prs = 3.40, As = 3.13, Asl = 2.83), PA-Index = 4.62.
M2 has three triangles posterior to AL, and 2 LSAs, 1 LRA and 3 BSAs, 2 BRAs. Prs is greater than 2.2. 2-rooted.
On M3, the three BRAs are shallower than the two LRAs, and the three BSAs are weaker than the two LSAs. PL is relatively broad and long. PA-Index = 3.03 (Prs = 2.00, As = 2.27). 2-rooted.
Comparisons The HH-Index for the lower molars and the PA-Index for the upper molars of Borsodia prechinensis sp. nov. are indicative of its intermediate stage of evolution between B. mengensis and B. chinensis, but closer to the latter (Table 2), which means the new species is much more derived than B. mengensis, but slightly more primitive than B. chinensis.
The morphology of molars of the new species is different from B. chinensis in: AC of m1 does not bend buccally, and retains a vestigial MR on some specimens; re-entrant angles and salient angles on both buccal and lingual side are more pronounced; LRA2 on M1 and M2 is narrow and V-shaped, but not broad and U-shaped.
Discussion The two m1s from Layer WL11-5 of Wenwanggou section, Lingtai County, Gansu Province were all referred to as Arvicolinae gen. et sp. indet. (Zhang et al., 2011:fig. 3K-L), but both of them were thought to be with little cementum in the re-entrant angles. However, except for the existence of cementum, their occlusal morphology is consistent with that of the new species in such characters as the AC not bending buccally, vestigial Mimomys-ridge, etc. So, the existence of cementum is most likely not true. Here we refer all the specimens of Borsodia sp. (Zhang et al., 2011:fig. 3A-F) and Arvicolinae gen. et sp. indet. of the same authors to the new species.
Based on the distribution of small mammals in the Donggou section (Zheng et al., 2006) and earlier paleomagnetic dating data (Yang et al., 1996; Yuan et al., 1996; Zhu et al., 2001), the boundary between Pliocene and Pleistocene was determined to be between L8 and L9 in the Donggou section, which is indicative of the diachronism of Borsodia prechinensis sp. nov. Similarly, in the Wengwanggou section, the boundary between Pliocene and Pleistocene was determined between WL7 and WL6, where the new species is also diachronous (Zheng and Zhang, 2000, 2001).
Myospalacinae Lilljeborg, 1866
Pliosiphneus Zheng, 1994
Pliosiphneus daodiensis sp. nov.
(Fig. 5)
Fig. 5
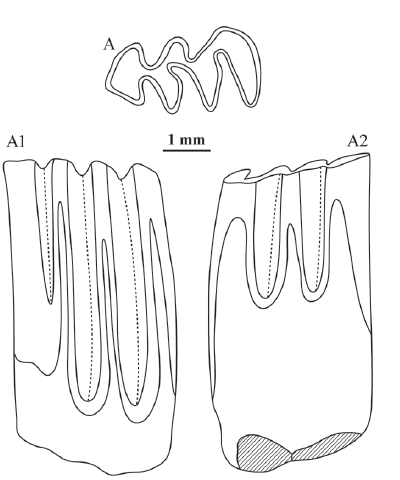 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 5Right m1 (IVPP V 15475, holotype) of Pliosiphneus daodiensis sp. nov. from Laowogou section near Daodi village, Yangyuan County, Hebei Province
A. occlusal view, A1. lingual view, A2. buccal view
Pliosiphneus sp. Cai et al., 2004, table 2
Holotype A right m1 of an adult individual (IVPP V 15475).
Type locality and horizon Layer 3 (red clay) of Laowogou section near Daodi village, Yangyuan County, Heibei Province (Upper Pliocene, ~3.08 Ma, Cai et al., 2013).
Etymology The species name refers to Daodi village, in the vicinity of which is the type locality.
Measurements Length of m1: 3.34 mm; parameter a, b, c, d, and e: 0.0, 3.8, 3.7, 4.2, and 3.7, respectively.
Diagnosis Distinctly smaller than Pliosiphneus lyratus, and crown height is lower than P. puluensis sp. nov. Length of m1 is 3.34 mm, and parameter a, b, c, d, and e are 0, 3.8, 3.7, 4.2, and 3.7, respectively.
Description Molars rooted. M1 has a pointed anterior tip, and a triangular ac; there exist three alternating, and tightly closed triangles; bras are distinctly shallower than lras; bsa2 is weaker than bsa1 and bsa3; the depth successively decreases from lra1 to lra3; bra1 and bra2 are of similar depth; bra2 and lra3 are transversely opposite to each other. Dentine tracts on both buccal and lingual side are three-peaked type, but the lingual ones are much higher. The value of parameter a is 0.
Comparisons The m1 of Pliosiphneus daodiensis sp. nov. (length: 3.34 mm) is slightly bigger than that of P. antiquus (averange length: 3.22 mm, Yushe Basin, Zheng, 2017:table 9.7), distinctly smaller than P. lyratus (average length: 4.35 mm, Yushe Basin, Zheng, 2017, remeasured) and P. puluensis sp. nov. (length: 4.5 mm). Values of parameter a, b, c, d, and e of the new species (0.0, 3.8, 3.7, 4.2 and 3.7, respectively) are greater than that of P. antiquus (0.07, 0.73, 1.07, 1.55 and 0.60, respectively, Yushe Basin, Zheng, 2017) and P. lyratus (0, 1.9, 1.7, >4.6 and >3.6, respectively, Yushe Basin, Zheng, 2017), but distinctly smaller than P. puluensis (2.4, >5.0, >6.0, >5.2, and >5.1, respectively). If they are lined up in order of crown height from lowest to highest, it should be P. antiquus→P. lyratus→P. daodiensis→P. puluensis, which also represents their successive order of evolution.
Remarks The new species should belong to convex occiput zokors based on its opposite lra3 and bra2. The 0 value of parameter a is indicative of either Prosiphneus or Pliosiphneus assignment. Further, the greater values of parameter b, c, d, and e indicate a derived form of Pliosiphneus.
The pointed anterior tip of ac of m1 is probably because of its young age. The roots of the tooth have not completely erupted, and the crown has not been subject to heavy wear.
Pliosiphneus puluensis sp. nov.
(Fig. 6)
Fig. 6
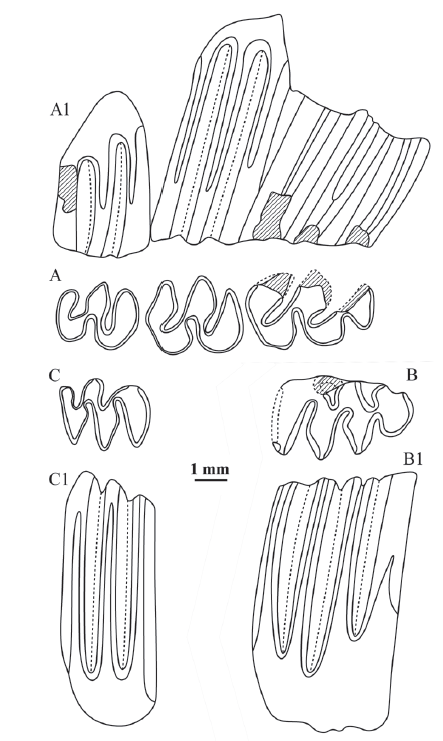 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 6Selected molars of Pliosiphneus puluensis sp. nov. from Yushe Basin, Shanxi Province (A) and Niutoushan section near Pulu village, Yuxian County, Hebei Province (B-C)
A-A1. right M1-3 (IVPP RV 42025),B-B1. left m1 (V 15476, holotype),C-C1. right m2 (V 15476.2); A-C. occlusal view, A1. buccal view, B1-C1. lingual view
Pliosiphneus sp. nov. Cai et al., 2007, tables 1-2
Holotype A left m1 of an adult, with the posterior edge damged (IVPP V 15476).
Paratypes A left and a right m2, anterior part of a right m3, anterior part of a left M1, a right M3 (V 15476.1-5).
Type locality and horizon Layer 9 of Niutoushan section near Pulu village, Beishuiquan Town, Yuxian County, Hebei Province (Daodi Formation of Upper Pliocene, correlated to Mazegou Formation in Yushe Basin).
Etymology The name refers to Pulu village, in the vicinity of which is the type locality.
Referred localitiy and material Yushe Basin, Shanxi Province: anterior part of a cranium with left and right M1-3 (IVPP RV 42025).
Age of referred localities Late Pliocene.
Measurements Length of diastema: 13.0 mm; length of incisive foramen: 6.2 mm; rostrum breadth anterior to infraorbital foramen: 10.5 mm; breadth of interorbital constriction: 9.0 mm; length of M1-3: 9.6 mm; length of M1: 3.9 mm; length / breadth of M2: 2.7 mm / 2.9 mm; length of M3: 2.5 mm; breadth of palate between the medial margins of M1s: 5.1 mm; breadth of palate between the medial margins of M3s: 5.6 mm; length of m1: 4.5 mm; parameter a, b, c, d, and e: 2.40, >5.0, >6.0, >5.2, and >5.1, respectively.
Diagnosis Large-sized. Extremely hypsodont in Pliosiphneus. Rostrum short and broad; frontal area broad; incisive foramen long. Ratio of incisive foramen length to diastema length is 47.7%. The upper molar rows are almost parallel (the ratio of breadth between M1s to that between M3s is 91.1 %). In the molar row, the ratio of length of M3 to length of M2 is 92.6%; the ratio of length of M3 to length of M1-3 is 26.0%. The ratio of breadth to length of M2 is 79.3%. The values of parameter a, b, c, d, and e of m1 (length: 4.5 mm) are 2.40, >5.0, >6.0, >5.2, and >5.1, respectively.
Description On the cranium from Yushe Basin (RV 42025), the heights of lingual dentine tracts of M1 are compatible with that of the Pulu m1 (Fig. 6). The premaxillary-maxillary suture runs across the incisive foramen (length: 6.2 mm) at the posterior third, which indicates the convex occiput type assignment. According to measurements, the cranium possesses Pliosiphneus characters such as a short and broad rostrum, broad frontal area, broad and concave sagittal area. At the same time, opposite bra2 and lra3 on m1 is also a character of convex occiput zokors. These morphological characteristics, combined together, justify the referral of the specimens to a new species of Pliosiphneus.
The ac of m1 (length: 4.5 mm) is oval-shaped. bsas are weaker than lsas, and bras are shallower than lras. The value of parameter a is close to half of the crown height; the dentine tracts of b, c, d, and e have penetrated the crown. On m2 (length: 2.8 mm), the dentine tracts of b and c have not penetrated the crown, but have been very close to the occlusal surface.
Comparisons Pliosiphneus puluensis sp. nov. is the largest known species in the genus. Its length of M1-3 (9.6 mm) is greater than P. antiquus (8.6 mm, Yushe Basin, Zheng, 2017:table 9.7) and P. lyratus (9.35 mm, Yushe Basin, calculated from Zheng, 2017:table 9.5). The length of M1 of the P. puluensis (3.9 mm) is also greater than P. antiquus (2.82 mm, Yushe Basin, Zheng, 2017:table 9.7) and P. lyratus (3.69 mm, Yushe Basin, calculated from Zheng, 2017:table 9.5). The m1 length of the Pulu new species (4.5 mm) is also greater than that of the latter two, 3.22 mm (Yushe Basin, Zheng, 2017:table 9.7) and 4.35 mm (remeasured on specimens from Yushe Basin described by Zheng, 2017), respectively, and greater than that of P. daodiensis (3.34 mm) as well. The Pulu new species is also the most hypsodont form in the genus, as indicated by the values of parameter a, b, c, d, and e of lower molars, and A, B, C, and D of upper molars. The most distinguishable difference is the value of m1 parameter a of the Pulu new species (2.4) is significantly greater than that of P. antiauus (0.07, Yushe Basin, Zheng, 2017:99), P. lyratus (0, Yushe Basin, Zheng, 2017:96), and P. daodiensis sp. nov. (0).
Discussion Judging from the fact that the value of m1 parameter a is about half of the crown height, the Pulu new species most likely represents a new genus with a convex occiput and hypsodont rooted molars. However, it is tentatively referred to Pliosiphneus, taking into consideration that no complete skulls have been found up to now. It is reasonably predictable that this species may have lost its interparietal bone as in the derived concave occiput Mesosiphneus with rooted molars.
The Pulu new species is characterized by a short and broad rostrum, broad frontal area, concave interorbital area and sagittal area, long incisive foramen, and nearly parallel upper molar rows, which is reminiscent of the cranial morphology of Pliosiphneus lyratus, and therefore is probably a descendant of it.
Acknowledgements
We are grateful to Professors Qiu Zhuding, Zhang Zhaoqun and Li Qiang, and Mr. Zhou Wei of the IVPP and Professor Cai Baoquan of the Xiamen University for their hard work in the field. We would also like to express our gratitude to Ms. Shi Aijuan of the IVPP for all the drawings she produced for us. The research was supported by the Strategic Priority Research Program of Chinese Academy of Sciences (grant no. XDB26000000), and the State Key Laboratory of Palaeobiology and Stratigraphy (Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (grant no. 173132).参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 7]
[本文引用: 4]
[本文引用: 15]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 7]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 4]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
DOIURL [本文引用: 6]
[本文引用: 2]
[本文引用: 2]
[本文引用: 12]
[本文引用: 6]
[本文引用: 5]
[本文引用: 9]
[本文引用: 2]
[本文引用: 6]
PMID [本文引用: 1]

The timing of the earliest habitation and oldest stone technologies in different regions of the world remains a contentious topic in the study of human evolution. Here we contribute to this debate with detailed magnetostratigraphic results on two exposed parallel sections of lacustrine sediments at Xiaochangliang in the Nihewan Basin, north China; these results place stringent controls on the age of Palaeolithic stone artifacts that were originally reported over two decades ago. Our palaeomagnetic findings indicate that the artifact layer resides in a reverse polarity magnetozone bounded by the Olduvai and Jaramillo subchrons. Coupled with an estimated rate of sedimentation, these findings constrain the layer's age to roughly 1.36 million years ago. This result represents the age of the oldest known stone assemblage comprising recognizable types of Palaeolithic tool in east Asia, and the earliest definite occupation in this region as far north as 40 degrees N.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}