 ,1,2, GavinYOUNG3, 胡雨致3,4, 乔妥1,2, 朱敏,1,2,5
,1,2, GavinYOUNG3, 胡雨致3,4, 乔妥1,2, 朱敏,1,2,5The posterior cranial portion of the earliest known tetrapodomorph Tungsenia paradoxa and the early evolution of tetrapodomorph endocrania
LU Jing,1,2, Gavin YOUNG3, HU Yu-Zhi3,4, QIAO Tuo1,2, ZHU Min,1,2,5收稿日期:2018-08-25网络出版日期:2019-04-20
| 基金资助: |
Corresponding authors: *lujing@ivpp.ac.cn,zhumin@ivpp.ac.cn
Received:2018-08-25Online:2019-04-20

摘要
首次报道了产于云南昭通早泥盆世布拉格期(约4.09亿年前)最古老的四足形类——奇异东生鱼(Tungsenia paradoxa)头后半部耳枕区的化石材料。东生鱼的耳枕区特征与早期的肺鱼形类,如杨氏鱼非常相似(如具发育的颈下脊,向耳突,以及不发育的桥窝等), 同时也展示出与之后更为进步的四足形类相似的特征(如背侧动脉汇合于颅后,具分离的腹中板等)。这些特征的镶嵌为厘清早期四足形类脑颅特征的获得序列提供了重要的数据。将新获得的东生鱼形态数据加入包含主要早期四足形类分类单元及形态特征的矩阵中进行系统发育分析,结果仍支持东生鱼位于四足动物支系最基干位置。东生鱼耳枕区新材料的发现填补了对早期四足形类脑颅耳枕区结构认识的空白,为认识四足形类后顶甲膜质骨型式及脑颅耳枕区的特征演化提供了新的证据。
关键词:
Abstract
Here the posterior cranial portion of the tetrapodomorph Tungsenia from the Lower Devonian (Pragian, ~409 million years ago) of Yunnan, southwest China, is reported for the first time. The pattern of posterior skull roof and the morphology of the otoccipital region of the neurocranium are described in detail, providing precious insight into the combination of cranial characters of the earliest known tetrapodomorph to date. The posterior cranium of Tungsenia displays a mosaic of features previously linked either to basal dipnomorphs such as Youngolepis (e.g., the well-developed subjugular ridge, the strong adotic process, and the poorly developed fossa bridgei) or to typical tetrapodomorphs (e.g., the lateral dorsal aortae commenced from the median dorsal aorta postcranially). The independent ventral arcual plate is also found in the advanced tetrapodomorph Eusthenopteron. The new endocranial material of Tungsenia further fills in the morphological gap between Tetrapodomorpha (tetrapod lineage) and Dipnomorpha (lungfish lineage) and unveils the sequence of character acquisition during the initial diversification of the tetrapod lineage. The new phylogenetic analysis strongly supports the basalmost position of Tungsenia amongst the tetrapod lineage.
Keywords:
PDF (1649KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
卢静, GavinYOUNG, 胡雨致, 乔妥, 朱敏. 云南昭通早泥盆世四足形类奇异东生鱼后顶甲及脑颅耳枕区化石兼论早期四足形类脑颅特征演化. 古脊椎动物学报[J], 2019, 57(2): 93-104 DOI:10.19615/j.cnki.1000-3118.181031
LU Jing, Gavin YOUNG, HU Yu-Zhi, QIAO Tuo, ZHU Min.
1 Introduction
Tetrapodomorpha, also known as the tetrapod lineage or tetrapod total group, includes all representatives that are more closely related to living tetrapods than to living lungfishes (Ahlberg, 1991). Hitherto, only two undisputed Early Devonian stem-tetrapods were known: Kenichthys and Tungsenia from Yunnan, China (Chang and Zhu, 1993; Zhu and Ahlberg, 2004; Lu et al., 2012), but the otoccipital region of these two early representatives has never been described due to the lack of sufficient material (Chang and Zhu, 1993; Zhu and Ahlberg, 2004; Lu et al., 2012). The recent discovery of the oldest known tetrapodomorph fish, Tungsenia, provides unique information to help improve our understanding of the origin and early evolution of the tetrapod lineage and additional valuable information based on the well-preserved anterior cranial region (ethmosphenoid shield), lower jaws, and cheek bones (Lu et al., 2012). However, our knowledge of the posterior cranial portion (otoccipital shield) of Tungsenia and the early evolution of this region in the tetrapod lineage remains limited due to the scarcity of known fossils.During fieldwork conducted from 2011-2015, we collected more Tungsenia cranial materials from the same site, including a well-preserved posterior cranial portion (IVPP V 15132.50). The new posterior cranial portion material possesses fine pores on the skull roof and the size of the posterior cranial portion is comparable with the anterior cranial portion of Tungsenia. Based on the shared ornamentation and comparable size with the reported anterior cranial portions, we assigned the new posterior cranial portion to Tungsenia.
After preparation, we found that the posterior cranial portion of Tungsenia is similar to that of Youngolepis in having a well-developed subjugular ridge and adotic process (Chang, 1982), while other endocranial features are quite close to those of ‘osteolepids’. Such combination of characters shed light on the early evolution of the tetrapod lineage.
2 Materials and methods
All specimens of Tungsenia are housed in the collections of the Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China (IVPP). The specimen was mechanically prepared to remove rock matrix.High-resolution computed tomography The posterior cranial portion (IVPP V 15132.50) was scanned at IVPP, using 225 kV microCT. The specimen was scanned with a beam energy of 180 kV and a flux of 100 mA at a detector resolution of 7.4 μm per pixel, using a 1440° rotation with a step size of 0.25° and an unfiltered aluminum reflection target. A total of 1440 transmission images were reconstructed in a 2048 × 2048 matrix of 1536 slices. Scan data was analyzed using Mimics v.18.01 (
Phylogenetic analysis The phylogenetic analysis was performed on a matrix of 263 characters scored for 38 taxa (see on-line supplementary appendix 3; taxon-by-character matrix). All characters were treated as unordered and weighted equally. The resulting data matrix was subjected to the parsimony analysis in PAUP* (version 4.0b10) (Swofford et al., 2001) using a heuristic search, with Guiyu, Psarolepis and Achoania (basal osteichthyans) specified as the outgroup. The analysis generated 24 most parsimonious trees (MPTs) with 581 steps (Consistency index (CI) = 0.5232; Homoplasy index (HI) = 0.4768; Retention index (RI) = 0.7247; Rescaled consistency index (RC) = 0.3792). All trees show Tungsenia as the most basal stem-tetrapod. The strict consensus tree highlights the poor phylogenetic resolution of the lower part of the tetrapodomorph lineage.
3 Systematic paleontology
Osteichthyes Huxley, 1880Sarcopterygii Romer, 1955
Tetrapodomorpha Ahlberg, 1991 (= total group Tetrapoda)
TungseniaLu et al., 2012
Tungsenia paradoxa (Lu et al., 2012)
Holotype IVPP V 10687, an anterior cranial portion.
Type locality and horizon Zhaotong, Yunnan Province, China; Posongchong Formation (Pragian, Early Devonian).
New material IVPP V 15132.50, a posterior cranial portion.
Locality and horizon The same as type locality and horizon.
Diagnosis (emended) The posterior cranial portion (otoccipital or postparietal shield) is as long as it is wide; W-shaped postparietals extend much further posteriorly than the tabulars; the tabulars turn down anterolaterally, forming the inner margin of the spiracular slit; the extratemporals are large; the basicranial fenestra is large; the ventral fenestra is present, with well-developed subjugular; a notch for the middle process of median extrascapular is present; the posterior dorsal fontanelle is present; the fossa bridgei is poorly developed.
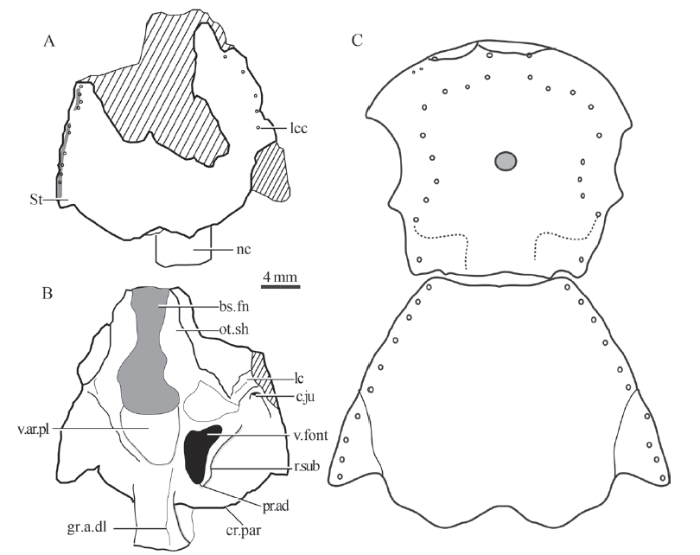
Description The posterior cranial portion of Tungsenia (IVPP V 15132.50) is well-preserved except for the broken left anterior part of the shield (Figs. 1-3). The length of the shield is about 8 mm, and basically equal to its width. In dorsal view, the dermal skull roof is covered with a continuous layer of cosmine and the sutures between the postparietals, and the marginal bones of the skull roof (supratemporals, temporals, and tabulars) cannot be traced. The typical anterolateral overlapped area for the postorbital plate of the anterior cranial portion in tetrapodomorphs has not be observed due to incomplete preservation. The posterior edge of the skull roof is well-preserved, showing a curved embayment in the mesial part of the postparietals and this may indicate the outline of the anterior margin of the median extrascapular. The tabular and the outer part of the postparietal have a slit in the edge of the bone, the lip below the slit extends posteriorly to form a shelf for the articulation of the lateral extrascapulars. A distinct posterolateral protruding corner recalls the presence of extratemporal, which is common in ‘osteolepiforms’ (Jarvik, 1948). This extratemporal, if present, is fused with the tabular and thus highly supports the hypothesis that the extratemporal is part of the skull table, rather than being an element of hyoid area. The pores of the otic branch of the main sensory canal indicate that the course of the lateral line canal goes through the marginal bones of the skull roof. However, it should be noted that the lateral line sensory canal goes through the lateral margin of the extratemporal, which differs from other known sarcopterygians in which the sensory canal passes through the tabular. No pore cluster or pit-line has been found in the dermal skull roof.
Fig. 1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1Photos of the new postparietal shield (IVPP V 15132.50) in dorsal (A) and ventral (B) views
Abbreviations: bs.fn. basicranial fenestra; cr.par. crista parotica; gr.a.dl. groove for lateral dorsal aorta; lc. lateral commissure; lcc. main lateral line canal; nc. notochord; ot.sh. otic shelf; r.sub. subjugular ridge; St. supratemporal; v.ar.pl. ventral arcual plate; v.font. vestibular fontanelle
In ventral view, the otoccipital region of Tungsenia is composed of two main elements: the basicranial fenestra (bs.fn, Figs. 1B, 2B) and the otoccipital ossification (Figs. 1, 2). The basicranial fenestra is oblong in outline and occupies about a half of the whole length of the otoccipital region. Anteriorly, it is bounded by the posteroventral parts of the ethmosphenoid below the notochordal pit. Laterally, the fenestra reaches the medial margin of the anteroventral articular process of the otoccipital and of the otic shelf (ot.sh, Fig. 2B), while posteriorly its extension is delimited by the independent ventral arcual plate, which is anterior to the otoccipital ossification. The ventral margin of the otic shelf is not fully preserved, so the natural shape of the otic shelf remains unclear. The lateral commissure continues with the otic shelf, extending laterally to the skull roof. The facets for the hyomandibular articulations are not well-shown due to the poor preservation. An independent arched ventral arcual plate is present anterior to the otoccipital, and the suture between the ventral arcual plate and the otoccipital is well shown. The otoccipital is of a polygonal, symmetrical shape and represents a single stout basicranial plate right behind the ventral arcual plate. A paired longitudinal groove for the dorsal aorta (gr.a.dl, Figs. 2B, 3B) at the posterolateral margin is obvious. At the same level with the ventral arcual plate, a large vestibular fontanelle (v.font, Fig. 2B) is present on the lateral braincase wall. A well-developed subjugular ridge (r.sub, Figs. 2B, 3) forms the lateral and posterior boundary for the vestibular fontanelle. The adotic process (pr.ad, Fig. 3) is situated at the end of the subjugular ridge, posterolateral to the otic capsule, like that of Youngolepis (Chang, 1982). According to the relative position with the vestibular fontanelle, the subjugular ridge corresponds to the jugular ridge in Eusthenopteron, and the adotic process could compare to the areas articulating with branchial arches in Eusthenopteron (Jarvik, 1980). The dorsal margin of the jugular groove is bounded off by a suprajugular ridge, as that in Youngolepis. Due to the preservation, the foramina for the nerves or vessels in the otoccipital region are not visible.
Fig. 2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Illustrative drawings of the posterior cranial portion of Tungsenia paradoxa (IVPP V 15132.50)
A. dorsal view; B. ventral view; C. the restoration of the whole shield showing the length proportion of the parietal shield vs the postparietal shield Abbreviations: c.ju. jugular canal; pr.ad. adotic process For other abbreviations see
In posterior view, the specimen is deformed in the right axis. The dorsomedian part of the posterior face of otoccipital has a large foramen, which represents the posterior dorsal fontanelle (pdf, Fig. 3B, D), a structure commonly present in osteichthyans (Chang, 1982; Yu, 1998; Zhu and Yu, 2002; Lu et al., 2016, 2017; Clement et al., 2018). Lateral to the posterior dorsal fontanelle, there are well-defined paired depressions called fossa tectosynotica (f.tect, Fig. 3). The poorly developed fossa bridgei (f.br, Fig. 3) is lateral to the fossa tectosynotica. The fossa bridgei and the fossa tectosynotica are separated by a short ridge. The occipitalis lateralis fissure, indicating the boundary between the otic and occipital regions, could not be traced. The suture between the notochordal canal (nc, Fig. 3) and the foramen magnum (fm, Fig. 3) is not visible.
Fig. 3
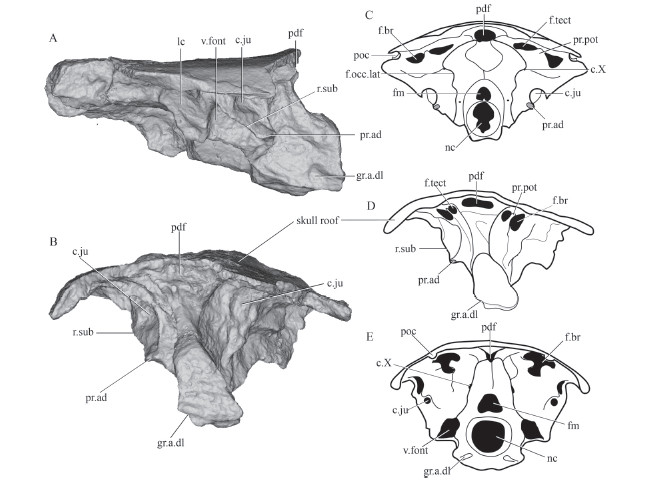 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3Rendered posterior cranial portion (IVPP V 15132.50) and comparisons of selected rhipidistian endocrania in posterior view
A. right lateral view of Tungsenia; B. posterior view of Tungsenia; C. line drawing of the endocranium of Youngolepis in posterior view, modified from Chang, 1982; D. line drawing of the endocranium of Tungsenia in posterior view; E. line drawing of the endocranium of Gogonasus in posterior view, modified from Long et al., 1997. Abbreviations: c.X. canal for vagus nerve; f.br. fossa bridgei; fm. foramen magnum; f.occ.lat. lateral occipital fissure; f.tect. fossa tectosynotica; pdf. posterior dorsal fontanelle; poc. postotic sensory canal; pr.pot. postotical process. For other abbreviations see Figs. 1, 2. Not to scale
4 Discussion
4.1 Phylogenetic implications of the tetrapodomorph evolution
To re-examine the phylogenetic position of Tungsenia in light of the new morphological data, we conducted phylogenetic analysis using a modified data matrix from Lu et al. 2012 (see on-line supplementary appendix 1). Our parsimony analysis recovers 12 trees with a length of 581 steps (Fig. 4) (character optimization for a single MPT given in on-line supplementary appendix 2). Our result shows Tungsenia is consistently resolved as the basalmost stem tetrapod and shares with the other tetrapodomorphs the following unambiguous characters:(1) The exit of pituitary vein situated in front of the basipterygoid (Ch. 50, code 0). represents a derived arrangement for the tetrapod lineage. In other sarcopterygians (e.g. Styloichthys, Youngolepis), the pituitary vein foramen lies dorsal to the basipterygoid process.
(2) In tetrapodomorphs, the accessory fenestration in the otic capsule is absent (Ch. 62, code 0). The accessory fenestration is present in Styloichthys, coelacanths, the onychodont Qingmenodus, and basal dipnomorphs.
(3) The parasymphysial tooth whorl is a feature of non-tetrapodomorph sarcopterygians. In tetrapdomorphs, the parasymphysial plate is either with a tooth pavement or paired tusks (Ch. 139, code 1).
(4) In tetrapodomorphs, the anterior mandibular (precoronoid) fossa is widely present (Ch. 161, code 1);
(5) The otic and supraorbital canals are in contact in the tetrapod lineage through the dermal demosphenoid bone (Ch. 183, code 1).
Previous to the discovery of Tungsenia, the tetrapodomorph clade was supported by the following characters:
(1) The absence of internasal pits (Ch. 40). Tungsenia is the only known tetrapodomorph with paired internasal pits. Usually, there is an unpaired prenasal pit present in the snout region of tetrapodomorphs.
(2) The intracranial joint running through the profundus foramen (Ch. 55). In Tungsenia and some basal dipnomorphs, such as Powichthys, the profundus foramen is in front of the intracranial joint.
(3) Posttemporal fossae present (Ch. 67). The posttemporal fossa is poorly developed in Tungsenia and in the basal dipnomorph Youngolepis.
(4) The extent of crista parotica not reaching the posterior margin of the tabular (Ch. 68).
(5) Parasphenoid slender and splint in shape (Ch. 96). The shape of the parasphenoid in Tungsenia is quite different compared with other tetrapodomorphs. A broad parasphenoid is present in Tungsenia and other primitive sarcopterygians.
(6) Lateral sides of the parasphenoid converging anteriorly (Ch. 97). The lateral sides of parasphenoid in Tungsenia are similar to that of Styloichthys and basal dipnomorphs (e.g., Youngolepis, Powichthys), which are parallel or slightly diverging anteriorly.
The strict consensus tree shows that the ‘lower part’ of the tetrapodomorph lineage - formed by a heterogenous group traditionally referred to as ‘osteolepiforms’ - remains significantly problematic. The evolutionary relationship of the four ‘osteolepiform’ groups (i.e. osteolepids, megalepids, canowindrids, and tristichopterids) is still in polytomy due to lack of sufficient information, particularly from key taxa representing early diverging members of the present groups. Many known ‘osteolepiform’ representatives were entirely bound to the Middle-Late Devonian. The rhizodonts surveyed in the current analysis look stable as the second basalmost position of the lineage. However, it is still worthwhile to note that the anatomical similarities between the rhizodonts and advanced representatives of the tetrapodomorph lineage should not be underestimated and indicate that the possibility that the group might occupy a much higher position in the tree still exists (Zhu et al. 2017).
Fig. 4
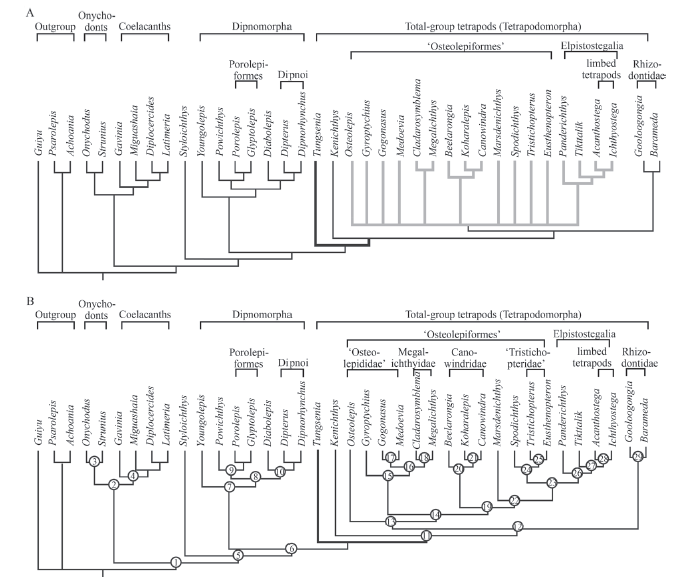 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 4Phylogenetic analysis with new codings of Tungsenia paradoxa shown in the strict consensus tree and one selected shortest tree
A. the strict consensus tree of the 24 shortest trees; B. one selected shortest tree the same as 50% majority-rule tree showing the apomorphic features at different nodes. More details see on-line supplementary appendix 2
Hitherto, only two undisputed Early Devonian tetrapodomorph fishes were known (Chang and Zhu, 1993; Zhu and Ahlberg, 2004; Lu et al., 2012). This apparent low diversity is now augmented by the Australian Early Devonian tetrapodomorph ‘Gyroptychius?’ australis (Young and Gorter, 1981), which shares some unique features with Kenichthys (Chang and Zhu, 1993). In addition, numerous new tetrapodomorph fishes from the Early-Middle Devonian have been found in Hatchery Creek, New South Wales, Australia, have never been properly described (Hunt and Young, 2012). Further investigations on the early diverging members of ‘osteolepiform’ groups are needed and may provide crucial information to resolve taxon instability in the tetrapod lineage.
4.2 Endocranial evolution in tetrapodomorphs
The endocranium is one of the most important components of the body plan, and more than 70% of characters in previous phylogenetic analyses are related to cranial features. However, due to the lack of primitive tetrapodomorph material, our understanding on the endocranial evolution in the tetrapod lineage remains obscure. The discovery of Tungsenia, the basalmost and oldest known tetrapodomorph, provides a unique opportunity to investigate the endocranial evolution at the initial stage of the tetrapod lineage (Fig. 5).The endocranium of Tungsenia shares many features with Youngolepis (Chang, 1982), as well as to a lesser extent Styloichthys (Zhu and Yu, 2002) and Powichthys (Jessen, 1980), such as the ventrally opened anterior nostril; the presence of paired internasal cavities; the elongate and broad parasphenoid, which is parallel or slightly diverging anteriorly; the vomers separated by the parasphenoid; the attachment for the basicranial muscle covered by the parasphenoid, which could not be seen in ventral view; the broad orbital tectum, and the well-developed basipterygoid.
Furthermore, the typical crista suspendens is poorly developed in Tungsenia, which is different from the well-developed crista suspendens in other tetrapodomorphs (e.g., Kenichthys, Gogonasus, Osteolepis, Meglichthys) (Thomson, 1964; Fox et al., 1995; Lebedev, 1995; Long et al., 1997; Zhu and Ahlberg, 2004; Borgen and Nakrem, 2016). The similarity between Tungsenia and other tetrapodomorph fishes lies in the opening of the pituitary vein, which is situated at the base of the basipterygoid (Coates and Friedman, 2010). However, the exit for the profundus nerve in Tungsenia opens in the posterior part of the ethmosphenoid region, which recalls the condition in Youngolepis. It is worth mentioning that the parasphenoid in Tungsenia, Styloichthys, Youngolepis, and some basal tetrapodomorphs (e.g., Kenichthys, and an undescribed Early Devonian tetrapodomorph from Australiaextend posteriorly, reaching the ventral opening of the notochord canal and covering the attachment for the basicranial muscle (Chang, 1982; Zhu and Yu, 2002; Zhu and Ahlberg, 2004; Hunt and Young, 2012).
Although a large basicranial fenestra is present in Tungsenia, the general features of the otoccipital region quite agree with those of Youngolepis, such as the well-developed subjugular ridge, the strong adotic process situated at the end of the subjugular ridge, and the poorly developed fossa bridgei (Chang, 1982). On the other hand, Tungsenia agrees with other tetrapodomorphs in the lateral dorsal aortae emerging from the median dorsal aorta postcranially, thus the impression for the median dorsal aorta is not preserved on the ventral side of the occipital unit (Long et al., 1997). An independent ventral arcual plate is also seen in Eusthenopteron (Jarvik, 1980).
It is remarkable that Tungsenia displays more primitive rather than advanced features in the endocranium, which indicates that the tertapodomorph endocranium retains the primitive condition present at the initial stage of tetrapod evolution. The major identical endocranial modifications of the tetrapod lineage, such as the elongate of the olfactory tract, anterior-posterior situated pineal and parapineal organs, anteriorly converging sword-like parasphenoid, well-developed crista suspendens, and well-developed fossa bridgei occurred later after the Pragian stage of the Devonian period (Jarvik, 1980; Zhu and Ahlberg, 2004).
Fig. 5
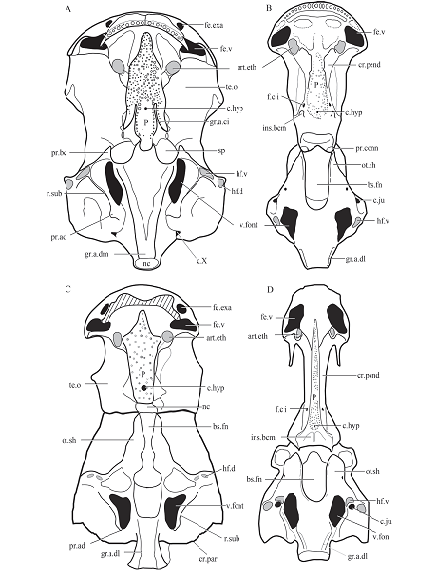 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 5Comparisons of rhipidistian neurocrania in ventral view
A. Youngolepis (after Chang, 1982); B. Gogonasus andrewsae (after Long et al., 1997); C. Tungsenia paradoxa; D. Eusthenopteron foordi (after Jarvik, 1980) Abbreviations: art.eth. ethmoid articulation for the palatoquadrate; c.hyp. canal for buccohypophysial duct; c.ju. jugular canal; cr.pend. crista suspendens; f.c.i. foramen for the internal carotid artery; fe.exa. fenestra exonarina anterior; fe.v. fenestra ventralis; gr.a.ci. groove for the internal carotid artery; gr.a.dm. groove for median dorsal aorta; hf.d. dorsal hyomandibular facet; hf.v. ventral hyomandibular facet; ins.bcm. insertion area for the basicranial muscle; P. parasphenoid; sp. space; te.o. orbital tectum For other abbreviations, see Figs. 1 and 2. Not to scale
5 Conclusion
The posterior cranial portion of Tungsenia provides unique evidence for the under-standing of otoccipital region evolution in the tetrapod lineage, and the new data strongly supports the basalmost position of Tungsenia among tetrapodomorphs. However, the phylogenetic relationships of the lower part of the tetrapod lineage are still not well resolved. Further investigations into the early representatives that occurred from both China and East Gondwana (Australia-Antarctica) will help to resolve ancestral conditions for tetrapod origins.Acknowledgments
We thank XIONG C-H for specimen preparation. HOU Y-M for CT scanning. This work was supported by the National Natural Science Foundation of China (41872023, 41472016), Key Research Program of Frontier Sciences, CAS, Grant No. QYZDB-SSW-DQC040.Supplementary material can be found on the website of Vertebrata PalAsiatica (
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 7]
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
[本文引用: 4]
[本文引用: 7]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 6]
[本文引用: 3]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}