摘要: 位于新生和成年哺乳动物延髓腹外侧区的前包钦格复合体被认为是呼吸节律产生的中枢. 正常状态下呼吸节律是均匀而整齐的, 而病理状态下呼吸节律会发生变化, 因此研究呼吸节律产生的动力学机制及其控制有重要意义. 本文基于前包钦格复合体中胞体-树突耦合神经元模型, 利用相平面分析、分岔分析、快慢动力学分析以及ISI (峰峰间期)分岔序列等方法, 研究了在钙离子动力学及
L IP3 (内质网泄漏渗透率)的影响下,
L IP3 的变化对耦合前包钦格复合体神经元放电节律的重要影响, 并研究了模型中反相簇放电的模式及其转迁机制. 结果表明, 钙的周期性波动是混合簇放电产生的关键因素, 但不是混合簇放电产生的必要条件. 本文的研究方法也可以应用于其他多时间尺度的神经系统中.
关键词: 前包钦格复合体 /
混合簇放电 /
分岔 /
快慢分析 English Abstract Dynamics of mixed bursting in coupled pre-B?tzinger complex Zhao Ya-Qi 1 ,Liu Mou-Tian 1 ,Zhao Yong 2 ,Duan Li-Xia 1 1.School of Science, North China University of Technology, Beijing 100144, China Fund Project: Project supported by the National Natural Science Foundation of China (Grant No. 11872003) and the Technology Research Fund Program for Key Discipline of North China University, China (Grant No. 110052972027/014)Received Date: 14 January 2021Accepted Date: 17 February 2021Available Online: 17 June 2021Published Online: 20 June 2021Abstract: The pre-B?tzinger complex, which is located at a ventrolateral medulla of human and mammal, is considered to be the center for the generation of respiratory rhythms. In a normal state, the respiratory rhythm is uniform and orderly. Otherwise, the respiratory rhythm will change to a pathological state. Therefore, the monitoring of respiratory rhythm is of great significance in monitoring the health. In this paper, according to a two-coupled model of pre-B?tzinger complex with calcium ion current, we investigate the generation and transition mechanism of anti-phase bursting synchronization by using phase-plane analysis, bifurcation and fast-slow decomposition. It is found that the pre-B?tzinger complex model can exhibit mixed bursting when calcium ion concentration is at steady state, which indicates that the oscillation of calcium is not a necessary condition for the generation of mixed bursting. This is quite different from the results obtained in previous studies, indicating that the mixed bursting is caused by the periodic fluctuations of calcium. The methods used in this paper can provide a new idea for investigating the dynamics of mixed bursting, and it can also be applied to the study of other neuronal systems on a multiple time scale.Keywords: pre-B?tzinger complex /mixed bursting /bifurcation /fast-slow analysis 全文HTML --> --> --> 1.引 言 呼吸节律的产生部位和原理一直是神经生物学研究领域中的热门课题. 近年来研究表明, 位于延髓中的前包钦格复合体是哺乳动物呼吸节律产生的关键部位[1 ,2 ] . Gourévitch和Mellen[3 ] 发现前包钦格复合体是新生大鼠腹侧呼吸柱网络活动的枢纽, 在功能不同的呼吸神经元中起到整合作用. 在生理实验和临床发现了很多不同类型的呼吸节律, 比如叹息样呼吸、陈氏呼吸等[4 -6 ] . 为了更好地研究呼吸节律, 相关的模型也在逐渐建立. 基于Hodgkin和Huxley[7 ] 建立的著名离子通道模型, Butera等[8 ,9 ] 建立了描述前包钦格复合体动作电位的两个最小的起搏神经元模型, 并研究了耦合神经元的频率控制和同步问题. 2004年, Kosmidis[10 ] 提出了一个切片的计算模型, 表明呼吸样节律可以在非起搏神经元的兴奋性网络模型中产生. Smith等[11 ] 建立了哺乳动物脑干的功能结构呼吸网络模型, 用以模拟三种振荡机理产生的层次结构. 2009年, Rubin等[12 ] 基于实验现象改进了前包钦格复合体的神经元模型, 并且在此基础上研究了反复激励和钙激活的非特异性阳离子电流${I_{{\rm{CAN}}}}$ 对放电模式的影响. 2011年, Toporikova和Butera[13 ] 提出了前包钦格复合体的双室数学模型(TB模型), 其中包括两种独立的簇放电产生机制, 即依赖于持续钠离子电流${I_{{\rm{Nap}}}}$ 的胞体簇放电和依赖于细胞内钙浓度振荡的树突簇放电. Park和Rubin[14 ] 在2013年对TB模型进行了改进, 建立了保持之前双室模型特征的单室模型, 并给出了胞体簇放电(S簇放电)、树突簇放电(D簇放电)和胞体-树突簇放电(SD-簇放电)产生及转迁的数学解释. Duan等[15 ] 结合Rubin单室模型和Butera等建立的网络模型, 构建了一个耦合单室神经元网络模型, 并研究了网络中不同类型簇放电的转迁机理.[16 ] . 快慢分解和分岔分析已经广泛应用于神经科学模型的研究中. 模型系统中的变量往往可以分为快、慢不同的时间尺度, 利用快慢分岔分析可以很好地解释不同簇放电产生的动力学机理[16 ,17 ] . 根据簇放电产生的分岔机理, Izhikevich[18 ] 将簇放电进行了归纳和分类.[19 ] 研究了单个神经元在钙离子影响下的混合簇放电模式及其鲁棒性. Lü等[20 ] 研究了钠电导$({g_{{\rm{Na}}}})$ 和钾电导$({g_{\rm{K}}})$ 对前包钦格复合体中混合簇放电模式的影响, 并进一步阐明参数${g_{\rm{K}}}$ 和${g_{\rm{L}}}$ 对混合簇放电中胞体簇放电个数的影响. 簇放电的同步行为更为复杂, 王付霞和谢勇[21 ] 研究了修正的 Morris-Lecar 神经元模型中“Hopf/homoclinic”簇放电和“SubHopf/homoclinic”簇放电之间的同步行为. 在前包钦格复合体模型中, 当引入突触耦合时, 神经元会产生同相和反相两种簇放电模式[22 ] , 且反相簇放电的模式更为复杂[15 ,23 ] . Wang等[23 ] 研究了钾电流对耦合前包钦格复合体中混合簇放电模式的影响, 并利用快慢分解和分岔分析, 研究了混合簇放电的动力学机制以及不同混合簇放电模式之间的转迁机理. 杨永霞等[24 ] 研究了兴奋性耦合强度对前包钦格复合体模型中同相同步和反相同步放电模式的影响, 并且给出了从簇放电到峰放电节律的同步转迁规律及复杂的分岔机制.[25 ,26 ] . 1990年, Goldbeter和Dupont[27 ] 建立了研究钙振荡的数学模型, 细胞钙动力学的理论研究与实验观察相结合, 使该学科领域的研究取得了深入的发展. 1995年, Jouaville等[28 ] 研究了线粒体对于非洲爪蟾卵母细胞内钙波斑图形成的影响, 实验表明, 由此形成的钙波具有较长的周期、较大的振幅和较高的波速(如当卵母细胞中通过注射 IP3S3 和丙酮酸/苹果酸建立了${\rm{C}}{{\rm{a}}^{2 + }}$ 波活动, 随后注射鱼藤酮逆转钙波活性, 可使得周期从$ (39.6\pm 9.0) \; \rm{s}$ 减少至$ (10.6\!\pm\! 2.3) \; \rm{s}$ ). 后来, Falcke 等[29 ] 利用 Othmer-Tang的钙离子动力学模型附加线粒体离子循环动力学方程, 重现了 Jouaville等[28 ] 的实验发现(不同刺激水平下重复${\rm{C}}{{\rm{a}}^{2 + }}$ 波的模拟显示了许多其他可兴奋系统中观察到的${\rm{C}}{{\rm{a}}^{2 + }}$ 浓度的波动与频率的关系, 两者为负相关关系, 当${\rm{C}}{{\rm{a}}^{2 + }}$ 浓度的波动范围是$(0.75 \pm 0.1)—({0}.95 \pm 0.1) \; {\rm{{\text{μ}} mol/L}}$ , 频率的变化范围是$(0.09\pm 0.01)—(0.02\pm 0.01) \; {\rm{s}}^{-1}$ ). 施小民[30 ] 首次验证了线粒体钙循环的参与可提高胞质钙振荡的频率的实验结论, 并且认定, 线粒体的活性越大, 胞质钙振荡的频率越高.[19 ] . 钙离子振荡将导致包括鞍结分岔、Hopf 分岔在内的快子系统平衡点的分岔曲线的相对位置发生改变, 从而产生了复杂模式簇放电[19 ,23 ] . 而细胞内钙离子浓度的影响因素有三磷酸肌醇(${\rm{I}}{{\rm{P}}_3}$ )、内质网泄漏渗透率(${L_{{\rm{I}}{{\rm{P}}_3}}}$ )等. 在一定范围内, 随着${L_{{\rm{I}}{{\rm{P}}_3}}}($ 或${\rm{I}}{{\rm{P}}_3})$ 的增加, 非特异性阳离子电流${I_{{\rm{CAN}}}}$ (${I_{{\rm{CAN}}}}$ 可以通过放大突触电流以产生强大的吸气驱动电位, 是前包钦格复合体神经元中强大的吸气驱动电位的基础$)$ 被激活, 导致钙离子浓度发生周期性波动. 随着钙离子浓度的波动, 变量的快慢尺度发生改变, 从而导致系统出现混合簇放电[31 ] . 虽然关于混合簇放电已经有了一些研究结果, 但是在耦合神经元系统中, 混合簇放电模式的产生机制及钙动力学更为复杂, 值得进一步研究.$ {L}_{{\mathrm{I}\mathrm{P}}_{3}} $ 对混合簇放电模式及其转迁机制的影响. 本文仅研究反相簇放电. 文章结构如下: 第2 节主要介绍耦合前包钦格复合体神经元的数学模型; 第3 节探讨参数$ {L}_{{\mathrm{I}\mathrm{P}}_{3}} $ 的变化对反相簇放电模式的影响; 最后一节给出本文的结论. 本文中分岔计算采用 XPPAUT 软件[32 ] .2.模型介绍 本文研究的耦合前包钦格复合体神经元双室模型(TB)[8 ,9 ,14 ,15 ] 描述如下:$i, j = \{ 1, 2\} $ 并且$i \ne j$ . ${C_{\rm{m}}}$ 是膜电容, ${V_i}$ 表示膜电位, ${h_i}$ 和${n_i}$ 是电压依赖性的门控变量, ${s_i}$ 是突触耦合变量, ${[{\rm{Ca}}]_i}$ 表示细胞内钙浓度, ${l_i}$ 表示未被灭活的${\rm{I}}{{\rm{P}}_3}$ 通道的部分, 该通道会影响由${J_{{\rm{E}}{{\rm{R}}_{{\rm{IN}}}}}}$ 和${J_{{\rm{E}}{{\rm{R}}_{{\rm{OUT}}}}}}$ 表示的胞浆和内质网(ER)之间的钙通量, (2 )式中$\varepsilon $ 是调节参数. ${x_\infty }({V_i})$ $(x \in \{ {m_{\rm{p}}}, m, h, n, s\} )$ 和${\tau _x}({V_i}) $ $(x \in \{ h, n, s\} )$ 分别代表$x$ 的稳态电压依赖激(失)活函数和电压依赖时间常数. 方程(1 )—(6 )是全系统, 方程(1 )—(3 )是胞体子系统, 方程(5 )和方程(6 )是树突子系统[14 ] . 各离子电流的表达式如下:$i, j = \{ 1, 2\};\; {I_{{\rm{NaP}}}}, {I_{{\rm{CAN}}}}, {I_{{\rm{Na}}}}, {I_{\rm{K}}}, {I_{\rm{L}}}, {I_{{\rm{syn \text- e}}}}, {I_{{\rm{tonic \text- e}}}}$ 分别代表持续钠电流、钙激活的非特异性阳离子电流、钠离子电流、钾离子电流、泄漏电流、耦合神经元网络连接所产生的电流以及细胞膜受到兴奋性刺激所产生的电流; ${g_{{\rm{NaP}}}}, \; {g_{{\rm{Na}}}}, \;{g_{\rm{K}}}, \; {g_{\rm{L}}}, \; {g_{{\rm{tonic \text- e}}}}, \; {g_{{\rm{syn \text- e}}}}, \; $ $ {g_{{\rm{CAN}}}}$ 分别为各通道的最大电导; ${E_{{\rm{Na}}}}, \; {E_{\rm{L}}}, \; {E_{{\rm{syn \text- e}}}}$ 分别是相应的平衡电位. 其他变量的表达式及参数值见附录A .3.结果与分析 在耦合前包钦格复合体神经元模型中, 在相同的参数集下, 对不同的初始条件, 簇放电通常包括两种类型, 即同相簇放电与反相簇放电. 当两个神经元的初始值相同时, 系统表现为同相簇放电. 反之, 系统会表现为反相簇放电[23 ,33 ] . 放电模式的影响因素有很多, 如不同浓度的离子电流、三磷酸肌醇(${\rm{I}}{{\rm{P}}_3}$ )、内质网的泄露渗透率(${L_{{\rm{I}}{{\rm{P}}_3}}}$ )等. 本文主要研究内质网的渗透性参数${L_{{\rm{I}}{{\rm{P}}_3}}}$ 对耦合前包钦格复合体神经元模型中反相复杂簇放电模式的影响.3.1.树突子系统中钙动力学 3.1.树突子系统中钙动力学 ${\rm{C}}{{\rm{a}}^{2 + }}$ 的动力学仅由方程(5 )和方程(6 )决定, 即细胞内钙动力系统仅限于树突子系统, 独立于胞体子系统. 树突子系统(方程(5 )和方程(6 ))相对于参数${L_{{\rm{I}}{{\rm{P}}_3}}}$ 的分岔如图1(a) 所示. 其中黑色曲线由平衡点组成, 黑色曲线上点${{\rm{F}}_2}$ 下方的分支自上而下分别为鞍点(黑色虚线)、稳定结点(黑色实线)和鞍点(黑色虚线)组成, 上支由不稳定焦点和稳定焦点构成.图 1 参数${L_{{\rm{I}}{{\rm{P}}_3}}}$ 变化时树突子系统(方程(5 )和方程(6 ))的动力学 (a) $ [{{\rm{IP}} _3}] = 1.2 \; {\rm{{\text{μ}} mol/L}} $ 时的单参数分岔, 其中黑色曲线表示平衡点, 红色曲线表示周期轨道的最大值和最小值; ${{\rm{F}}_1}$ , ${{\rm{F}}_{\rm{2}}}$ 表示平衡点的鞍结分岔; ${\rm{HB}}$ 表示Hopf分岔; (b) 极限环的变化周期; (c) 极限环的变化频率. 参数值见附录A Figure1. Dynamics of the dendritic subsystem with parameter ${L_{{\rm{I}}{{\rm{P}}_3}}}$ : (a) One-parameter bifurcation of dendritic subsystem with $ [{{\rm{IP}} _3}] = 1.2 \; {\rm{{\text{μ}} mol/L}} $ . Black curve indicates equilibrium points and red curve indicates maximum and minimum values of periodic orbits respectively. ${\operatorname{F} _1}$ and ${\operatorname{F} _2}$ are the fold bifurcation points, ${\rm{HB}}$ is the Hopf bifurcation. (b) Changes of period of limit cycle with the parameter ${L_{{\rm{I}}{{\rm{P}}_3}}}$ . (c) Changes of frequency of limit cycle with the parameter ${L_{{\rm{I}}{{\rm{P}}_3}}}$ . Defaulted parameter values are shown in Appendix A .${L_{{\rm{I}}{{\rm{P}}_3}}} = 13.9694\; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ 时, Hopf分岔$({\rm{H}}{{\rm{B}}_1})$ 产生, 同时伴随着稳定的极限环产生(红色曲线). 另外还存在一个几乎与不变圆上的鞍结分岔(SNIC)重合的Hopf分岔(${\rm{H}}{{\rm{B}}_2}$ , ${L_{{\rm{I}}{{\rm{P}}_3}}}\!=\! 0.1319 \,{\rm{PL}}\cdot {{\rm{s}}^{ - 1}}$ ), 并对应存在范围极小、可以忽略不记的不稳定极限环(其不会影响本文所讨论的内容). 当${L_{{\rm{I}}{{\rm{P}}_3}}} = $ $ 0.1317\; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ 时, 稳定极限环经由不变圆上的鞍结分岔(SNIC)消失. 因此当${L_{{\rm{I}}{{\rm{P}}_3}}} > $ $ 13.9694 \;{\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ 时, 树突子系统存在稳定的平衡点; 当$0.1317 \; {\rm{PL}}\! \cdot\! {{\rm{s}}^{ - 1}} < $ $ {L_{{\rm{I}}{{\rm{P}}_3}}} < 13.9694\; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ 时, 树突子系统存在稳定的极限环, 即钙处于振荡状态. 此时极限环所对应的周期如图1(b) 所示.${L_{{\rm{I}}{{\rm{P}}_3}}}$ 的增加, 周期减小而频率增大, 具体如下: $ {L_{{\rm{I}}{{\rm{P}}_3}}} = 13.9694 \; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}} $ 时, Hopf分岔处极限环产生, 此时极限环的周期最小. 随着${L_{{\rm{I}}{{\rm{P}}_3}}}$ 的减小, 周期逐渐增大. 当$ {L_{{\rm{I}}{{\rm{P}}_3}}} = 0.1317 \; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}} $ 时, 极限环经由不变圆上的鞍结分岔消失, 周期趋于无穷, 如图1(b) 所示. 频率随${L_{{\rm{I}}{{\rm{P}}_3}}}$ 的变化如图1(c) 所示.$[{{\rm{IP}} _3}]$ 不同的情况下, 树突子系统(方程(5 )和方程(6 ))相对于参数${L_{{\rm{I}}{{\rm{P}}_3}}}$ 的分岔结构如图2(a) 所示. 随着$[{\rm{I}}{{\rm{P}}_3}]$ 的增加, 位于右侧的不变圆上的鞍结分岔变化很小, 但Hopf分岔向左移动, 不变圆上鞍结分岔点与Hopf分岔点之间的距离缩短. 从而导致稳定极限环的存在区域减小. 树突子系统(方程(5 )和方程(6 ))对$[{{\rm{IP}} _3}]$ 和${L_{{\rm{I}}{{\rm{P}}_3}}}$ 的双参数分岔如图2(b) 所示. ${\rm{G}}{{\rm{H}}_1}\;{\rm{(G}}{{\rm{H}}_2})$ 和${\rm{BT}}$ 分别表示余维-2分岔的广义Hopf分岔以及Bogdanov-Takens分岔. 随着$[{{\rm{IP}} _3}]$ 的增加, snic 曲线和Hopf分岔曲线(hb )的相对距离减小, 表明树突子系统稳定极限环的区域减小, 即钙振荡的区域变小. $[{{\rm{IP}} _3}]$ 分别取不同值时对应的树突子系统中关键点以及稳定极限环的具体存在区域如表1 所列.图 2 树突子系统 (方程(5 )和方程(6 ))的动力学分析 (a) 单参数分岔, 其中$ [{\rm{I}}{{\rm{P}}_3}] = 1, 1.05, 1.1, 1.2 \; {\rm{{\text{μ}} mol/L}} $ ; (b) $({L_{{\rm{I}}{{\rm{P}}_3}}}, $ $ [{{\rm{IP}} _3}])$ 平面上的双参数分岔, 分岔曲线包括平衡点的鞍结分岔($f$ ), Hopf分岔(hb )和不变圆上的鞍结分岔($snic$ ); (c) 图(b)的部分放大图Figure2. Dynamics analysis of the dendritic subsystem: (a) One-parameter bifurcation with$ [{\rm{I}}{{\rm{P}}_3}] = 1, 1.05, 1.1, 1.2 \; {\rm{{\text{μ}} mol/L}} $ , respectively. (b) Two-parameter bifurcation with $[{{\rm{IP}} _3}]$ and ${L_{{\rm{I}}{{\rm{P}}_3}}}$ as parameters. The bifurcation curves represent fold bifurcation ($f$ ), Hopf bifurcation (hb ) and the saddle-node on invariant circle bifurcation ($snic$ ) of equilibrium points. (c) Enlargement part of panel (b).$[{{\rm{IP}} _3}]$的取值/–1 ) ${\rm{SNIC}}$ ${\rm{Hopf}}$ 稳定极限环所在 $1 $ $0.2789$ $20.8584$ $[0.2789, 20.8584]$ $1.05$ $0.2239$ $19.3199$ $[0.2239, 19.3199]$ $1.1 $ $0.1842$ $17.6358$ $[0.1842, 17.6358]$ $1.2$ $0.1317$ $13.9694$ $[0.1317, 13.9694]$
表1 不同$[{{\rm{IP}} _3}]$ 下树突子系统中关键点的${L_{{\rm{I}}{{\rm{P}}_3}}}$ 的值Table1. Values of ${L_{{\rm{I}}{{\rm{P}}_3}}}$ of the key points at different $[{{\rm{IP}} _3}]$ values.3.2.耦合神经元反相簇放电 -->3.2.耦合神经元反相簇放电 33.2.1.参数${L_{{\rm{I}}{{\rm{P}}_3}}}$ ![]()
![]()
-->3.2.1.参数${L_{{\rm{I}}{{\rm{P}}_3}}}$ 对反相簇放电模式的影响 本文主要研究反相簇放电的模式及其转迁机制. 本节将$[{\rm{I}}{{\rm{P}}_3}]$ 固定在1.2 μmol/L, 研究参数${L_{{\rm{I}}{{\rm{P}}_3}}}$ 对系统动力学的影响. 随着${L_{{\rm{I}}{{\rm{P}}_3}}}$ 的增加, 反相簇放电表现为复杂的放电模式, 如图3 所示. 图中的黑色曲线是膜电位${V_1}$ (或${V_2}$ )随$t$ 的变化趋势, 红色曲线为钙离子浓度${[{\rm{Ca}} ]_1}$ (或${[{\rm{Ca}} ]_2}$ )随$t$ 的变化趋势, 蓝色曲线为${h_1}\;({h_2}$ )随$t$ 的变化趋势. 其中, ${L_{{\rm{I}}{{\rm{P}}_3}}}$ 分别对应树突子系统(方程(5 )和方程(6 ))具有稳定不动点的范围$({L_{{\rm{I}}{{\rm{P}}_3}}} < 0.1317 \;{\rm{PL}} \cdot {{\rm{s}}^{ - 1}})$ , 如图3(a) 和图3(b) 所示; 有稳定极限环的范围$(0.1317 \! < \! {L_{{\rm{I}}{{\rm{P}}_3}}}\! < $ $ 13.9694\; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}})$ 如图3(c) —(e) 所示; 具有稳定焦点的范围$({L_{{\rm{I}}{{\rm{P}}_3}}} > 13.9694 \;{\rm{PL}} \cdot {{\rm{s}}^{ - 1}})$ 如图3(f) 所示.图 3 反相簇放电模式 (a) $ {L}_{{\rm{IP}}_{3}}=0\;{\rm{PL}}\cdot {\rm{s}}^{-1}$ ; (b) ${L_{{\rm{I}}{{\rm{P}}_3}}} = 0.1\; {\rm{PL}} \cdot {{\rm{s}}^{{\rm{ - 1}}}}$ ; (c) ${L_{{\rm{I}}{{\rm{P}}_3}}} = 0.4\; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ ; (d) $ {L}_{{\rm{IP}}_{3}}=0.5\;{\rm{PL}}\cdot {\rm{s}}^{\rm{-1}}$ ; (e) ${L}_{{\rm{IP}}_{3}}= $ $ 3\;{\rm{PL}}\cdot {\rm{s}}^{-1}$ ; (f) ${L_{{\rm{I}}{{\rm{P}}_3}}} = 15\; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ Figure3. Anti-phase bursting of the system: (a) $ {L}_{{\rm{IP}}_{3}}=0\;{\rm{PL}}\cdot {\rm{s}}^{\rm{-1}}$ ; (b) ${L_{{\rm{I}}{{\rm{P}}_3}}} = 0.1\; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ ; (c) ${L_{{\rm{I}}{{\rm{P}}_3}}} = 0.4 \;{\rm{PL}} \cdot {{\rm{s}}^{{\rm{ - 1}}}}$ ; (d) ${L}_{{\rm{IP}}_{3}}= $ $ 0.5\;{\rm{PL}}\cdot {\rm{s}}^{\rm{-1}}$ ; (e) $ {L}_{{\rm{IP}}_{3}}=3\;{\rm{PL}}\cdot {\rm{s}}^{\rm{-1}}$ ; (f) ${L_{{\rm{I}}{{\rm{P}}_3}}} = 15\; {\rm{PL}} \cdot {{\rm{s}}^{{\rm{ - 1}}}}$ .图3(c) 所示, 第一部分从初始时刻到三角形(▲), 对应${[{\rm{Ca}} ]_1}$ 处于稳定态; 另一部分从三角形(▲)到五角星(★), 对应${[{\rm{Ca}} ]_1}$ 处于振荡态. 根据${[{\rm{Ca}} ]_1}$ 振荡的动力学特性, 混合簇放电包括胞体簇放电和树突簇放电[20 ] . 图中圆点(●)代表初始时刻, 即$t = 0$ 的时刻, 菱形(◆)和正方形(■)分别代表该组参数值下胞体簇放电由静息态跃迁至放电态和由放电态跃迁至静息态的关键点. 参数${L_{{\rm{I}}{{\rm{P}}_3}}}$ 变化时对应的峰峰间期${\rm{ISI}}$ 序列如图4 所示. 图中两条红色直线对应${L_{{\rm{I}}{{\rm{P}}_3}}} = 0.131 \; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ 和${L_{{\rm{I}}{{\rm{P}}_3}}} = 13.9694\; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ , 分别表示钙浓度${[{\rm{Ca}} ]_1}$ 由静息到振荡, 以及由振荡到静息的临界值. 当${L_{{\rm{I}}{{\rm{P}}_3}}} < 0.131 \;{\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ 或${L_{{\rm{I}}{{\rm{P}}_3}}} > 13.9694\;{\rm{PL}} \cdot {{\rm{s}}^{{\rm{ - 1}}}}$ 时, 随着${L_{{\rm{I}}{{\rm{P}}_3}}}$ 的改变两个连续簇放电之间的时间间隔几乎不再变化, 如图4(c) 和图4(d) 所示.图 4 (a) ${L_{{\rm{I}}{{\rm{P}}_3}}}$ 变化时膜电位$ {V}_{1} $ 对应的${\rm{ISI}}$ 分岔序列; (b) 纵坐标${\rm{ISI}}$ 取对数对应的分岔图; (c), (d) 图(a)的部分放大图Figure4. (a) ${\rm{ISI}}$ diagram of membrane potential ${V_1}$ with parameter ${L_{{\rm{I}}{{\rm{P}}_3}}}$ ; (b) logarithm as ordinate of ${\rm{ISI}}$ for panel (a); (c), (d) partially enlarged diagram of panel (a).3.2.2.胞体簇放电模式的单参数分岔分析 -->3.2.2.胞体簇放电模式的单参数分岔分析 部分有代表性的胞体簇放电(绿色曲线)的快慢分岔分析如图5 所示, 本文只分析膜电位${V_1}$ 的放电模式. 当${L_{{\rm{I}}{{\rm{P}}_3}}} = 0\; {\rm{PL}} \cdot {{\rm{s}}^{{\rm{ - 1}}}}$ 时, 系统的反相簇放电的分岔分析如图5(a) 和图5(b) 所示(分别对应图3(a) 中第1个和第4个簇放电), 此时${[{\rm{Ca}} ]_1}$ 为定值($ {[{\rm{Ca}} ]_1} = 0$ μmol/L). 如图5(a) 所示, 系统轨线的下状态即静息态经由鞍结分岔(${{\rm{F}}_1}$ )跃迁至上状态, 由于稳定焦点的吸引, 轨线的振幅逐渐减少并逐渐回到稳定焦点. 但轨线经过${\rm{sub}}{{\rm{H}}_1}$ 分岔时, 稳定焦点变为不稳定, 同时稳定极限环产生. 故受稳定极限环的吸引和不稳定焦点的排斥作用, 轨线的振幅逐渐增大并接近稳定极限环, 进入快速峰放电状态. 最终, 放电态经由极限环的鞍结分岔(${\rm{LP}}{{\rm{C}}_2}$ )转迁至下支的稳定结点, 回到静息态. 因此, 这种簇放电模式为经由“fold/fold limit cycle”滞后环的“subHopf/fold limit cycle”型簇放电. 在图5(b) 中, 系统的轨线通过鞍结分岔(${{\rm{F}}_1}$ )由静息态跃迁至放电态, 又经极限环上的鞍结分岔(${\rm{LP}}{{\rm{C}}_2}$ )跃迁至下状态的静息态. 因此, 称其为“fold/fold limit cycle”型簇放电.图 5 反相簇放电的快慢分岔分析 (a), (b) ${L_{{\rm{I}}{{\rm{P}}_3}}} = 0 \;{\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ ; (c), (d) ${L_{{\rm{I}}{{\rm{P}}_3}}} = 0.1 \;{\rm{PL}} \cdot {{\rm{s}}^{{\rm{ - 1}}}}$ ; (e) ${L_{{\rm{I}}{{\rm{P}}_3}}} = 0.4 \;{\rm{PL}} \cdot {{\rm{s}}^{{\rm{ - 1}}}}$ ; (f) ${L_{{\rm{I}}{{\rm{P}}_3}}} = $ $ 0.5 \;{\rm{PL}} \cdot {{\rm{s}}^{{\rm{ - 1}}}}$ Figure5. Fast-slow bifurcation analysis of anti-phase bursting with (a), (b) ${L_{{\rm{I}}{{\rm{P}}_3}}} = 0\; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ ; (c), (d) ${L_{{\rm{I}}{{\rm{P}}_3}}} = 0.1\; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ ; (e) ${L_{{\rm{I}}{{\rm{P}}_3}}} = $ $ 0.4 \;{\rm{PL}} \cdot {{\rm{s}}^{{\rm{ - 1}}}}$ ; (f) ${L_{{\rm{I}}{{\rm{P}}_3}}} = 0.5 \;{\rm{PL}} \cdot {{\rm{s}}^{{\rm{ - 1}}}}$ .${L_{{\rm{I}}{{\rm{P}}_3}}}$ 增加到$0.1\; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ 时, ${[{\rm{Ca}} ]_1}$ 仍为定值($ {[{\rm{Ca}} ]_1} = 0.0119 \; {\rm{{\text{μ}} mol/L}} $ ). 如图5(c) 和图5(d) 所示(分别对应图3(b) 中第1和第2个簇放电), 分别为“fold/fold limit cycle”型簇放电和经由“fold/fold limit cycle” 滞后环的“subHopf/fold limit cycle”型簇放电(分别与图5(b) 和图5(a) 的簇放电模式相同).${L_{{\rm{I}}{{\rm{P}}_3}}} = 0.4 \;{\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ 和${L_{{\rm{I}}{{\rm{P}}_3}}} = 0.5\; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ 时, 如图5(e) 以及图5(f) 所示(分别对应图3(c) 以及图3(d) 中的第1个簇放电), 簇放电的模式都是经由“fold/fold limit cycle”滞后环的“subHopf/fold limit cycle”型簇放电(都与图5(a) 相同).3.2.3.树突簇放电模式的双参数分岔分析 -->3.2.3.树突簇放电模式的双参数分岔分析 当${L_{{\rm{I}}{{\rm{P}}_3}}}$ 变化时, 系统表现为不同类型的复杂簇放电模式. 而复杂模式簇放电是一种至少包含两种时间尺度的不规则簇放电现象[19 ] , 可能会有多种因素对它的放电模式产生影响, 比如分岔的类型、分岔曲线的相对位置等. 下面通过双参数分岔来研究${L_{{\rm{I}}{{\rm{P}}_3}}}$ 的变化对快子系统分岔结构的影响, 并进一步揭示复杂簇放电模式产生的动力学机制. 据快慢动力学分析选择慢变量${h_i}$ 和${\rm {[Ca]}}_{i} $ 作为分岔参数. 这里添加一个辅助变量${g_{{\rm{CANTo}}{{\rm{t}}_i}}} = {g_{{\rm{CAN}}}}f({\rm {[Ca]}}_{i} )$ , 代替${\rm {[Ca]}}_{i}$ 作为分岔参数, $f({\rm {[Ca]}}_{i} )$ 是一个单调递增的下凹函数(见附录A ). 双参数分岔如图6 所示, 红色曲线表示平衡点的鞍结分岔f , 蓝色曲线表示 Hopf 分岔$hb$ (与${\rm{sub}}{{\rm{H}}_1}$ 对应), 绿色曲线表示极限环的鞍结分岔${lpc}_{\rm{1}}{\rm{, }}\;{lpc}_{\rm{2}}$ (分别对应于${\rm{LP}}{{\rm{C}}_1}$ , ${\rm{LP}}{{\rm{C}}_2}$ 两个极限环的鞍结分岔点), 黑色曲线表示系统轨迹在(${h_1}, \; {g_{{\rm{CA}}{{\rm{N}}_{{\rm{Tot}}1}}}}$ )平面上的投影.图 6 ${L_{{\rm{I}}{{\rm{P}}_3}}} $ 取不同值时反相簇放电的双参数$({h_1}, \; {g_{{\rm{CA}}{{\rm{N}}_{{\rm{Tot}}1}}}})$ 分岔分析 (a) ${L_{{\rm{I}}{{\rm{P}}_3}}} = $ $ 0.4\; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ ; (b) ${L_{{\rm{I}}{{\rm{P}}_3}}} = 0.5 \;{\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ ; (c) ${L_{{\rm{I}}{{\rm{P}}_3}}} = 3 \;{\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ Figure6. Two-parameter bifurcation of the anti-phase bursting in $({h_1}, \; {g_{{\rm{CA}}{{\rm{N}}_{{\rm{Tot}}1}}}})$ -plane with different $ {L_{{\rm{I}}{{\rm{P}}_3}}} $ : (a) ${L_{{\rm{I}}{{\rm{P}}_3}}} = $ $ 0.4\; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ ; (b) ${L_{{\rm{I}}{{\rm{P}}_3}}} = 0.5\; {\rm{PL}} \cdot {{\rm{s}}^{{\rm{ - 1}}}}$ ; (c) ${L_{{\rm{I}}{{\rm{P}}_3}}} = 3\; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ .$ {L_{{\rm{I}}{{\rm{P}}_3}}} = 0.4\; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}} $ 时, 双参数分岔如图6(a) 所示. 图中的标记分别与图3(c) 中的相对应, 即圆点(●)代表初始时刻, 菱形(◆)和正方形(■)分别代表该组参数值下胞体簇放电的静息态和放电态两者转迁的关键点, 即对应图5(e) 中的鞍结分岔${{\rm{F}}_1}$ 和极限环上的鞍结分岔${\rm{LP}}{{\rm{C}}_2}$ 两个重要的分岔点. 混合簇放电中, 从$t = 0$ , 即圆点(●)开始, 穿过平衡点的鞍结分岔曲线$f$ 和极限环的鞍结分岔曲线$lpc_{\rm 2}$ , 完成第一个簇放电即胞体簇放电. 之后随着${g_{{\rm{CA}}{{\rm{N}}_{{\rm{Tot}}1}}}}$ 向右跳跃的增加, 混合簇放电的胞体簇放电转迁为树突簇放电, 最终完成一个混合簇放电周期.${L_{{\rm{I}}{{\rm{P}}_3}}}$ 增加至$0.5\; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ 时, 如图6(b) 所示, 此时系统的轨迹为周期2, 对应图3(d) 两组不同的混合簇放电(每组包括1个胞体簇放电和1个树突簇放电). 当${L_{{\rm{I}}{{\rm{P}}_3}}}$ 继续增加至$3 \;{\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ , 如图6(c) 所示, 此时, 系统轨线在${g_{{\rm{CA}}{{\rm{N}}_{{\rm{Tot}}1}}}}$ 向右跳跃, 即突增之前没有在平衡点的鞍结分岔曲线f 和极限环的鞍结分岔曲线$ lpc_{\rm 2} $ 之间转迁. 故系统不存在胞体簇放电, 而只存在树突簇放电.3.2.4.空间中胞体簇放电的单参数分岔分析 -->3.2.4.空间中胞体簇放电的单参数分岔分析 为了更清楚地了解簇放电的运行轨迹, 下面研究三维空间中${L_{{\rm{I}}{{\rm{P}}_3}}} = 0\; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ 时系统的胞体簇放电(图3(a) 中第一个簇放电). $({h_1}, {n_1}, {V_1})$ 空间中单参数分岔分析如图7(a) 所示, 其中绿色和蓝色曲线分别为稳定和不稳定极限环. 浅黑色、浅蓝色和浅绿色等曲线分别为黑色、蓝色和绿色等同色号的曲线在(${h_1}, {V_1}$ )平面上的投影(如浅蓝色和浅绿色曲线分别为稳定(绿色)和不稳定(蓝色)极限环在(${h_1}, {V_1}$ )平面上的投影). 轨迹运动的顺序为红色 → 橙色 → 紫色.图 7 (a) $ {L}_{{\rm{IP}}_{3}}=0\;{\rm{PL}}\cdot {\rm{s}}^{-1}$ 时反相簇放电的快子系统在$({h_1}, {n_1}, {V_1})$ 空间中的分岔结构图; (b) 图(a)的部分放大Figure7. (a) Bifurcation structure of the fast subsystem of anti-phase bursting in $({h_1}, {n_1}, {V_1})$ space with ${L}_{{\rm{IP}}_{3}}= $ $ 0\;{\rm{PL}}\cdot {\rm{s}}^{-1}$ ; (b) partial enlargement of panel (a).${h_1} = 0.2671$ 时, 第一个亚临界Hopf分岔产生, 当${h_1} = 0.2816$ 时发生了稳定和不稳定极限环相遇的流分岔(TR), 故当$0.2671\leqslant {h}_{1}\leqslant 0.2816$ 时, 稳定焦点、稳定和不稳定的极限环共存. 当${h_1} = $ $ 0.279$ 时, 稳定焦点、稳定和不稳定极限环相应的在$({n_1}, {s_1}, {V_1})$ 空间的相图如图8(a) 所示, 轨迹都是呈顺时针旋转. 在$({n_1}, {V_1})$ 平面上的投影如图8(b) 所示. 由于稳定平衡点—稳定焦点(红色点)、不稳定(蓝色曲线)和稳定极限环(橄榄绿色曲线)共存, 使得系统的轨线表现为复杂的混合簇放电模式.图 8 ${h_1} = 0.279$ 时, 系统的稳定焦点、稳定和不稳定的极限环 (a) $({n_1}, {s_1}, {V_1})$ 空间; (b) $({n_1}, {V_1})$ 平面Figure8. Stable focus, stable and unstable limit cycle in (a) $({n_1}, {s_1}, {V_1})$ -space, (b) $({n_1}, {V_1})$ -plane.4.结 论 前包钦格复合体中兴奋性吸气神经元的节律活动与呼吸节律密切相关. 呼吸节律受离子电流, 特别是${I_{{\rm{NaP}}}}$ 和${I_{{\rm{CAN}}}}$ 的影响, 并且还受到与${\rm{C}}{{\rm{a}}^{2 + }}$ 相关的$[{\rm{I}}{{\rm{P}}_3}]$ 和${L_{{\rm{I}}{{\rm{P}}_3}}}$ 的影响. 当$[{\rm{I}}{{\rm{P}}_3}]$ 处于一定范围时, 钙离子浓度发生周期性波动, 导致系统中各个变量的快慢尺度发生相应变化, 从而导致了混合簇放电的产生[14 ] . 以往的大部分研究表明, 复杂模式簇放电是由钙的周期性波动引起的[19 ,20 ] .${L_{{\rm{I}}{{\rm{P}}_3}}}$ 的变化对反相簇放电特别是混合簇放电的影响, 并从动力学角度分析了混合簇放电产生的机理. 结果表明, 当${L_{{\rm{I}}{{\rm{P}}_3}}}$ 增加时, 耦合神经元的反相簇放电分别表现为钙离子浓度处于稳态时的混合簇放电、钙离子浓度处于波动时的簇放电以及常规簇放电等模式. 本文的研究表明${L_{{\rm{I}}{{\rm{P}}_3}}}$ 的变化对耦合前包钦格复合体神经元放电节律有重要的影响, 钙离子浓度为定值时也可以促进混合簇放电的产生(如图3(a) 和图3(b) 所示). 故钙离子浓度的周期性波动不是混合簇放电产生的必要条件. 本文的研究结果对于了解前包钦格复合体网络的动态节律具有重要意义, 有助于了解正常呼吸节律的产生机制, 为研究病态呼吸节律的产生机理提供一定的理论支持, 并为将来实验和临床中对病态呼吸节律的控制提供一定的理论依据. 本文的研究方法也可以推广并应用于其他具有快慢时间尺度的神经系统的研究中.附录A.模型中涉及的函数 方程(1 )—(6 )中变量的表达式及参数值如(A1 )式—(A5 )式和表A1 所示.参数 参数值 参数 参数值 参数 参数值 ${C_{\rm{m}}}$ $ 21 \; {\rm{ {\text{μ} } F} }$ ${g_{{\rm{Na}}}}$ $28 \; {\rm{nS}}$ ${L_{{\rm{I}}{{\rm{P}}_3}}}$ ${\rm{varied, } } \;{\rm{PL} } \cdot { {\rm{s} }^{ - 1} }$ ${E_{{\rm{Na}}}}$ $50 \; {\rm{mV}}$ ${g_{\rm{K}}}$ $11.2 \; {\rm{nS}}$ ${P_{{\rm{I}}{{\rm{P}}_3}}}$ $31000 \; {\rm{PL}} \cdot {{\rm{s}}^{ - 1}}$ ${E_{\rm{K}}}$ $ - 85 \; {\rm{mV}}$ ${g_{{\rm{Nap}}}}$ $15 \; {\rm{nS}}$ ${K_{\rm{l}}}$ 1.0 μmol/L ${E_{\rm{L}}}$ $ - 58 \; {\rm{mV}}$ ${g_{ {\rm{tonic \text- e} } } }$ $0.4 \; {\rm{nS}}$ ${K_{\rm{a}}}$ 0.4 μmol/L ${E_{ {\rm{syn \text- e} } } }$ $0 \; {\rm{mV}}$ ${g_{{\rm{CAN}}}}$ $0.7 \; {\rm{nS}}$ ${V_{{\rm{SERCA}}}}$ $400 \; {\rm{aMol}} \cdot {{\rm{S}}^{ - 1}}$ ${\theta _m}$ $ - 34 \; {\rm{mV}}$ ${g_{ {\rm{syn \text- e} } } }$ $9 \; {\rm{nS}}$ ${E_{{\rm{SERCA}}}}$ 0.2 μmol/L ${\theta _n}$ $ - 29 \; {\rm{mV}}$ ${g_{\rm{L}}}$ $11.2 \; {\rm{nS}}$ $A$ $0.001 \; {({\rm{ {\text{μ} } mol} }/{\rm{L)} }^{ - 1} } \cdot {\rm{m} }{ {\rm{s} }^{ - 1} }$ ${\theta _{mp}}$ $ - 40\; {\rm{mV}}$ ${\sigma _{\rm{s}}}$ $ - 5 \; {\rm{mV}}$ ${K_{{\rm{d}}}}$ 0.4 μmol/L ${\theta _h}$ $ - 48 \; {\rm{mV}}$ ${\sigma _n}$ $ - 4 \;{\rm{mV}}$ ${K_{{\rm{CAN}}}}$ 0.74 μmol/L ${\alpha _{\rm{s}}}$ 0.2 ms-1 ${\sigma _{mp}}$ $ - 6 \; {\rm{mV}}$ ${n_{{\rm{CAN}}}}$ $0.97$ ${\bar \tau _{\rm{s}}}$ 5 ms ${\sigma _h}$ $5 \; {\rm{mV}}$ $[{\rm{I}}{{\rm{P}}_3}]$ varied, μmol/L ${\bar \tau _h}/\varepsilon $ 10000 ms ${\theta _s}$ $ - 10 \; {\rm{mV}}$ ${[{\rm{Ca}}]_{{\rm{Tot}}}}$ 1.25 μmol/L ${\bar \tau _n}$ 10 ms ${\sigma _m}$ $ - 5 \; {\rm{mV}}$ ${f_m}$ $0.000025 \; {\rm{P}}{{\rm{L}}^{ - 1}}$ $\sigma $ $0.185$
表A1 理论模型中的参数值TableA1. Parameter values used in the theoretical model 


















































 图 1 参数
图 1 参数



































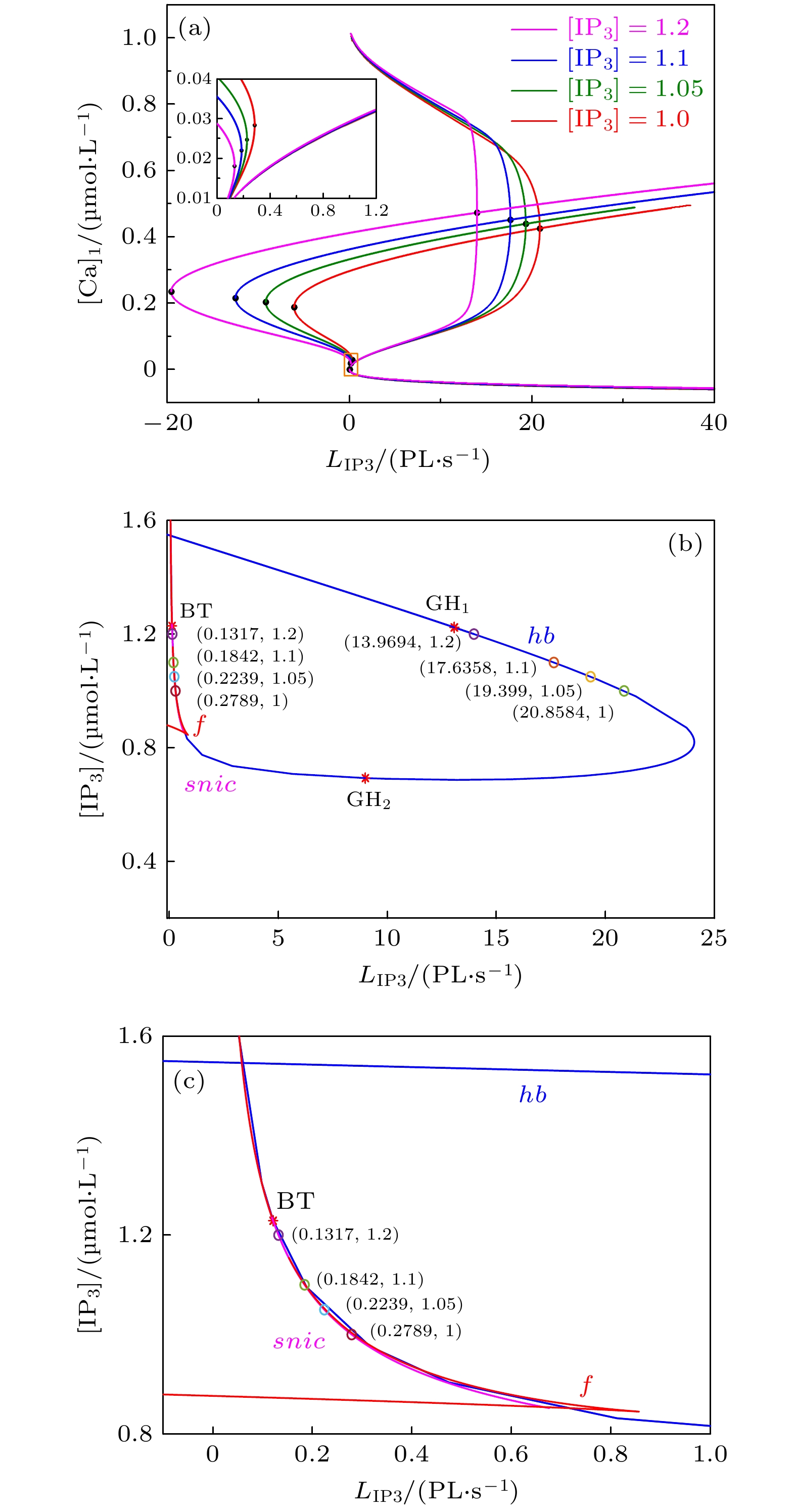 图 2 树突子系统 (方程(5)和方程(6))的动力学分析 (a) 单参数分岔, 其中
图 2 树突子系统 (方程(5)和方程(6))的动力学分析 (a) 单参数分岔, 其中






























 图 3 反相簇放电模式 (a)
图 3 反相簇放电模式 (a) 

























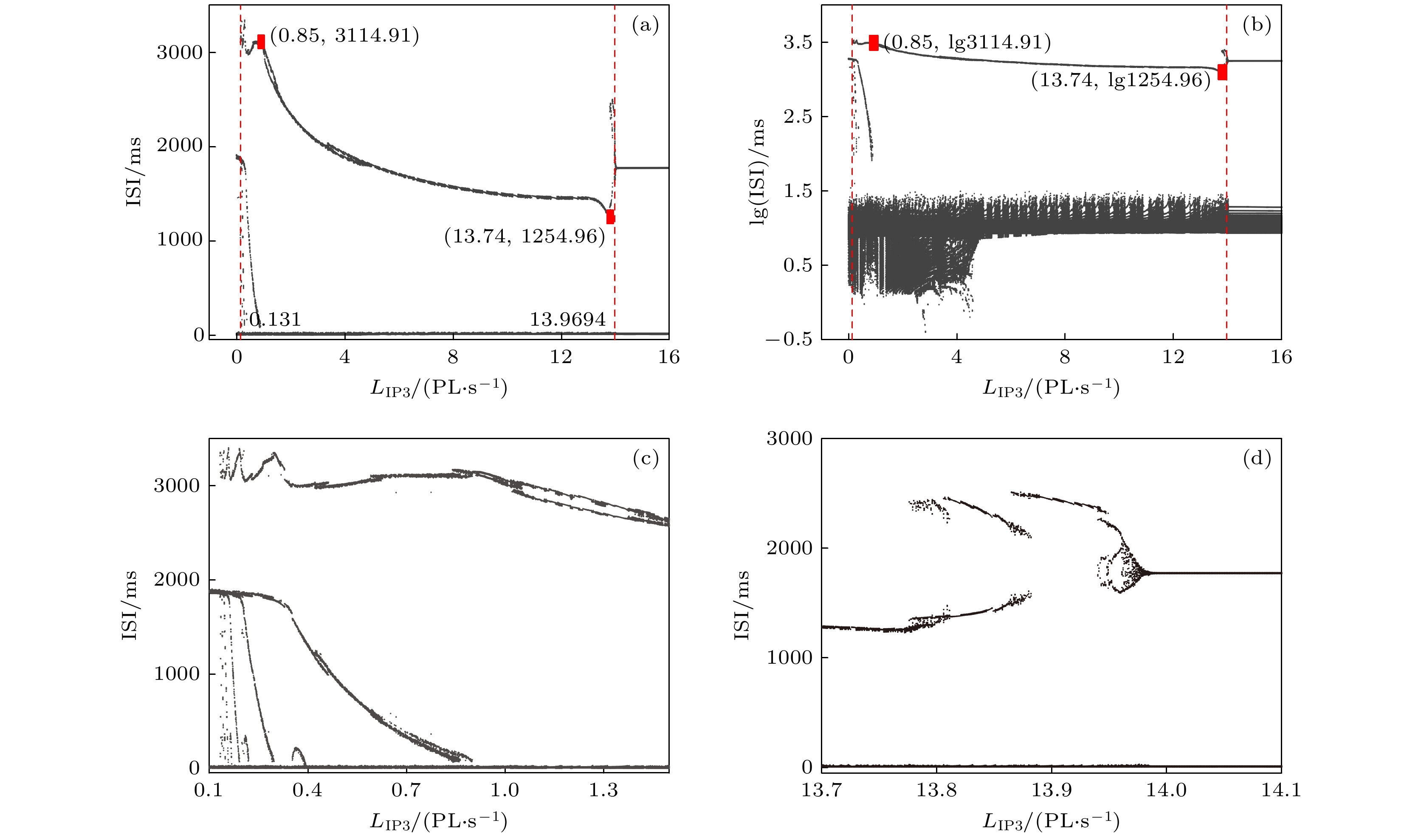 图 4 (a)
图 4 (a) 
















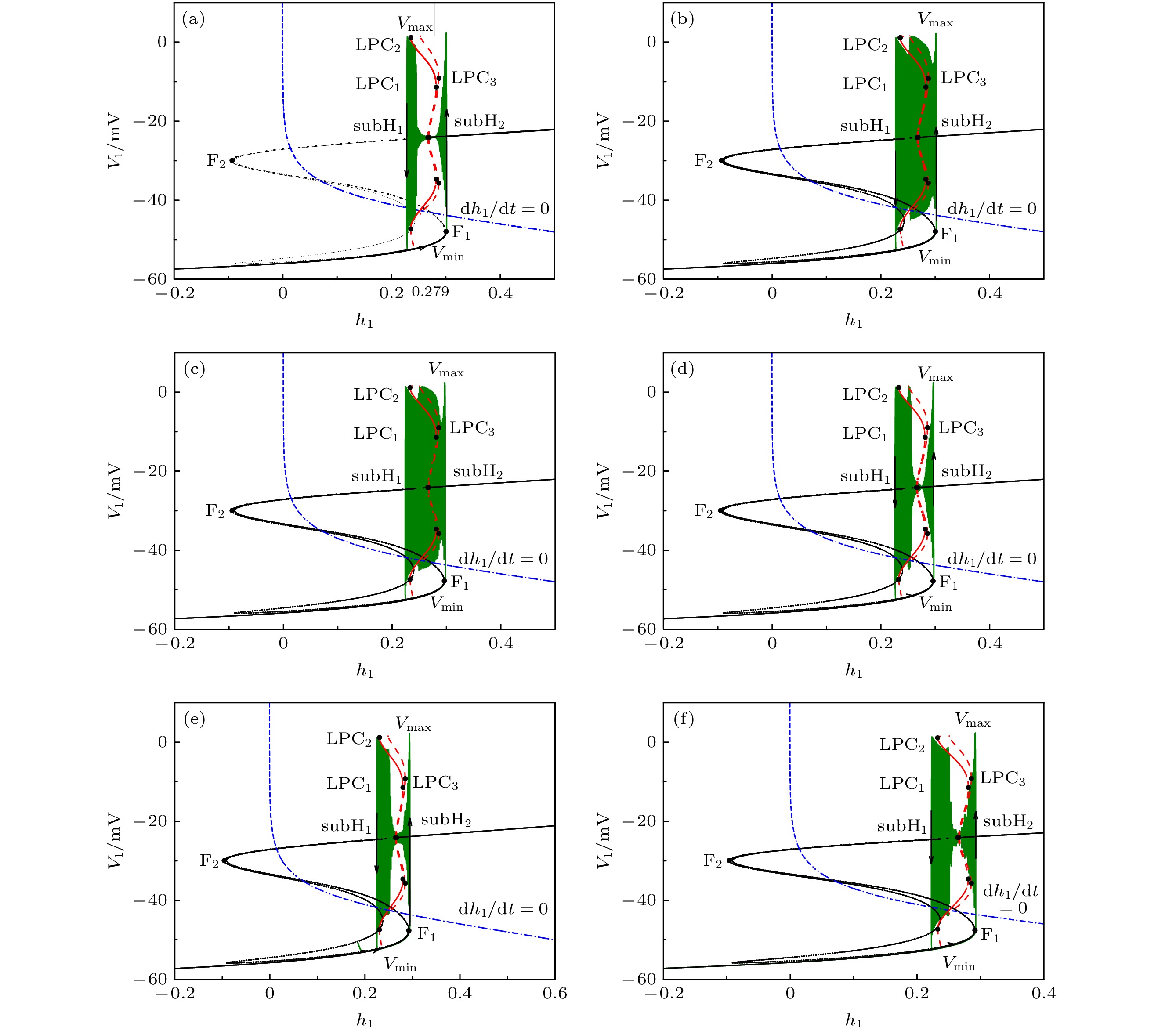 图 5 反相簇放电的快慢分岔分析 (a), (b)
图 5 反相簇放电的快慢分岔分析 (a), (b) 




























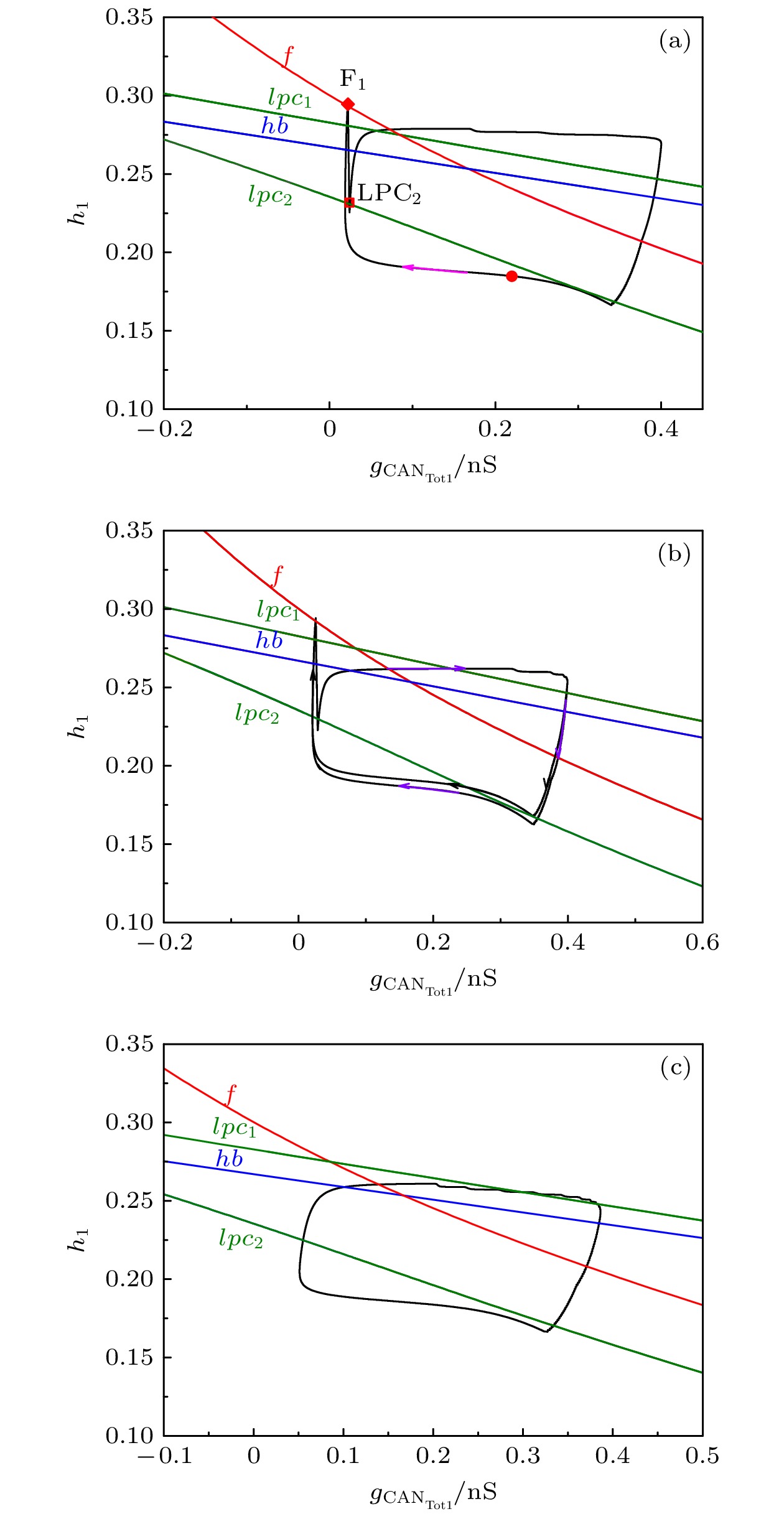 图 6
图 6 




























 图 7 (a)
图 7 (a) 











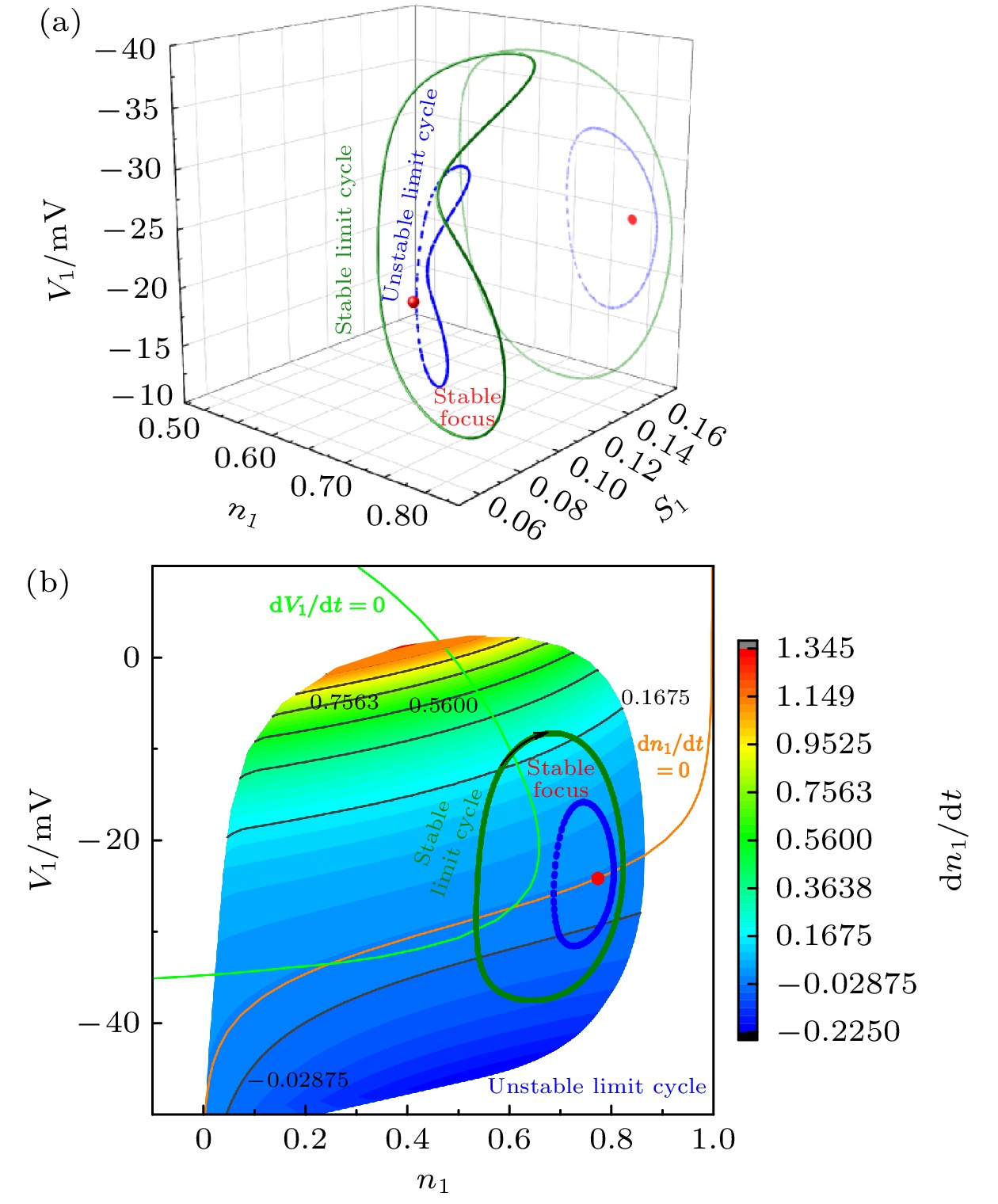 图 8
图 8 













