Application of genetic modification technologies in molecular design breeding of sheep
Haitao Wang,1,2, Tingting Li3, Xun Huang1,2, Runlin Ma,1,2,4, Qiuyue Liu,11. Innovation Academy for Seed Design, Chinese Academy of Sciences, Beijing 100101, China 2. State Key Laboratory of Molecular Developmental Biology, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing 100101, China 3. Zhejiang A&F University, Hangzhou 311300, China; 4. University of Chinese Academy of Sciences, Beijing 100049, China
Abstract Genetic modification technologies can be used for modifying animal genome to express exogenous genes or affect the function of endogenous genes. In animal breeding, genetic modification technologies allow the rapid generation of germplasms with beneficial traits. It includes traditional genetic modification, virus or sperm carrier-mediated genetic modification and nuclease-mediated genome editing, especially the CRISPR/Cas9, one of the artificial nuclease genome editing technologies, have been applied in genome editing in many domestic animals including sheep (Ovis aries). Compared with conventional strategies used for animal breeding, there is great value for sheep breeding improvement by using genome editing technology, which is more effective and timesaving. In this review, we summarize the approaches of genetic modification in sheep and discuss the possibility of molecular design and breeding of sheep by genome editing technologies. We also identify the potential bottlenecks and challenges of these technologies in sheep breeding. Keywords:sheep;genetic modification;genome editing;animal breeding
PDF (1612KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 王海涛, 李亭亭, 黄勋, 马润林, 刘秋月. 遗传修饰技术在绵羊分子设计育种中的应用. 遗传[J], 2021, 43(6): 580-600 doi:10.16288/j.yczz.21-087 Haitao Wang. Application of genetic modification technologies in molecular design breeding of sheep. Hereditas(Beijing)[J], 2021, 43(6): 580-600 doi:10.16288/j.yczz.21-087
通过对性状相关的基因进行修饰,在绵羊中可以实现多个方向的应用,例如生产性状,包括肉产量、肉质、生长速度等;毛纤维性状,包括羊毛产量、结构、毛色等;繁殖性状,包括产羔数等;高抗病能力;制备人类疾病模型;通过乳腺反应器生产药物等。 Fig. 4Applications of molecular design breeding in sheep using genetic modification
ZhaoJG, LaiLX, JiWZ, ZhouQ. Genome editing in large animals: current status and future prospects Natl Sci Rev, 2019,6(3):402-420. [本文引用: 1]
IiizumiS, KurosawaA, SoS, IshiiY, ChikaraishiY, IshiiA, KoyamaH, AdachiN. Impact of non-homologous end-joining deficiency on random and targeted DNA integration: implications for gene targeting Nucleic Acids Res, 2008,36(19):6333-6342. [本文引用: 1]
ChoulikaA, PerrinA, DujonB, NicolasJF. Induction of homologous recombination in mammalian chromosomes by using the I-SceI system of Saccharomyces cerevisiae Mol Cell Biol, 1995,15(4):1968-1973. [本文引用: 1]
GordonJW, ScangosGA, PlotkinDJ, BarbosaJA, RuddleFH. Genetic transformation of mouse embryos by microinjection of purified DNA Proc Natl Acad Sci USA, 1980,77(12):7380-7384.
HammerRE, PurselVG, RexroadCE, WallRJ, BoltDJ, EbertKM, PalmiterRD, BrinsterRL. Production of transgenic rabbits, sheep and pigs by microinjection Nature,1985,315(6021):680-683. [本文引用: 5]
IshinoY, ShinagawaH, MakinoK, AmemuraM, NakataA. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product J Bacteriol, 1987,169(12):5429-5433. [本文引用: 1]
LeeMS, GippertGP, SomanKV, CaseDA, WrightPE. Three-dimensional solution structure of a single zinc finger DNA-binding domain Science, 1989,245(4918):635-637. [本文引用: 1]
CampbellKH, McWhirJ, RitchieWA, WilmutI. Sheep cloned by nuclear transfer from a cultured cell line Nature, 1996,380(6569):64-66. [本文引用: 1]
SchniekeAE, KindAJ, RitchieWA, MycockK, ScottAR, RitchieM, WilmutI, ColmanA, CampbellKH. Human factor IX transgenic sheep produced by transfer of nuclei from transfected fetal fibroblasts Science, 1997,278(5346):2130-2133. [本文引用: 3]
MoscouMJ, BogdanoveAJ. A simple cipher governs DNA recognition by TAL effectors Science, 2009,326(5959):1501. [本文引用: 1]
RémyS, TessonL, MénoretS, UsalC, ScharenbergAM, AnegonI. Zinc-finger nucleases: a powerful tool for genetic engineering of animals Transgenic Res, 2010,19(3):363-371. [本文引用: 1]
CrispoM, MuletAP, TessonL, BarreraN, CuadroF, dosSantos-Neto PC, NguyenTH, CrénéguyA, BrusselleL, AnegónI, MenchacaA. Efficient generation of myostatin knock-out sheep using CRISPR/Cas9 technology and microinjection into zygotes PLoS One, 2015,10(8):e0136690. [本文引用: 4]
MurrayJD, NancarrowCD, MarshallJT, HazeltonIG, WardKA. Production of transgenic merino sheep by microinjection of ovine metallothionein-ovine growth hormone fusion genes Reprod Fertil Dev, 1989,1(2):147-155. [本文引用: 4]
AdamsNR, BriegelJR, WardKA. The impact of a transgene for ovine growth hormone on the performance of two breeds of sheep J Anim Sci, 2002,80(9):2325-2333. [本文引用: 4]
ClementsJE, WallRJ, NarayanO, HauerD, SchoborgR, ShefferD, PowellA, CarruthLM, ZinkMC, RexroadCE. Development of transgenic sheep that express the visna virus envelope gene Virology, 1994,200(2):370-380. [本文引用: 4]
RexroadCE, HammerRE, BoltDJ, MayoKE, FrohmanLA, PalmiterRD, BrinsterRL. Production of transgenic sheep with growth-regulating genes Mol Reprod Dev, 1989,1(3):164-169. [本文引用: 4]
RexroadCE, MayoK, BoltDJ, ElsasserTH, MillerKF, BehringerRR, PalmiterRD, BrinsterRL. Transferrin- and albumin-directed expression of growth-related peptides in transgenic sheep J Anim Sci, 1991,69(7):2995-3004. [本文引用: 3]
WrightG, CarverA, CottomD, ReevesD, ScottA, SimonsP, WilmutI, GarnerI, ColmanA. High level expression of active human alpha-1-antitrypsin in the milk of transgenic sheep Biotechnology (NY), 1991,9(9):830-834. [本文引用: 3]
ClarkAJ, BessosH, BishopJO, BrownP, HarrisS, LatheR, McClenaghanM, ProwseC, SimonsJP, WhitelawCBA, WilmutI. Expression of human anti-hemophilic factor IX in the milk of transgenic sheep Bio Tech, 1989,7:487-492. [本文引用: 2]
NiemannH, HalterR, CarnwathJW, HerrmannD, LemmeE, PaulD. Expression of human blood clotting factor VIII in the mammary gland of transgenic sheep Transgenic Res, 1999,8(3):237-247. [本文引用: 3]
DamakS, JayNP, BarrellGK, BullockDW. Targeting gene expression to the wool follicle in transgenic sheep Biotechnology (NY), 1996,14(2):181-184. [本文引用: 4]
BawdenCS, PowellBC, WalkerSK, RogersGE. Expression of a wool intermediate filament keratin transgene in sheep fibre alters structure Transgenic Res, 1998,7(4):273-287. [本文引用: 2]
DengSL, YuK, WuQ, LiY, ZhangXS, ZhangBL, LiuGS, LiuYX, LianZX. Toll-like receptor 4 reduces oxidative injury via glutathione activity in sheep Oxid Med Cell Longev, 2016,2016:9151290. [本文引用: 1]
DenningC, BurlS, AinslieA, BrackenJ, DinnyesA, FletcherJ, KingT, RitchieM, RitchieWA, RolloM, deSousa P, TraversA, WilmutI, ClarkAJ. Deletion of the alpha(1,3)galactosyl transferase (GGTA1) gene and the prion protein (PrP) gene in sheep Nat Biotechnol, 2001,19(6):559-562. [本文引用: 4]
McCreathKJ, HowcroftJ, CampbellKH, ColmanA, SchniekeAE, KindAJ. Production of gene-targeted sheep by nuclear transfer from cultured somatic cells Nature, 2000,405(6790):1066-1069. [本文引用: 5]
DuanB, ChengL, GaoY, YinFX, SuGH, ShenQY, LiuK, HuX, LiuX, LiGP. Silencing of fat-1 transgene expression in sheep may result from hypermethylation of its driven cytomegalovirus (CMV) promoter Theriogenology, 2012,78(4):793-802. [本文引用: 3]
HeX, LiHT, ZhouZY, ZhaoZS, LiW. Production of brown/yellow patches in the SLC7A11 transgenic sheep via testicular injection of transgene J Genet Genomics, 2012,39(6):281-285. [本文引用: 4]
QinYR, ChenH, ZhangYN, ZhuCY, GaoB, YinYH, LiW, ShiQQ, ZhengMM, XuQ, SongJZ, LiBC. Cloning of the Xuhuai goat PPARγ gene and the preparation of transgenic sheep Biochem Genet, 2013,51(7-8):543-553. [本文引用: 4]
QinYR, ZhangYN, YinYH, XuF, GaoB, ShiQQ, ZhuCY, LiW, LiBC. Cloning of Xuhuai goat lipoprotein lipase gene and the preparation of transgenic sheep Mol Biol Rep, 2012,39(8):8439-8446. [本文引用: 4]
RitchieWA, KingT, NeilC, CarlisleAJ, LillicoS, McLachlanG, WhitelawCBA. Transgenic sheep designed for transplantation studies Mol Reprod Dev, 2009,76(1):61-64. [本文引用: 2]
BevacquaRJ, Fernandez-MartinR, CanelNG, GibbonsA, TexeiraD, LangeF, Vans LandschootG, SavyV, BriskiO, HiriartMI, GruesoE, IvicsZ, TabogaO, KuesWA, FerrarisS, SalamoneDF. Assessing Tn5 and Sleeping Beauty for transpositional transgenesis by cytoplasmic injection into bovine and ovine zygotes PLoS One, 2017,12(3):e0174025. [本文引用: 2]
HuSW, NiW, SaiWJF, ZhangH, CaoXD, QiaoJ, ShengJL, GuoF, ChenCF. Sleeping Beauty-mediated knockdown of sheep myostatin by RNA interference Biotechnol Lett, 2011,33(10):1949-1953. [本文引用: 6]
Rodriguez-SosaJR, SilvertownJD, FosterRA, MedinJA, HahnelA. Transduction and transplantation of spermatogonia into the testis of ram lambs through the extra-testicular rete Reprod Domest Anim, 2009,44(4):612-620. [本文引用: 2]
ZhangXM, WangLQ, WuYS, LiWR, AnJ, ZhangFC, LiuMJ. Knockout of myostatin by zinc-finger nuclease in sheep fibroblasts and embryos Asian-Australas J Anim Sci, 2016,29(10):1500-1507. [本文引用: 2]
ZhaoXX, NiW, ChenCF, SaiWJF, QiaoJ, ShengJL, ZhangH, LiGZ, WangDW, HuSW. Targeted editing of myostatin gene in sheep by transcription activator-like effector nucleases Asian-Australas J Anim Sci, 2016,29(3):413-418. [本文引用: 5]
LiHH, WangG, HaoZQ, ZhangGZ, QingYB, LiuSH, QingLL, PanWR, ChenL, LiuGC, ZhaoRP, JiaBY, ZengLY, GuoJX, ZhaoLX, ZhaoH, LvCX, XuKX, ChengWM, LiHS, ZhaoHY, WangW, WeiHJ. Generation of biallelic knock-out sheep via gene-editing and somatic cell nuclear transfer Sci Rep, 2016,6:33675. [本文引用: 5]
HanHB, MaYH, WangT, LianT, TianXZ, HuR, DengSL, LiKP, WangF, LiN, LiuGS, ZhaoYF, LianZX. One-step generation of myostatin gene knockout sheep via the CRISPR/Cas9 system Front Agr Sci & Eng,2014,1(1):2-5. [本文引用: 5]
WilliamsDK, PinzónC, HugginsS, PryorJH, FalckA, HermanF, OldeschulteJ, ChavezMB, FosterBL, WhiteSH, WesthusinME, SuvaLJ, LongCR, GaddyD. Genetic engineering a large animal model of human hypophosphatasia in sheep Sci Rep, 2018,8(1):16945. [本文引用: 4]
VilarinoM, RashidST, SuchyFP, McNabbBR, vander Meulen T, FineEJ, AhsanSD, MursaliyevN, SebastianoV, DiabSS, HuisingMO, NakauchiH, RossPJ. CRISPR/Cas9 microinjection in oocytes disables pancreas development in sheep Sci Rep, 2017,7(1):17472. [本文引用: 4]
FanZQ, PerisseIV, CottonCU, RegouskiM, MengQG, DombC, Van WettereAJ, WangZD, HarrisA, WhiteKL, PolejaevaIA. A sheep model of cystic fibrosis generated by CRISPR/Cas9 disruption of the CFTR gene JCI Insight, 2018,3(19):e123529. [本文引用: 3]
HuR, FanZY, WangBY, DengSL, ZhangXS, ZhangJL, HanHB, LianZX. Rapid communication: generation of FGF5 knockout sheep via the CRISPR/Cas9 system J Anim Sci, 2017,95(5):2019-2024. [本文引用: 2]
LiWR, HeSG, LiuCX, ZhangXM, WangLQ, LinJP, ChenL, HanB, HuangJC, LiuMJ. Ectopic expression of FGF5s induces wool growth in Chinese merino sheep Gene,2017,627:477-483. [本文引用: 2]
MaT, TaoJL, YangMH, HeCJ, TianXZ, ZhangXS, ZhangJL, DengSL, FengJZ, ZhangZZ, WangJ, JiPY, SongYK, HePL, HanHB, FuJC, LianZX, LiuGS. An AANAT/ASMT transgenic animal model constructed with CRISPR/Cas9 system serving as the mammary gland bioreactor to produce melatonin-enriched milk in sheep J Pineal Res, 2017,63(1). [本文引用: 3]
KimYG, ChaJ, ChandrasegaranS. Hybrid restriction enzymes: zinc finger fusions to Fok I cleavage domain Proc Natl Acad Sci USA, 1996,93(3):1156-1160. [本文引用: 1]
WolfeSA, NekludovaL, PaboCO. DNA recognition by Cys2His2 zinc finger proteins Annu Rev Biophys Biomol Struct, 2000,29:183-212. [本文引用: 1]
SmithJ, BergJM, ChandrasegaranS. A detailed study of the substrate specificity of a chimeric restriction enzyme Nucleic Acids Res, 1999,27(2):674-681. [本文引用: 2]
ValerieK, PovirkLF. Regulation and mechanisms of mammalian double-strand break repair Oncogene, 2003,22(37):5792-5812. [本文引用: 1]
BibikovaM, GolicM, GolicKG, CarrollD. Targeted chromosomal cleavage and mutagenesis in Drosophila using zinc-finger nucleases Genetics, 2002,161(3):1169-1175. [本文引用: 2]
MillerJ, McLachlanAD, KlugA. Repetitive zinc- binding domains in the protein transcription factor IIIA from Xenopus oocytes EMBO J, 1985,4(6):1609-1614. [本文引用: 1]
HwangWY, FuYF, ReyonD, MaederML, TsaiSQ, SanderJD, PetersonRT, YehJRJ, JoungJK. Efficient genome editing in zebrafish using a CRISPR-Cas system Nat Biotechnol, 2013,31(3):227-229. [本文引用: 2]
KomorAC, KimYB, PackerMS, ZurisJA, LiuDR. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage Nature, 2016,533(7603):420-424. [本文引用: 1]
ReesHA, LiuDR. Base editing: precision chemistry on the genome and transcriptome of living cells Nat Rev Genet, 2018,19(12):770-788. [本文引用: 1]
JinS, ZongY, GaoQ, ZhuZX, WangYP, QinP, LiangCZ, WangDW, QiuJL, ZhangF, GaoCX. Cytosine, but not adenine, base editors induce genome-wide off-target mutations in rice Science,2019,364(6437):292-295. [本文引用: 1]
DengSL, LiGD, YuK, TianXZ, WangF, LiWT, JiangWQ, JiPY, HanHB, FuJC, ZhangXS, ZhangJL, LiuYX, LianZX, LiuGS. RNAi combining Sleeping Beauty transposon system inhibits ex vivo expression of foot-and-mouth disease virus VP1 in transgenic sheep cells Sci Rep, 2017,7(1):10065. [本文引用: 3]
HanrahanJP, GreganSM, MulsantP, MullenM, DavisGH, PowellR, GallowaySM. Mutations in the genes for oocyte-derived growth factors GDF9 and BMP15 are associated with both increased ovulation rate and sterility in Cambridge and Belclare sheep (Ovis aries) Biol Reprod, 2004,70(4):900-909. [本文引用: 1]
ZhangZF, SunYW, DuW, HeSG, LiuMJ, TianCY. Effects of vertebral number variations on carcass traits and genotyping of Vertnin candidate gene in Kazakh sheep Asian-Australas J Anim Sci, 2017,30(9):1234-1238. [本文引用: 1]
ValetteM, PoitouC, Le BeyecJ, BouillotJL, ClementK, CzernichowS. Melanocortin-4 receptor mutations and polymorphisms do not affect weight loss after bariatric surgery PLoS One, 2012,7(11):e48221. [本文引用: 1]
ChmurzyńskaA. The multigene family of fatty acid-binding proteins (FABPs): function, structure and polymorphism J Appl Genet, 2006,47(1):39-48. [本文引用: 1]
WangX, NiuY, ZhouJ, ZhuH, MaB, YuH, YanH, HuaJ, HuangX, QuL, ChenY. CRISPR/Cas9-mediated MSTN disruption and heritable mutagenesis in goats causes increased body mass Anim Genet, 2018,49(1):43-51. [本文引用: 1]
GuoRH, WanYJ, XuD, CuiLB, DengMT, ZhangGM, JiaRX, ZhouWJ, WangZ, DengKP, HuangMR, WangF, ZhangYL. Generation and evaluation of Myostatin knock-out rabbits and goats using CRISPR/Cas9 system Sci Rep, 2016,6:29855. [本文引用: 1]
NiW, QiaoJ, HuSW, ZhaoXX, RegouskiM, YangM, PolejaevaIA, ChenCF. Efficient gene knockout in goats using CRISPR/Cas9 system PLoS One, 2014,9(9):e106718. [本文引用: 1]
HaoF, YanW, LiXC, WangH, WangYM, HuX, LiuX, LiangH, LiuDJ. Generation of cashmere goats carrying an EDAR gene mutant using CRISPR-Cas9-mediated genome editing Int J Biol Sci, 2018,14(4):427-436. [本文引用: 1]
PolejaevaIA, ChenSH, VaughtTD, PageRL, MullinsJ, BallS, DaiY, BooneJ, WalkerS, AyaresDL, ColmanA, CampbellKH. Cloned pigs produced by nuclear transfer from adult somatic cells Nature, 2000,407(6800):86-90. [本文引用: 1]
DavisBT, WangXJ, RohretJA, StruzynskiJT, MerricksEP, BellingerDA, RohretFA, NicholsTC, RogersCS. Targeted disruption of LDLR causes hypercholesterolemia and atherosclerosis in Yucatan miniature pigs PLoS One,2014,9(4):e93457. [本文引用: 1]
YangDS, YangHQ, LiW, ZhaoBT, OuyangZ, LiuZM, ZhaoY, FanNN, SongJ, TianJT, LiF, ZhangJF, ChangL, PeiDQ, ChenYE, LaiLX. Generation of PPARγ mono-allelic knockout pigs via zinc-finger nucleases and nuclear transfer cloning Cell Res, 2011,21(6):979-982. [本文引用: 1]
FengC, LiXR, CuiHT, LongC, LiuX, TianXH, PanDK, LuoYZ. Highly efficient generation of GGTA1 knockout pigs using a combination of TALEN mRNA and magnetic beads with somatic cell nuclear transfer J Integr Agr, 2016,15(7):1540-1549. [本文引用: 1]
ZouYL, LiZY, ZouYJ, HaoHY, LiN, LiQY. An FBXO40 knockout generated by CRISPR/Cas9 causes muscle hypertrophy in pigs without detectable pathological effects Biochem Biophys Res Commun, 2018,498(4):940-945. [本文引用: 1]
WangHT, ShenLC, ChenJY, LiuXJ, TanT, HuYQ, BaiXF, LiYX, TianKG, LiN, HuXX. Deletion of CD163 exon 7 confers resistance to highly pathogenic porcine reproductive and respiratory viruses on pigs Int J Biol Sci, 2019,15(9):1993-2005. [本文引用: 1]
YuHH, ZhaoH, QingYB, PanWR, JiaBY, ZhaoHY, HuangXX, WeiHJ. Porcine zygote injection with Cas9/sgRNA results in DMD-modified pig with muscle dystrophy Int J Mol Sci, 2016,17(10):1668. [本文引用: 1]
WhitworthKM, BenneJA, SpateLD, MurphySL, SamuelMS, MurphyCN, RichtJA, WaltersE, PratherRS, WellsKD. Zygote injection of CRISPR/Cas9 RNA successfully modifies the target gene without delaying blastocyst development or altering the sex ratio in pigs Transgenic Res, 2017,26(1):97-107. [本文引用: 1]
ZhouXY, WangLL, DuYN, XieF, LiL, LiuY, LiuCH, WangSQ, ZhangSB, HuangXX, WangY, WeiH. Efficient generation of gene-modified pigs harboring precise orthologous human mutation via CRISPR/Cas9- induced homology-directed repair in zygotes Hum Mutat, 2016,37(1):110-118. [本文引用: 1]
ParkKE, KaucherAV, PowellA, WaqasMS, SandmaierSES, OatleyMJ, ParkCH, TibaryA, DonovanDM, BlombergLA, LillicoSG, WhitelawCBA, MilehamA, TeluguBP, OatleyJM. Generation of germline ablated male pigs by CRISPR/Cas9 editing of the NANOS2 gene Sci Rep, 2017,7:40176. [本文引用: 1]
AppelMJ, van VeenHA, VietschH, SalaheddineM, NuijensJH, ZiereB, de LoosF. Sub-chronic (13-week) oral toxicity study in rats with recombinant human lactoferrin produced in the milk of transgenic cows Food Chem Toxicol, 2006,44(7):964-973. [本文引用: 1]
YuDW, WangJ, ZouHY, FengT, ChenL, LiJ, QiXL, LiZF, DuanXY, XuCL, ZhangL, LongX, LanJ, ChenC, WangC, XuXY, RenJL, ZhaoYQ, HuXX, LianZX, MenHS, PanDD, LiN, CapecchiMR, DuXG, ZhaoYF, WuS. Silencing of retrotransposon-derived imprinted gene RTL1 is the main cause for postimplantational failures in mammalian cloning Proc Natl Acad Sci USA, 2018,115(47):E11071-11080. [本文引用: 1]
LiangS, JinYX, YuanB, ZhangJB, KimNH. Melatonin enhances the developmental competence of porcine somatic cell nuclear transfer embryos by preventing DNA damage induced by oxidative stress Sci Rep, 2017,7(1):11114. [本文引用: 1]
HashimotoM, TakemotoT. Electroporation enables the efficient mRNA delivery into the mouse zygotes and facilitates CRISPR/Cas9-based genome editing Sci Rep, 2015,5:11315. [本文引用: 1]
ZhangYM, WangQQ, LiuKL, GaoEE, GuanH, HouJ. Treatment of donor cells with recombinant KDM4D protein improves preimplantation development of cloned ovine embryos Cytotechnology, 2018,70(5):1469-1477. [本文引用: 1]
AppelMJ, van VeenHA, VietschH, SalaheddineM, NuijensJH, ZiereB, de LoosF. Sub-chronic (13-week) oral toxicity study in rats with recombinant human lactoferrin produced in the milk of transgenic cows Food Chem Toxicol, 2006,44(7):964-973. [本文引用: 1]
 ,1,2, 李亭亭3, 黄勋1,2, 马润林
,1,2, 李亭亭3, 黄勋1,2, 马润林
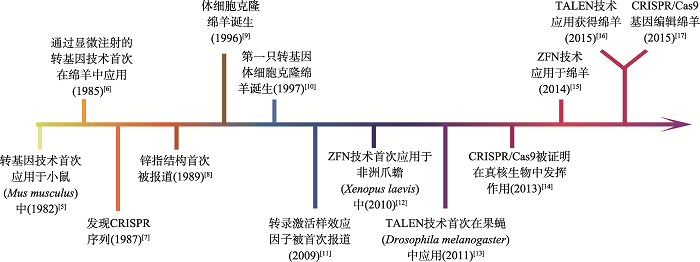 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT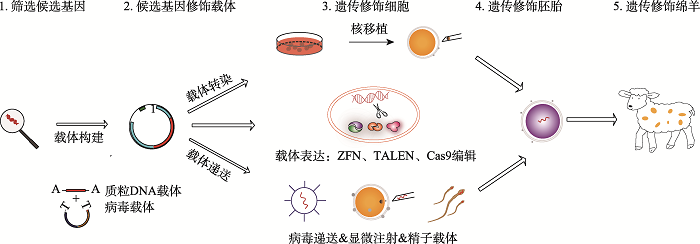 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT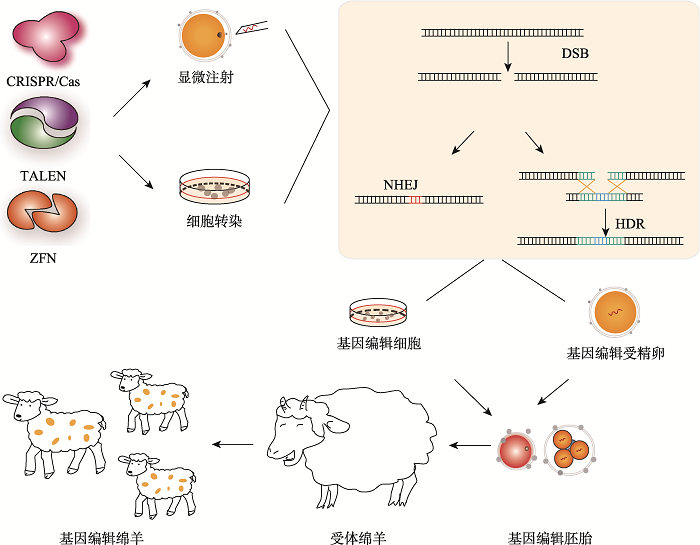 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT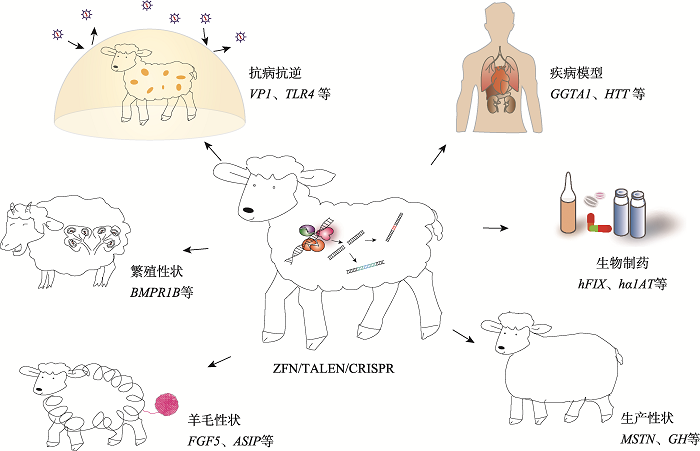 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT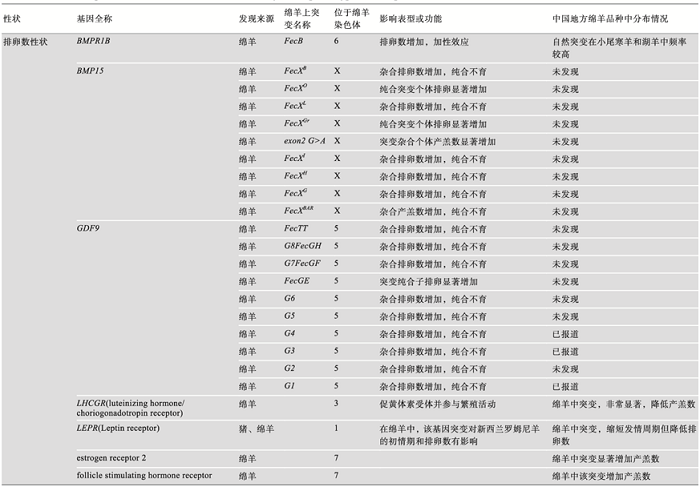

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}