 ,, Ья,ЙѓжнДѓбЇЩњУќПЦбЇбЇдК,Йѓбє 550025
,, Ья,ЙѓжнДѓбЇЩњУќПЦбЇбЇдК,Йѓбє 550025The F-box gene Ppa promotes lipid storage in Drosophila
Guangwu Yang,, Yuan Tian,College of Life Sciences, Guizhou University, Guiyang 550025, ChinaЭЈбЖзїеп: ЬяЃЌВЉЪПЃЌИБНЬЪкЃЌбаОПЗНЯђЃКЙћгЌжЌЗОДњаЛЁЃE-mail:ytian1@gzu.edu.cn
д№ШЮБрМ: бжбд
ЪеИхШеЦк:2021-03-5аоЛиШеЦк:2021-04-28ЭјТчГіАцШеЦк:2021-06-20
| ЛљН№зЪжњ: |
Received:2021-03-5Revised:2021-04-28Online:2021-06-20
| Fund supported: |
зїепМђНщ About authors
бюЙтЮф,дкЖСЫЖЪПбаОПЩњ,зЈвЕЗНЯђЃКЗЂг§ЩњЮябЇЁЃE-mail:

еЊвЊ
жЌжЪЪЧЙЙГЩЩњЮяЬхЕФживЊГЩЗжжЎвЛ,жЌжЪДњаЛЕФОЋШЗЕїНкКЭЮШЬЌЮЌГжЖдШЫРрНЁПЕжСЙиживЊЁЃЗКЫиЛЏЭООЖЭЈЙ§НЕНтжЌжЪЯрЙиЕААзРДЕїПижЌжЪДњаЛЁЃPpa (partner of paired)БрТывЛжжF-boxЕААз,КѓепЪЧSCF (Skp1-Cullin1-F-box)ЗКЫиЛЏИДКЯЬхГЩдБжЎвЛЁЃвбгаЕФбаОПБэУїPpaдкЕїПиЙћгЌЬхНкЕФе§ГЃЗЂг§КЭзХЫПСЃзщЕААзЕФе§ШЗЖЈЮЛЗНУцЗЂЛгживЊЕФзїгУ,ЕЋдкжЌЗОДњаЛЗНУцЕФЙІФмбаОПШДЮДМћБЈЕРЁЃБОбаОПвдКкИЙЙћгЌ(Drosophila melanogaster)зїЮЊбаОПЖдЯѓ,ЬНОПСЫPpaдкжЌЗОДЂДцжаЕФЙІФмЁЃЭЈЙ§гыТЬЩЋгЋЙтЕААзШкКЯБэДя,МьВтPPAЕФбЧЯИАћЖЈЮЛ;гІгУCRISPR/Cas9ММЪѕ,ЙЙНЈPpaЕФШБЪЇЭЛБфЬх;ЭЈЙ§BODIPY 493/503ЛђNile redжЌЕЮШОЩЋ,ЖдШБЪЇЭЛБфЬхКЭГЌСПБэДяPpaЙћгЌЕФжЌЕЮаЮЬЌИФБфНјааЗжЮіЁЃЫцКѓ,дкШБЪЇЭЛБфЬхжаГЌСПБэДяPpaвдбщжЄЦфЙІФмЁЃНсЙћБэУї,PPA-GFPШкКЯЕААзЖЈЮЛгкЭйвКЯйКЭжЌЗОЬхЕФЯИАћКЫжаЁЃгыЖдеезщЙћгЌЯрБШ,PpaШБЪЇЭЛБфЬхБэЯжЮЊжЌЕЮБфаЁ;ГЌСПБэДяPpaЯдЪОГіжЌЕЮБфДѓЁЃдкЭЛБфЬхжаГЌСПБэДяPpaФмЙЛНЋжЌЕЮЛиИДжСе§ГЃзДЬЌЁЃзлЩЯЫљЪі,БОбаОПНвЪОСЫPpaдкЙћгЌжаОпгаДйНјжЌЗОДЂДцЕФЙІФмЁЃ
ЙиМќДЪЃК
Abstract
Lipid is one of the important components of living organisms. The precise regulation and homeostasis maintenance of lipid metabolism are essential to human health. The ubiquitination pathway regulates lipid metabolism by degrading lipid-related proteins. Ppa encodes an F-box protein, which is a member of the SCF ubiquitination complex. Previous studies reported that Ppa regulated the body segmentation and the correct localization of centromere histones, while its function in lipid metabolism has not been reported. In this study, Drosophila melanogaster was used to explore the function of Ppa in lipid storage. The subcellular localization of PPA was detected by fusion with green fluorescent protein. The deletion mutant of Ppa was constructed via CRISPR/Cas9 technology. The morphological changes of lipid droplets in deletion mutants and Ppa overexpression flies were analyzed by BODIPY 493/503 or Nile red staining. Further, Ppa was overexpressed in the deletion mutant to verify its function. The results showed that PPA-GFP fusion protein were localized in the nuclei of salivary gland and fat body. Compared with the control flies, the lipid droplets in Ppa deletion mutants became smaller, and overexpression of Ppa exhibited larger lipid droplets. Overexpression of Ppa in the deletion mutant could restore the lipid droplets to normal state. In summary, this study demonstrated that Ppa could promote lipid storage in Drosophila.
KeywordsЃК
PDF (758KB)дЊЪ§ОнЖрЮЌЖШЦРМлЯрЙиЮФеТЕМГіEndNote|Ris|BibtexЪеВиБОЮФ
БОЮФв§гУИёЪН
бюЙтЮф, Ья. ЙћгЌF-boxЛљвђPpaДйНјжЌЗОДЂДц. вХДЋ[J], 2021, 43(6): 615-622 doi:10.16288/j.yczz.21-084
Guangwu Yang.
жЌжЪДњаЛЮЩТвгыЗЪХжЁЂИпбЊбЙЁЂЬЧФђВЁЕШДњаЛзлКЯеїЯЂЯЂЯрЙи[1]ЁЃжЌЕЮ(lipid droplets, LDs)ЪЧгЩЕЅВуСзжЌЁЂжаадКЫаФжЌжЪКЭЯрЙиЕААззщГЩЕФЯИАћЦї,ДѓЖрЪ§ЖЏЮяЕФжааджЌжЪЖМДЂДцдкжЌЕЮжа[2]ЁЃжЌЕЮдкаэЖрЗНУцЖМЗЂЛгзХживЊЕФЩњЮябЇЙІФмЃКЪзЯШ,ЫќЪЧЯИАћФкжЌЗОЫсзЊдЫЕФживЊЮЛЕу,жЌЗОЫсКЌСПИпЕФИпжЌЯИАћФмЙЛЭЈЙ§жЌЕЮЯђЯрСйЕФЕЭжЌЯИАћзЊдЫжЌЗОЫс[3];ЦфДЮ,ЫќЪЧЬиЖЈВЁЖОЕФзщзАЦНЬЈ,ИЮЯИАћжаЕФжЌЕЮЮЊМЋЕЭУмЖШжЌЕААзЕФВњЩњЬсЙЉжЌжЪЛљДЁ,ЖјБћИЮВЁЖОФмЙЛЧжШыМЋЕЭУмЖШжЌЕААзЗжУкЭООЖВЂИНзХдкЦфБэУц,ДйНјБћИЮВЁЖОвдЕЭУмЖШжЌВЁЖОПХСЃЕФаЮЪНЪЭЗХ[4];дкЩёОЕїНкЗНУц,вХДЋадОЗТЮадНиЬБЯрЙиЛљвђDDHD2ЕФЧУГ§аЁЪѓБэЯжГідЫЖЏаЕїКЭШЯжЊФмСІЕФШБЯн,ВЂЧвдкЩёОдЊжаЯдЪОГіДѓСПЕФвьЮЛжЌЗОРлЛ§[5]ЁЃжЌЕЮЕФДѓаЁЪмЕНКЫаФжЌжЪКЌСПЁЂЕЅВуСзжЌзщГЩКЭДйНјжЌЕЮШкКЯЯрЙиЕААзЕФгАЯь[6,7]ЁЃДѓжЌЕЮЕФаЮГЩгаСНжжЛњжЦ,вЛжжЪЧжЌЕЮЕФЩњГЄКЭХђеЭ,ЭЈГЃЪЧЭЈЙ§НЋжааджЌжЪЬэМгЕНжЌЕЮКЫаФРДЪЕЯж,СэвЛжжЪЧгЩНЯаЁжЌЕЮШкКЯаЮГЩ,ЕЋШкКЯЛюадКмЕЭ[8]ЁЃ
ецКЫЩњЮяжаЙуЗКДцдквЛРрНазіF-boxЕФЕААзМвзх[9]ЁЃF-boxЕААзМвзхЕФNЖЫОпгаЬиеїадЕФF-boxНсЙЙгђ,ИКд№гыЦфЫћЕААзНсКЯаЮГЩSCFИДКЯЬх,ИУИДКЯЬхЪЧвЛРрE3ЗКЫиЛЏСЌНгУИ,ЦфCЖЫОпгаЕзЮяЪЖБ№НсЙЙгђ[10]ЁЃF-boxЕААзгыНсКЯЕААзSkp1ЁЂЙЧМмЕААзCullin1КЭRING-fingerЕААзRbx1ЙВЭЌзщГЩSCFИДКЯЬхЁЃИУИДКЯЬхеаФМаЏДјЗКЫиЗжзгЕФE2НсКЯУИВЂЪЖБ№ЕзЮяЕААз,ВЮгыбЁдёадНЕНтЩњЮяЛюадЕААзжЪЕФЗКЫи-ЕААзУИЬхЭООЖ(ubiquitin proteasome pathway, UPP),дкЯИАћЕђЭіЁЂЯИАћжмЦкЕїПиЁЂаХКХзЊЕМКЭУтвпгІМЄЕШЗНУцЗЂЛгживЊзїгУ[11,12]ЁЃбаОПБэУї,UPPНщЕМЖрИіжЌЕЮЯрЙиЕААзЕФНЕНт,Р§Шч,PLIN1ЪЧЮЛгкжЌЕЮБэУцЕФЬиеїадНсКЯЕААз,ФмЙЛвжжЦжЌжЪНЕНт,дкаЁЪѓЕФ3T3-L1жЌЗОЯИАћжа,PLIN1гыбЁдёадздЪЩЪмЬхSQSTM1ЯрЛЅзїгУ,ВЂЭЈЙ§гыЗКЫиЛЏЕААзЬивьадНсКЯРДНЕНтPLIN1[13,14]ЁЃ
PpaЪЧЙћгЌжаМЬslmbКѓЕкЖўИіБЛЗЂЯжЕФБрТыF-boxЕААзЕФЛљвђ[15]ЁЃбаОПБэУї,PpaдкдчЦкХпЬЅЗЂг§Й§ГЬжаЗЂЛгзїгУЁЃPpaдкЬхФкгыгЩeven skipped(eve)БрТыЕФEveЙВЭЌзїгУ,вжжЦзЊТММЄЛювђзгPrd(гЩpairedБрТы,prd),ЕїПиАќРЈen (engrailed)дкФкЕФЬхНкМЋадЛљвђЕФБэДяЁЃдкeveе§ГЃБэДяЕФЯИАћжа,Ppa mRNAБЛEveвжжЦ,ШЗБЃСЫPrdБЛМЄЛю,ДгЖјМЄЛюЯТгЮАаЛљвђenЕФБэДя;ЖјдкeveВЛБэДяЕФЯИАћжа,Ppa mRNAНєАЄPrd,ВЂНЋPrdЬѕДјНЕНт,вжжЦЯТгЮАаЛљвђenЕФБэДя[16,17]ЁЃДЫЭт,PpaдкзХЫПСЃЕФЮШЖЈадЮЌГжЗНУцвВЗЂЛгживЊзїгУ,зХЫПСЃзщЕААзCIDОіЖЈзХЫПСЃЕФЬиадКЭЙІФм,ГЌСПБэДяcidЛсЪЙЕУИУЕААзДэЮѓЖЈЮЛгкШОЩЋЬхЕФЗЧзХЫПСЃЧјгђ,ЕМжТвьЮЛзХЫПСЃЕФВњЩњ,ДгЖјв§ЗЂШОЩЋЬхЕФДэЮѓЗжРы[18]ЁЃPpaНщЕМЕФЗКЫиЛЏЭООЖФмЙЛНЕНтДэЮѓЖЈЮЛЕФCID,ЪЙИУЕААзжиаТЖЈЮЛгкзХЫПСЃ[19]ЁЃБОбаОПЭЈЙ§ЕААзЖЈЮЛЁЂЭЛБфЬхЗжЮіКЭЛљвђЭьОШЪЕбщ,ЗЂЯжPpaПЩвдДйНјЙћгЌжаЕФжЌЗОДЂДцЁЃ
1 ВФСЯгыЗНЗЈ
1.1 ЙћгЌЦЗЯЕ
ЪЕбщжаЫљгУЕНЕФЙћгЌОљдкгёУзУцХрбјЛљжаЫЧбј,бЯИёАДееЪЕбщЙцеТжЦЖШВйзї,ЙћгЌРДдДМАгУЭОМћБэ1ЁЃTable 1
Бэ1
Бэ1БОбаОПжаЫљгУЕФЙћгЌЦЗЯЕ
Table 1
| ЛљвђаЭ | РДдД | гУЭО |
|---|---|---|
| w1118 | жаЙњПЦбЇдКвХДЋгыЗЂг§ЩњЮябЇбаОПЫљЛЦбЋбаОПдБЪЕбщЪв | Ждеезщ |
| w*; ln(2LR)noc4LScorv9R, b1/CyO, P {ActGFP}JMR1 | жаЙњПЦбЇдКвХДЋгыЗЂг§ЩњЮябЇбаОПЫљЛЦбЋбаОПдБЪЕбщЪв | ЦНКтжТЫРЯЕ |
| da-Gal4 | ЧхЛЊДѓбЇЙћгЌжааФ | ШЋЩэБэДяЕФGal4 |
| y1M{nos-Cas9.P}ZH-2A w* | Bloomington | БэДяCas9ЕААз |
| w;L/CyO | БОЪЕбщЪв | ЦНКтжТЫРЯЕ |
| Ppa-sgRNA | БОЪЕбщЙЙНЈ | БэДяСНЖЮsgRNA |
| UAS-Ppa-RA/RC | БОЪЕбщЙЙНЈ | ГЌСПБэДяPPA |
| UAS-Ppa-RA/RC-eGFP | БОЪЕбщЙЙНЈ | PPAЕФбЧЯИАћЖЈЮЛ |
аТДАПкДђПЊ|ЯТдиCSV
1.2 ЙћгЌзщжЏШОЩЋ
НЋ3СфгзГцдк1×PBSЛКГхвКжаНтЦЪ,ЛёЕУжЌЗОЬхКЭЭйвКЯйЁЃНЋзщжЏЗХШы4%ЖрОлМзШЉжаЙЬЖЈ30 minКѓ,гУ1×PBSЧхЯД2ДЮ,УПДЮ5 minЁЃЪЙгУжЌЕЮШОСЯBODIPY 493/503ЛђепNile redБмЙтШОЩЋ30 min,ЪЙгУЯИАћКЫШОСЯDAPIБмЙтШОЩЋ10 min,ЪЙгУЯИАћЙЧМмШОСЯPhalloidin 594БмЙтШОЩЋ30 minЁЃгУ1×PBSЧхЯД3ДЮ,УПДЮ5 minЁЃдкдиВЃЦЌЩЯЕЮМг30 μLЕФ70%ИЪгЭ,НЋШОЩЋЭъБЯЕФбљЦЗзЊвЦжСдиВЃЦЌЩЯ,ИЧВЃЦЌбЙЦЌ,зюКѓгУжИМзгЭЗтЦЌ,НЋжЦзїЭъГЩЕФЦЌзгдкМЄЙтЙВОлНЙЯдЮЂОЕЩЯЩЈУшГЩЯёЁЃ1.3 PPAЕФбЧЯИАћЖЈЮЛЗжЮі
ЭЈЙ§дкPpa-RA/RCБрТыЧјЕФCЖЫЬэМгБэДяТЬЩЋгЋЙтЕААзЕФGFPађСа,ЙЙНЈЕУЕНзЊЛљвђЙћгЌUAS-Ppa- RA/RC-eGFPЁЃ29ЁцЯТ,РћгУda-Gal4Ч§ЖЏUAS-Ppa- RA/RC-eGFPЕФШЋЩэБэДя,НтЦЪ3СфгзГцЕФЭйвКЯйКЭжЌЗОЬх,ЪЙгУDAPIКЭPhalloidin 594ШОЩЋБъМЧЯИАћКЫКЭЯИАћЙЧМм,МЄЙтЙВОлНЙЯдЮЂОЕЯТЩЈУшГЩЯёЁЃ1.4 sgRNAБэДядиЬхЕФЙЙНЈ
ЭЈЙ§ЭјеОwww.flyrnai.org/crispr2ЩшМЦСНЖЮsgRNAађСа,ЦфЮЛгкPpaЕФCDSЧјЕФЕк78~97 ntКЭЕк518~538 nt,ВЂдкв§ЮяЧАЬэМгBbsIУИЧаЮЛЕуЁЃНЋЩшМЦКУЕФsgRNAађСадкFlybaseЩЯНјааађСаБШЖд,ШЗБЃдкЛљвђзщжаУЛгаЦфЫћХфЖдЧјгђЁЃsgRNAМАМьВтв§ЮяађСаМћБэ2ЁЃTable 2
Бэ2
Бэ2БОбаОПжаЫљгУЕФв§ЮяађСа
Table 2
| в§ЮяУћГЦ | в§ЮяађСа(5′→3′) |
|---|---|
| Ppa-sgRNA1-F | gtcgCGGATCCGGGAGCAACGGAG |
| Ppa-sgRNA1-R | aaacCTCCGTTGCTCCCGGATCCG |
| Ppa-sgRNA2-F | gtcgTCAGGTTTGCACCGCATGGC |
| Ppa-sgRNA2-R | aaacGCCATGCGGTGCAAACCTGA |
| Ppa-chk-F | CCCTTCCATCATCAAACA |
| Ppa-chk-R | GAACGCCCAGGACCAAAT |
аТДАПкДђПЊ|ЯТдиCSV
ИљОнЩшМЦЕФsgRNAађСаКЯГЩЕЅСДЙбКЫмеЫсЦЌЖЮ,ОЙ§ЭЫЛ№аЮГЩСНЖЫгаеГадЭЛГіФЉЖЫЕФЫЋСДЦЌЖЮЁЃгУT4СЌНгУИНЋsgRNA1КЭsgRNA2ЕФЭЫЛ№ЦЌЖЮЗжБ№СЌНгЕНBbsIУИЧаКѓЕФЯпадЛЏдиЬхpMD18T-U63- BsmB1ЩЯЁЃОЙ§зЊЛЏЪЕбщ,ЩИбЁЕУЕНСНИіСЌНгГЩЙІЕФжЪСЃ,pMD18T-U63-Ppa-sgRNA1КЭpMD18T-U63- Ppa-sgRNA2ЁЃЭЈЙ§PCRРЉдіИјU63-Ppa-sgRNA1КЭU63-Ppa-sgRNA2ЬэМгНгЭЗађСаЁЃРћгУGolden-gateСЌНгНЋСНЖЮгЩU63ЦєЖЏзгЧ§ЖЏЕФsgRNAСЌНгЕНжаМфдиЬхPCR/TOPO/GW-BsaI-BbЁЃзюКѓНЋзщзАКУЕФжЪСЃЭЈЙ§LRЗДгІжизщСЌНг,ЕУЕНpUAST-attB-Ppa-sgRNAsБэДядиЬхЁЃ
1.5 ЭЛБфЬхМјЖЈгыжЌЕЮБэаЭЗжЮі
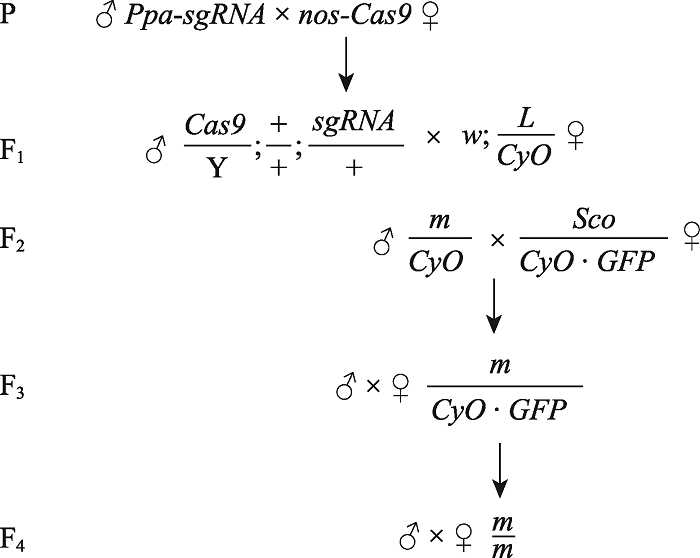
НЋЙЙНЈЕФpUAST-attB-Ppa-sgRNAжЪСЃЫЭжСжщКЃСЊКЯЛЊвцЙЋЫОНјааЯдЮЂзЂЩф,ЕУЕНзЊЛљвђЙћгЌPpa-sgRNAЁЃНЋPpa-sgRNAгыБэДяCas9ЕААзЕФЙћгЌдгНЛ,КѓДњжаЕУЕНЭЌЪББэДяCas9ЕААзгыsgRNAЕФЙћгЌЁЃЪЙгУPpa-chk-FКЭPpa-chk-RНјааFounderЕФPCRМјЖЈ,ЭЈЙ§дгНЛЕУЕНДПКЯЕФЭЛБфЙћгЌ,ВЂЭЈЙ§PCRРЉдіКЭDNAВтађБШЖдШЗЖЈЭЛБфЮЛЕуЁЃОпЬхЕФдгНЛВпТдШчЭМ1ЫљЪОЁЃЭМ1
 аТДАПкДђПЊ|ЯТдидЭМZIP|ЩњГЩPPT
аТДАПкДђПЊ|ЯТдидЭМZIP|ЩњГЩPPTЭМ1РћгУCRISPR/Cas9ММЪѕЕФЛљвђЧУГ§дгНЛВпТд
Fig. 1The gene knockout strategy mediated by CRISPR/Cas9
баОПбЁШЁСЫСНжъЭЛБфЬхPpa1КЭPpa2вдМАСНепЕФдгНЛКѓДњPpa1/Ppa2НјааЗжЮі,НтЦЪЛёЕУ3СфгзГцЕФжЌЗОЬх,ЪЙгУDAPIКЭBODIPY 493/503БъМЧЯИАћКЫКЭжЌЕЮ,МЄЙтЙВОлНЙЯдЮЂОЕЯТЩЈУшГЩЯёЁЃЭЈЙ§Image-Pro Plus 6.0ШэМўЖдЬиЖЈУцЛ§Фк(143 μm×143 μm)ЕФВЛЭЌжБОЖжЌЕЮНјааЪ§СПЭГМЦ,ЖдеезщгыЪЕбщзщИїбЁШЁ3ИібљБОНјааЭГМЦЁЃЭЈЙ§GraphPad Prism 8ШэМўНјааЭГМЦбЇЗжЮіВЂЛцжЦЭГМЦЭМЁЃ
1.6 ГЌСПБэДяPpaЕФжЌЕЮБэаЭЗжЮі
T-AПЫТЁPpa-RA/RCЕФБрТыЧј,ЭЈЙ§EcoRⅠКЭKpnⅠСЌНгжСpUAST-attB,ЛёЕУСЫзЊЛљвђЙћгЌUAS- Ppa-RA/RCЁЃ29ЁцЯТ,РћгУda-Gal4Ч§ЖЏUAS-Ppa-RA /RCЕФШЋЩэБэДя,НтЦЪ3СфгзГцЕФжЌЗОЬх,ЪЙгУDAPIКЭNile redШОЩЋБъМЧЯИАћКЫКЭжЌЕЮ,МЄЙтЙВОлНЙЯдЮЂОЕЯТЩЈУшГЩЯёЁЃЭЈЙ§Image-Pro Plus 6.0ШэМўЖдЬиЖЈУцЛ§Фк(143 μm×143 μm)ЕФВЛЭЌжБОЖжЌЕЮНјааЪ§СПЭГМЦ,ЖдеезщгыЪЕбщзщИїбЁШЁ3ИібљБОНјааЭГМЦЁЃЭЈЙ§GraphPad Prism 8ШэМўНјааЭГМЦбЇЗжЮіВЂЛцжЦЭГМЦЭМЁЃ1.7 ЛљвђЭьОШЪЕбщ
ЭЈЙ§вХДЋВйзї,ЙЙНЈЕУЕНPpa1;da>UAS-Ppa- RA/RCЕФЙћгЌЁЃ29ЁцЯТ,дкЭЛБфЬхPpa1жаРћгУda-Gal4Ч§ЖЏUAS-Ppa-RA/RCЕФШЋЩэБэДя,НтЦЪ3СфгзГцЕФжЌЗОЬх,ЪЙгУDAPIКЭBODIPY 493/503ШОЩЋБъМЧЯИАћКЫКЭжЌЕЮ,МЄЙтЙВОлНЙЯдЮЂОЕЩЈУшГЩЯёЁЃ2 НсЙћгыЗжЮі
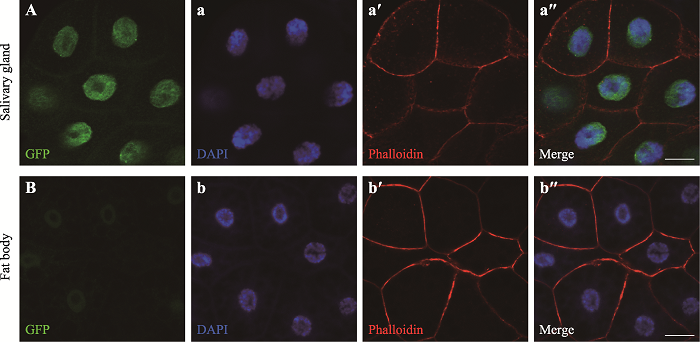
2.1 PPAШкКЯгЋЙтЕААзЖЈЮЛгкЯИАћКЫ
дкЧАЦкЙЄзїжа,ЗЂЯжГЌСПБэДяPpaФмЙЛдіЧПЙћгЌЭйвКЯйЕФвьЮЛжЌЗОРлЛ§ЁЃБОбаОПЪзЯШЭЈЙ§гыТЬЩЋгЋЙтЕААзШкКЯБэДя,ЖдPPAЕФЖЈЮЛНјааЗжЮіЁЃОВщFlybaseЕУжЊ,PpaЛљвђОпга3ИізЊТМБО,ЦфжаRAКЭRCОпгаЯрЭЌЕФБрТыЧј,RAгыRBЕФmRNAађСавЛжТ,ЧвRBЕФБрТыЧјдкRA/RCЕФЛљДЁЩЯдНЙ§СЫвЛИіжежЙУмТызгUGAЁЃБОбаОППЫТЁСЫPpa-RA/RCЕФБрТыЧј,ЙЙНЈСЫШкКЯЕААзБэДядиЬхВЂЕУЕНзЊЛљвђЙћгЌUAS- Ppa-RA/RC-eGFPЁЃЭЈЙ§da-Gal4Ч§ЖЏUAS-Ppa-RA/ RC-eGFPЕФШЋЩэБэДя,ЗЂЯжPPA-PA/PCдк3СфгзГцЭйвКЯйЕФЯИАћКЫжаМьВтГіЧПСвЕФгЋЙтаХКХ(ЭМ2A),ВЂдк3СфгзГцжЌЗОЬхЕФЯИАћКЫжаМьВтГіНЯШѕЕФгЋЙтаХКХ(ЭМ2B),БэУїPpaПЩФмВЮгыжЌжЪДњаЛЁЃЭМ2
 аТДАПкДђПЊ|ЯТдидЭМZIP|ЩњГЩPPT
аТДАПкДђПЊ|ЯТдидЭМZIP|ЩњГЩPPTЭМ2PPA-GFPдк3СфгзГцжаЕФбЧЯИАћЖЈЮЛ
ЪЙгУda-Gal4Ч§ЖЏUAS-Ppa-RA/RC-eGFPЕФШЋЩэБэДяЁЃAЃКЭйвКЯйЕФGFPаХКХ;aЃКЭйвКЯйЕФЯИАћКЫШОЩЋ;aʹЃКЭйвКЯйЕФЯИАћЙЧМмШОЩЋ;aʺЃКЭйвКЯйЕФгЋЙтаХКХКЯВЂЭМЁЃBЃКжЌЗОЬхЕФGFPаХКХ;bЃКжЌЗОЬхЕФЯИАћКЫШОЩЋ;bʹЃКжЌЗОЬхЕФЯИАћЙЧМмШОЩЋ;bʺЃКжЌЗОЬхЕФгЋЙтаХКХКЯВЂЭМЁЃDAPI (РЖЩЋ)ЃКЯИАћКЫШОЩЋ;GFP (ТЬЩЋ)ЃКPPA-GFPЕФШкКЯгЋЙтЕААз;Phalloidin (КьЩЋ)ЃКЯИАћЙЧМмШОЩЋ;Salivary glandЃКЭйвКЯй;Fat bodyЃКжЌЗОЬхЁЃБъГпЃК25 μmЁЃ
Fig. 2Subcellular localization of PPA-GFP in the 3rd instar larvae
2.2 PpaШБЪЇЭЛБфЬхЕФЛёЕУгыжЌЕЮБэаЭЗжЮі
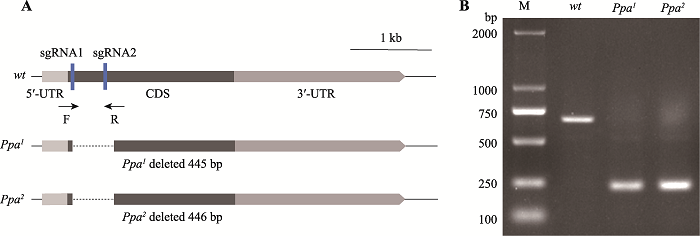
ЮЊСЫНјвЛВНЬНОПPpaЖджЌЗОДњаЛЕФгАЯь,БОбаОПРћгУCRISPR/Cas9ММЪѕЙЙНЈЛљвђШБЪЇЭЛБфЬхЁЃЪзЯШ,ЙЙНЈСЫattB-Ppa-sgRNAБэДядиЬх,ЭЈЙ§ЯдЮЂзЂЩфЕУЕНБэДяsgRNAЕФзЊЛљвђЙћгЌPpa-sgRNAЁЃНЋPpa-sgRNAгыБэДяCas9ЕААзЕФЙћгЌдгНЛ,ЖдPpaНјааЛљвђЧУГ§ЁЃЭЈЙ§ЖддгНЛКѓДњЕФвХДЋЩИбЁКЭPCRМјЖЈ,ЕУЕНСЫСНжъДПКЯПЩЛюЕФШБЪЇЭЛБфЬхPpa1КЭPpa2,ОпЬхВпТдЯъМћВФСЯгыЗНЗЈЁЃЫцКѓЖдЭЛБфЬхНјааВтађМјЖЈ,ЗЂЯжPpa1ШБЪЇСЫ445 bp,Ppa2ШБЪЇСЫ446 bp,ШчЭМ3ЫљЪО,ШБЪЇЕФЦЌЖЮЮЛгкБрТыЧјЕФЧАЖЫЁЃЭМ3
 аТДАПкДђПЊ|ЯТдидЭМZIP|ЩњГЩPPT
аТДАПкДђПЊ|ЯТдидЭМZIP|ЩњГЩPPTЭМ3ЭЛБфЬхжаЕФШБЪЇЧјгђЪОвтЭМгыPCRРЉдіФ§НКЕчгОЭМ
AЃКЭЛБфЬхШБЪЇЧјгђЪОвтЭМЁЃPpaжЛгавЛИіЭтЯдзг,РЖЩЋЗНПђБэЪОsgRNAЮЛЕу,ащЯпБэЪОШБЪЇЦЌЖЮЁЃBЃКЭЛБфЬхЕФPCRРЉдіНсЙћ(ЪЙгУМьВтв§ЮяPpa-chk-FКЭPpa-chk-RНјааМјЖЈ)ЁЃMЃК2 kb DNA marker;wtЃКЖдеезщ(w1118);Ppa1КЭPpa2ЃКЦЌЖЮШБЪЇЕШЮЛЛљвђЁЃ
Fig. 3The schematic diagram of deletion regions in mutants and gel electrophoresis of PCR amplification
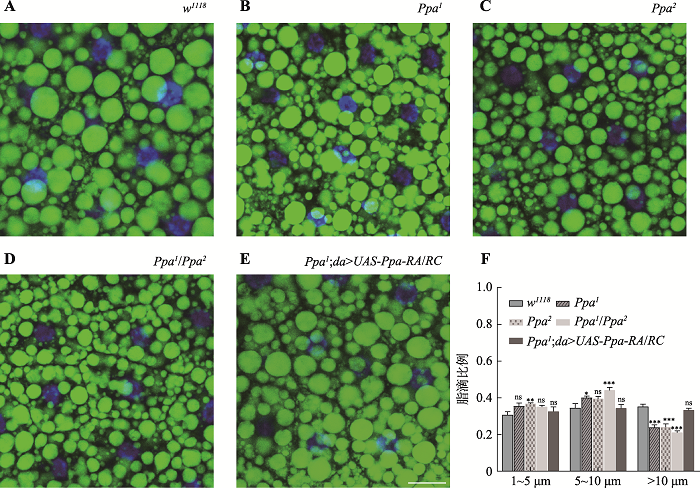
ЭЈЙ§BODIPYЖд3СфгзГцжЌЗОЬхжаЕФжЌЕЮНјааШОЩЋБъМЧ,ЗЂЯжгыЖдеезщw1118ЯрБШ,ЭЛБфЬхPpa1КЭPpa2жаЕФжЌЕЮУїЯдМѕаЁ(ЭМ4ЃКA~C)ЁЃдкЖдеезщжаЙлВьЕНжБОЖНЯДѓЕФДѓжЌЕЮ,ЕЋдкЭЛБфЬхжадђБэЯжЮЊаЁжЌЕЮЕФдіМгЁЃгЩгкCRISPR/Cas9ММЪѕОпгаЧБдкЕФЭбАааЇгІ,ЙЪБОбаОПЖдСНИіЦЌЖЮШБЪЇЕШЮЛЛљвђЕФЗДЪНдгКЯЭЛБфЬхPpa1/Ppa2ЕФжЌЕЮНјааСЫЗжЮі,вдЯћГ§вХДЋБГОАЩЯЕФЦфЫћЮДжЊЭЛБфЖдБэаЭЕФгАЯь,баОПЗЂЯжPpa1/Ppa2ЕФжЌЕЮНЯЖдеезщУїЯдМѕаЁ,аЁжЌЕЮЪ§СПУїЯддіЖр(ЭМ4D)ЁЃЫцКѓЖдЬиЖЈУцЛ§ФкВЛЭЌжБОЖЕФжЌЕЮНјааЭГМЦ,ЗЂЯжВЛЭЌжБОЖЕФжЌЕЮЫљеМЕФБШР§ОпгаЯджјВювьЁЃгыЖдеезщЯрБШ,ЭЛБфЬхБэЯжГі10 μmвдЩЯДѓжЌЕЮМѕЩй(ЭМ4F)ЁЃЮЊСЫНјвЛВНбщжЄЭЛБфЬхжажЌЕЮМѕаЁЕФБэаЭ,ЫцКѓдкЭЛБфЬхPpa1жаГЌСПБэДяPpa,ЗЂЯжжЌЕЮНЯЭЛБфЬхУїЯддіДѓ,ЕЋНЯЖдеезщЮоЯджјВювь(ЭМ4ЃКEКЭF),ЫЕУїPpaФмЙЛЛиИДЭЛБфЬхжажЌЕЮМѕаЁЕФБэаЭ,ВЂЧвЬсЪОPpaЕФШБЪЇЪЧЕМжТжЌЕЮМѕаЁЕФдвђ,вВНјвЛВННвЪОСЫPpaВЮгыжЌЗОДЂДцЕФЕїПиЁЃ
ЭМ4
 аТДАПкДђПЊ|ЯТдидЭМZIP|ЩњГЩPPT
аТДАПкДђПЊ|ЯТдидЭМZIP|ЩњГЩPPTЭМ4PpaШБЪЇЭЛБфЬхдк3СфгзГцжаЕФжЌЕЮжБОЖБфаЁ
A~EЃК3СфгзГцЕФжЌЕЮШОЩЋЭМЁЃw1118ЃКЖдеезщ;Ppa1КЭPpa2ЃКЦЌЖЮШБЪЇЕШЮЛЛљвђ;Ppa1/Ppa2ЃКСНИіЕШЮЛЛљвђЕФЗДЪНЭЛБфЬх;Ppa1;da>UAS-Ppa-RA/RCЃКЭЛБфЬхжаГЌСПБэДяPpa;BODIPY (ТЬЩЋ)ЃКжЌЕЮШОЩЋ;DAPI (РЖЩЋ)ЃКЯИАћКЫШОЩЋЁЃFЃКЬиЖЈУцЛ§Фк(143 μm× 143 μm)ВЛЭЌжБОЖжЌЕЮЕФЗжВМБШР§ЭГМЦЭМЁЃXжсЪЧжЌЕЮжБОЖ(μm),YжсЮЊЬиЖЈУцЛ§ФкВЛЭЌжБОЖЕФжЌЕЮЫљеМЕФБШР§ЁЃ1~5 μmЃКжБОЖ1~5 μmЕФжЌЕЮ;5~10 μmЃКжБОЖ5~10 μmЕФжЌЕЮ;>10 μmЃКжБОЖ10 μmвдЩЯЕФжЌЕЮЁЃnsЃКP>0.05;*ЃКP<0.05;**ЃКP<0.01;***ЃКP<0.001ЁЃБъГпЃК25 μmЁЃ
Fig. 4Ppa deletion mutant exbibited smaller lipid droplets in the 3rd instar larvae
2.3 ГЌСПБэДяPpaдіЧПжЌЕЮДЂДцЕФФмСІ
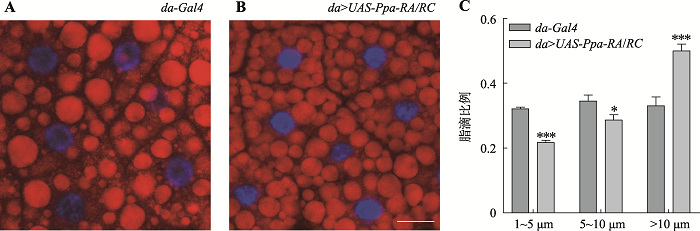
ЮЊСЫНјвЛВНбщжЄPpaдкжЌЗОДЂДцжаЕФзїгУ,БОбаОПМьВтСЫГЌСПБэДяPpaЖджЌЕЮЕФгАЯьЁЃЪЙгУNile redЖд3СфгзГцжЌЗОЬхжаЕФжЌЕЮНјааШОЩЋБъМЧ,ЗЂЯжгыЖдеезщda-Gal4ЯрБШ,ГЌСПБэДяPpa-RA/RCЕМжТжЌЕЮжБОЖУїЯддіМг,БэЯжЮЊДѓжЌЕЮЕФЪ§СПЯджјдіЖр(ЭМ5ЃКAКЭB)ЁЃеыЖджЌЕЮДѓаЁЕФВювь,ЭГМЦСЫЬиЖЈУцЛ§ФкВЛЭЌжБОЖжЌЕЮЕФЗжВМБШР§,гыЖдеезщЯрБШ,ГЌСПБэДяPpa-RA/RCБэЯжГіжБОЖ10 μmвдЩЯЕФДѓжЌЕЮБШР§УїЯддіМг,Жј1~5 μmЕФаЁжЌЕЮБШР§УїЯдМѕЩй(ЭМ5C),БэУїГЌСПБэДяPpaФмЙЛЕМжТжЌЗОЬхжааЁжЌЕЮМѕЩйКЭДѓжЌЕЮдіМгЁЃзлЩЯЫљЪі,БОбаОПЭЈЙ§бЧЯИАћЖЈЮЛЗжЮі,ЗЂЯжPpaЖЈЮЛгкЭйвКЯйгыжЌЗОЬхЕФЯИАћКЫжа;ЭЈЙ§PpaШБЪЇЭЛБфЬхЕФжЌЕЮаЮЬЌЗжЮі,ЗЂЯжжЌЕЮНЯЖдеезщБэЯжЮЊжБОЖБфаЁ;ЭЈЙ§ГЌСПБэДяPpa,ЗЂЯжжЌЗОЬхжаЕФжЌЕЮНЯЖдеезщБэЯжЮЊжБОЖдіМг,ВЂЧвЭЛБфЬхжаГЌСПБэДяPpaФмЙЛНЋЭЛБфЬхЕФжЌЕЮМѕаЁБэаЭЛиИДжСвАЩњаЭзДЬЌЁЃвдЩЯНсЙћБэУїPpaОпгаДйНјжЌЗОДЂДцЕФзїгУЁЃ
ЭМ5
 аТДАПкДђПЊ|ЯТдидЭМZIP|ЩњГЩPPT
аТДАПкДђПЊ|ЯТдидЭМZIP|ЩњГЩPPTЭМ5ГЌСПБэДяPpaЕМжТ3СфгзГцжаЕФжЌЕЮДѓаЁдіМг
AЁЂBЃК29ЁцЯТ,da-Gal4КЭda>UAS-Ppa-RA/RCЕФ3СфгзГцЕФжЌЕЮШОЩЋЭМЁЃNile red (КьЩЋ)ЃКжЌЕЮШОЩЋ;DAPI (РЖЩЋ)ЃКЯИАћКЫШОЩЋЁЃCЃКЬиЖЈУцЛ§Фк(143 μm×143 μm)ВЛЭЌжБОЖжЌЕЮЕФЗжВМБШР§ЭГМЦЭМЁЃXжсЪЧжЌЕЮжБОЖ(μm),YжсЮЊЬиЖЈУцЛ§ФкВЛЭЌжБОЖЕФжЌЕЮЫљеМЕФБШР§ЁЃ1~5 μmЃКжБОЖ1~5 μmЕФжЌЕЮ;5~10 μmЃКжБОЖ5~10 μmЕФжЌЕЮ;>10 μmЃКжБОЖ10 μmвдЩЯЕФжЌЕЮЁЃ*ЃКP<0.05;***ЃКP<0.001ЁЃБъГпЃК25 μmЁЃ
Fig. 5Overexpression of Ppa led to increased sizes of lipid droplets in the 3rd instar larvae
3 ЬжТл
дкаЁЪѓ(Mus musculus)жа,F-boxЛљвђМвзхГЩдБFbxw7ЕФШБЪЇв§ЗЂСЫИпбЊЬЧКЭвШЕКЫиЕжПЙ;ЭЌЪБ,дкаЁЪѓИЮдржаГЌСПБэДяFbxw7ВЛНіФмЙЛЯджјвжжЦИпжЌЮЙбјв§ЗЂЕФвШЕКЫиЕжПЙ,ЖјЧвПЩвдНЕЕЭЗЪХжаЁЪѓЕФИпбЊЬЧ[20,21];ШЫЬхЯИАћжмЦкЫивРРЕадМЄУИвжжЦЕААз(CDKIs)ГЩдБp27ЕФШБЪЇЕМжТжЌЗОЯИАћЪ§СПЕФдіМг,F-boxЕААзSЦкМЄУИSkp2ФмЙЛЪЖБ№p27ВЂЖдЦфНјааНЕНт;ВЂЧвдкSkp2ЕФЛљвђЧУГ§аЁЪѓжа,ЦЄЯТЯИАћКЭФкдрЕФжЌЗОЯИАћЪ§СПЯТНЕСЫ50%,БэУїSkp2ОпгаДйНјжЌЗОЕФзїгУ[22]ЁЃдкБОбаОПжа,бЁШЁСЫF-boxМвзхГЩдБPPAзїЮЊбаОПЖдЯѓ,ЗЂЯжPpaЦЌЖЮШБЪЇЭЛБфЬхБэЯжЮЊаЁжЌЕЮдіЖр,ЖјГЌСПБэДяPpaЕМжТДѓжЌЕЮдіМг,ВЂЧвЭЛБфЬхжаГЌСПБэДяPpaФмЙЛЛиИДЭЛБфЬхжЌЕЮМѕаЁЕФБэаЭ,етгыSkp2ЕФзїгУЪЧЯрЫЦЕФ,БэУїСЫF-boxМвзхЕААзШЗЪЕВЮгыСЫжЌжЪДњаЛЕїПиЁЃвЛАуЖјбд,F-boxЕААзЭЈЙ§ЗКЫи-ЕААзУИЬхЭООЖНщЕМАаЕААзЕФНЕНт,ФПЧАвбБЈЕРЕФPpaАаЕААзАќРЈзЊТМвђзгPrdКЭзХЫПСЃзщЕААзCID,ЫќУЧЕїПизХЬхНкЗЂг§КЭзХЫПСЃЖЈЮЛ[17,19],ЕЋЮДМћБЈЕРгыжЌЗОЯрЙиЕФАаЕААзЁЃвбгаЕФбаОПБэУї,дкHeLaЯИАћжаЧУГ§E3ЗКЫиСЌНгУИБрТыЛљвђMARCHF6ЕМжТжЌЕЮЗсЖШдіМг,MARCHF6ЕФЗКЫиЛЏЕзЮяАќРЈPerilipin2 (PLIN2)ЁЂЕЈЙЬДМзЊдЫЕААзNPC1КЭЕЈЙЬДМКЯГЩЯоЫйУИLDM,етаЉЕААзжБНгВЮгыжЌЕЮзщГЩЛђЕЈЙЬДМКЯГЩ [23,24]ЁЃДЫЭт,ЗКЫи-ЕААзУИЬхЭООЖВЮгыЕїПижЌжЪДњаЛЭЈТЗЕФЖрИіЙиМќУИЁЃдкаЁЪѓИЮдржа,жЌЗОИЪгЭШ§ѕЅжЌУИATGLЪЧвЛжжжївЊЕФИЪгЭШ§ѕЅЗжНтУИ,ФмЙЛНЋИЪгЭШ§ѕЅЗжНтГЩгЮРыжЌЗОЫсЁЃбаОПБэУї,E3ЗКЫиСЌНгУИCOP1АаЯђНЕНтATGL,Cop1ЧУГ§аЁЪѓЕФжЌЕЮЪ§СПЯджјМѕЩй,ЖјГЌСПБэДяCop1в§ЗЂаЁЪѓжЌЕЮЪ§СПЕФдіМг,БэУїCOP1ЕФЙІФмЪЧвжжЦжЌЗОЗжНт[25]ЁЃДЫЭт,дкФЬХЃШщЯйЩЯЦЄЯИАћжавжжЦЕААзУИЬхЛюадКѓ,ввѕЃИЈУИAєШЛЏУИACCЕФmRNAБэДяСПЯджјЯТЕї,ЕЋжЌЗОЫсКЯГЩУИFASNЕФmRNAБэДяСПЯджјЩЯЕї,ЫЕУїЗКЫиЕААзУИЬхЭООЖФмЙЛЕїПижЌЗОЫсКЯГЩ[26]ЁЃPpaзїЮЊSCFЗКЫиЛЏИДКЯЬхЕФзщЗж,ОпгаИЛССАБЫсжиИДађСа,ИУНсЙЙгђФмЙЛЪЖБ№ЕзЮяЕААз,ЭЦВтЦфЕзЮяЕААзПЩФмгыжЌжЪКЯГЩЛђепжЌЕЮНсЙЙаЮГЩЯрЙиЁЃвВгабаОПБЈЕР,F-boxЕААзПЩвдгыЦфЫћЕААзНсКЯаЮГЩИДКЯЬхЗЂЛгзїгУ,дкФ№ОЦНЭФИжа,F-boxЕААзCtf13гыSkp1ЁЂCep3аЮГЩКЫаФ,етИігЩ3жжЕААззщГЩЕФКЫаФгыNdc10НсКЯ,аЮГЩзХЫПСЃИДКЯЬхCBF3,ИУИДКЯЬхЖдЖЏСЃЕФзщзАжСЙиживЊ[9,27]ЁЃБОбаОПЗЂЯжPpaОпгаДйНјжЌЗОДЂДцЕФЙІФм,ЖјЦфЯТгЮЕФЕїПиЭјТчЛЙгаД§НјвЛВНбаОПЁЃ
(д№ШЮБрЮЏ: бжбд)
жТаЛЃК
ИааЛжаЙњПЦбЇдКвХДЋгыЗЂг§ЩњЮябЇбаОПЫљЛЦбЋбаОПдБЬсЙЉЕФЙћгЌЙЄОпжъКЭБэДядиЬх;ИааЛЙужнЛєЗђТќУтвпбаОПЫљНЙШЪНмНЬЪкКЭЫФДЈДѓбЇЛЊЮївНдКГТКЃбѓНЬЪкЬсЙЉЕФsgRNAдиЬхЙЙНЈКЭЛљвђЧУГ§ЗНАИЁЃВЮПМЮФЯз дЮФЫГађ
ЮФЯзФъЖШЕЙађ
ЮФжав§гУДЮЪ§ЕЙађ
БЛв§ЦкПЏгАЯьвђзг
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 2]
[БОЮФв§гУ: 2]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 2]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 2]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]
[БОЮФв§гУ: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}