
自驯化以来, 绵羊(Ovis aries)一直为人类提供肉, 毛, 皮, 奶和其它副产品,其对于生活在一些亚洲, 非洲和南美洲国家农业社区的数亿人的民生福祉至关重要。自大约10,000年前在中东由亚洲摩弗伦(Ovis orientalis)驯化以来, 绵羊扩张到亚洲其它地区以及欧洲, 非洲和新世界地缘。一个早期的重大事件是hair型绵羊驯化后向wool型绵羊的转变。
长期以来一直认为基因渗入是进化的强大力量, 基因组数据越来越多地提供证据表明基因渗入是遗传变异的一个至关重要的来源, 并且这种变异可用于动植物的适应性进化。绵羊属中也发生了从野生近缘种向家羊种群的基因渗入。因此, 绵羊通过适应各种环境和不同生产系统下的遗传改良, 分布到了全球范围的不同地域, 发展成为独特的品种。地方品种和改良品种的广泛变异是家养绵羊基因组变异, 适应性特征和重要农业相关性状的基础。
亚洲摩弗伦和乌里亚尔羊具有最高的核苷酸多样性, 而在大角羊中观察到的最低。亚洲摩弗伦和乌里亚尔羊在遗传上没有完全分离, 表明它们是扩大家养绵羊改良遗传资源的潜在资源。来自不同大洲地理区域的种群之间没有太大差异。家养绵羊的多样性在π值范围内变化很大, 从π = 3.1×10-3(中国新疆的巴什拜羊)到1.7×10-8(瑞典Sv?rdsj?品种)。所有家羊种群在约8,000-10,000 BP期间持续下降, 反映了驯化的瓶颈效应。它们随后的种群扩张显示了社会中农业的扩张。
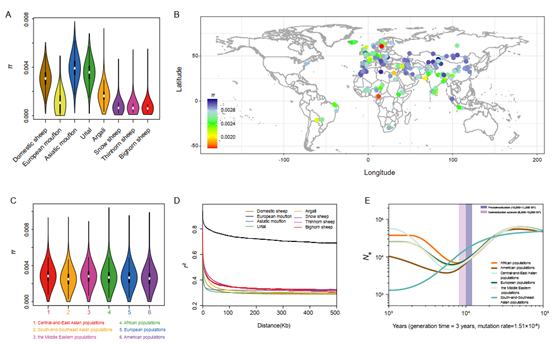
图1 遗传多样性(A)七种野生和家养绵羊的平均核苷酸多样性 (π)(B)全球158个家羊种群的π(C)六个地理区域家羊种群的π(D)七个野生近缘种和家羊的连锁不平衡衰减 (E)SMC++推断六个地理区域的家羊种群的Ne
家养绵羊分为6个地理结构的进化枝(中亚&东亚、南亚&东南亚、中东、非洲、欧洲和美洲种群),并有明显例外的现象出现:例如,瘦尾藏羊和中国西南羊(红色)接近印度品种(黄色);中东绵羊集群里的阿塞拜疆绵羊(红色);亚洲细毛品种(红色)集群接近欧洲美利奴羊。PCA在很大程度上将6个地理组分开, 韦桑羊(Ouessant)和哥特兰羊(Gotland)处于极端位置。欧洲, 非洲种群在很大程度上与亚洲种群分开, 也具有重叠(如美利奴羊衍生个体)。7种野羊存在于5个遗传簇(亚洲摩弗伦和乌里亚尔羊在同一遗传簇;大角羊和扁角羊在同一遗传簇)。家养绵羊含有6个地理分布的基因组成分。家羊品种有来自于野羊的遗传成分。欧洲、非洲和亚洲种群中遗传杂合性的明确证据, 表明在当地品种的培育过程中,来自不同大陆的优良遗传种群之间存在大量基因流。
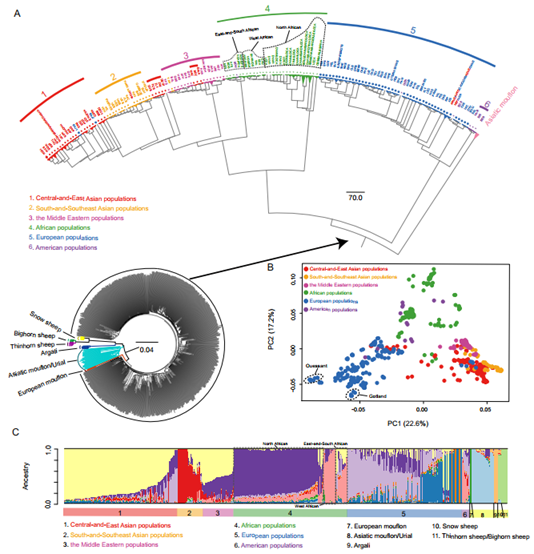
图2 系统发育关系(A)野生和家养绵羊的邻接树(B)主成分分析(C)种群遗传结构(K =11)
本研究探索了羊毛细度的遗传基础,针对于羊毛性状,划分为hair型(hair), 粗毛型(coarse-wool), 中等细度型(medium-wool), 细毛型(fine-wool)这四个表型,通过XP-CLR, π ratio, 等位基因频率分布, 单倍型模式, 不同物种序列的比较, 共同揭示了IRF2BP2基因的3'-UTR中的一个新突变(chr25: T7,068,586C)作为羊毛纤维直径的合理因果变异。对于羊毛进化过程(hair型→粗毛型→中等细度型→细毛型), 本研究发现一系列与羊毛性状相关的基因,例如VEGF信号通路, 转化生长因子β2产生的调节,毛囊发育 (IRF2BP2, VEGF, FGF7, HR, SMAD4, EDAR, FGFR2,和TP63),毛发生长方向 (ASTN2), β-catenin结合 (LEF1)等。

图3 hair型、粗毛型、中等细度型、细毛型的基因组进化(A) 家羊hair型和细毛型之间的XP-CLR测试 (B) π比值的6个比较(πhair型/π粗毛型、πhair型/π中等细度型、πhair型/π细毛型、π粗毛型/π中等细度型、π粗毛型/π细毛型和π中等细度型/π细毛型)在IRF2BP2基因中显示出强选择性信号 (C)IRF2BP2(chr25: T7,068,586C) 3'UTR中突变位点的等位基因频率和CC、TT和CT基因型频率,并测试基因型差异的显著性 (D)野羊、家养绵羊的hair型、粗毛型、中等细度型、细毛型种群中IRF2BP2的单倍型模式 (E)不同物种间在突变chr25: 7,068,586和chr25: 7,070,630处的序列比较 (F)GO和KEGG通路富集分析,显示了显著GO和通路以及相关基因
利用mRNA-seq, miRNA-seq发现IRF2BP2的mRNA表达的FPKM值显示粗毛型和细毛型绵羊在不同产前阶段的皮肤组织存在显著差异(P< 0.05),没有观察到粗毛羊IRF2BP2的表达。粗毛型湖羊和细毛型敖汉细毛羊的皮肤样本中的miRNA表达谱揭示了粗毛羊中仅oar-miR-20a-3p存在表达。功能验证将野生型IRF2BP2的3'-UTR引入荧光素酶表达载体,发现与阴性对照模拟物相比,oar-miR-20a-3p的表达降低了报告基因活性,但突变型IRF2BP2的3'-UTR消除了该进程(oar-miR-20a-3p介导的荧光素酶的减少)。野生型IRF2BP2的3'-UTR是oar-miR-20a-3p的直接功能靶点,oar-miR-20a-3p可以直接结合IRF2BP2并可能调节粗毛羊皮肤组织中IRF2BP2的表达。在H&E染色中观察到,与中国美利奴羊(细毛)相比,洼地绵羊(粗毛)的毛囊直径比例(DS/DP)更大,但次级毛囊和初级毛囊的数量比例(NS/NP)更小,表明细毛羊中均匀小直径纤维的产量更高,羊毛更细。oar-miR-20a-3p可能通过其对IRF2BP2的影响作用于粗毛羊毛囊的发育,可能会抑制毛囊中VEGFA信号的表达。简而言之,oar-miR-20a-3p→IRF2BP2→VEGFA是通过抑制粗毛羊IRF2BP2基因表达而导致抑制次级毛囊发育的潜在调控通路。
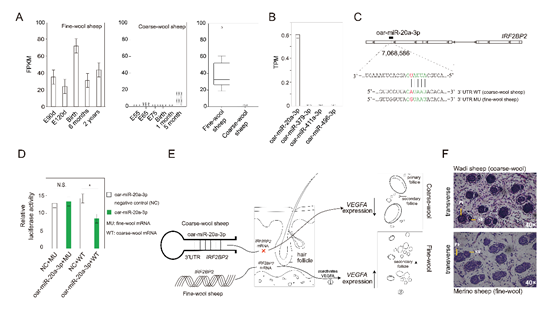
图4 试验验证(A,B)转录组和小RNA分析(C,D)双荧光素酶报告基因检测(E,F)家羊羊毛纤维变异相关基因IRF2BP2和oar-miR-20a-3p的遗传机制以及组织学观察
该研究完善了我们对绵羊基因组变异的理解。对遗传学家和育种人来说,本研究的绵羊属遗传变异种质资源是无价的。本研究的创新点在于: (1)全面揭示全球野羊和家羊的遗传多样性和系统发育关系,并且发现种群结构分化的一些例外情况,这些例外可以反应物种/品种的混合历史。(2)发现南亚&东南亚绵羊迁徙的历史与人类迁徙历史类似,为鲜少研究的南亚&东南亚人类历史提供佐证。(3)收集齐全所有野生近缘种,尽可能集齐全世界所有家羊品种, 利用高深度重测序序列展开分析,检测到了被基因渗入的具体品种及其野生供体,也表明渗入区段所调节的基因功能。(4)发现了新的选择信号, 尤其是对于绵羊矮小性状筛选到的信号和基因。(5)发现位于IRF2BP2基因 3'-UTR 中的一个新突变(chr25:T7,068,586C)作为羊毛纤维直径的合理因果变异,并表明oar-miR-20a-3p→IRF2BP2→VEGFA是抑制粗毛羊中IRF2BP2基因表达而抑制次级毛囊发育的潜在调控通路。
动物科技学院李孟华教授为论文通讯作者。中国农业大学吕锋骅副研究员,中国科学院动物研究所操银红博士,刘广建博士为论文共同第一作者。该工作得到了国家重点研发计划“政府间国际科技创新合作”重点专项,国家自然科学基金,中国科学院国际伙伴计划,第二次青藏高原综合科学考察研究(STEP)等项目的资助。伊朗、埃及、西班牙、德国、埃塞俄比亚、肯尼亚、英国、奥地利、冰岛、芬兰、加拿大、瑞典、意大利、巴西、尼泊尔、克罗地亚、捷克、挪威和荷兰等国科研人员参与了本项研究工作。
供稿:动物科学技术学院 操银红
供图:动物科学技术学院
编辑:马文哲
责编:于哲
