0 引言
【研究意义】红壤坡耕地是中国耕地资源的重要组成部分,该区水热资源丰富,是中国重要粮油生产基地[1]。近年来在降雨、地形等自然因素和耕作、施肥等人为因素的作用下,该区水土流失加剧,土地生产力下降,农业生态系统脆弱,严重制约了区域农业的可持续发展和生态环境的改善[2-3]。因此,探索、研究和实施有效的控制坡耕地水土流失技术,对实现我国资源、环境和人口的协调发展意义重大。【前人研究进展】等高植物篱和秸秆地面覆盖因其成本低、简单实用,生态、经济、社会效益显著,业已成为阻控南方红壤坡地水土流失和恢复重建侵蚀型红壤的有效措施[4-5]。近年来,植物篱和秸秆覆盖等水土保护措施在有效减流减沙,提高土壤养分,改善土壤团粒结构,增强土壤抗侵蚀能力等方面效果显著[6-11]。本研究团队之前研究发现,香根草篱种植5年后,土壤中>0.25 mm 粒级机械稳定性和水稳性团聚体含量均显著增加,香根草篱结合稻草覆盖增加了红壤坡耕地土壤及团聚体内有机碳含量[12]。有机碳对土壤团聚体的形成和稳定的重要作用已得到公认[13-14]。土壤微生物及其分泌的酶与土壤养分的生物转化过程紧密相关,是土壤有机碳转化过程的重要调控者,其在形成土壤肥力和稳定土壤团聚结构上有积极作用[15]。土壤团聚结构是制约土壤抗蚀性和抗冲性的重要因子[16]。因此,从水土保持的角度来讲,土壤微生物量及其结构变化一定程度上反映了水土保护措施阻控水土流失的机理和作用方式。【本研究切入点】目前,有关稻草覆盖、植物篱的研究多集中在其对土壤肥力、土壤结构状况和水土保持效益研究,对土壤微生物区系及多样性的研究较少,且大多集中在植物篱、覆盖等单一措施下[17-19],对两种措施联合作用下土壤微生物组成特征及其与土壤碳及碳转化相关土壤酶活性的关系研究鲜有报道。【拟解决的关键问题】本文采用始于2009年的红壤坡地不同生物水土保护措施的水土保护效果野外试验,利用磷脂脂肪酸方法(PLFA)研究香根草篱和稻草覆盖对红壤坡耕地土壤微生物数量及群落组成的影响,结合微生物生物量碳及土壤酶活性变化,探讨不同生物水土保护措施下土壤有机碳与土壤微生物种类及功能间的关系。旨在为探讨侵蚀红壤结构重建的途径,筛选更利于红壤坡地土壤结构改良和碳累积的合理管理模式提供理论依据。1 材料与方法
1.1 试验区概况
试验区位于江西省进贤县江西省红壤研究所水土保持试验站(116°20′24″E, 28°15′30″N)。该区属中亚热带季风气候,年均降雨量1 537 mm,年蒸发量1 100—1 200 mm,年均气温17.7—18.5℃。土壤为第四纪黏土母质发育的红壤旱地,供试土壤的基本理化性质如下:pH(H2O)5.0,有机质 16.2 g·kg-1,全氮 1.03 g·kg-1,全磷 0.66 g·kg-1,有效磷(Olsen-P)6.8 mg·kg-1,全钾 1.39 g·kg-1。 土壤容重1.27 g·cm-3,土壤总孔隙度58.4%,土壤砂粒、粉粒和黏粒的组成分别是16.31%、42.35%、41.35%。1.2 试验设计
该试验始于2009年,设置等高花生常规耕作(CK)、花生+香根草篱(H)、花生+稻草覆盖(M)、花生+香根草篱+稻草覆盖(HM)4个处理,3次重复,完全随机排列。各处理基础肥力及花生种植方式和农事操作相同。样地坡度10°,小区面积120 m2(24 m×5 m)。供试花生品种为粤油991,种植密度为32 cm×20 cm,花生等高种植,对照小区和秸秆覆盖小区种植72行,草篱小区和草篱+秸秆覆盖小区种植66行(其余6行为草篱)。香根草(Vetiveria zizanioides)篱每隔8 m双行种植,株行距为50 cm×50 cm,试验期间确保香根草定期刈割至30—50 cm。每隔3年施一次石灰,于土壤翻耕后均匀播撒石灰1 875 kg·hm-2,播种前施三元复合肥(N-P2O5-K2O: 15-15-15)416.7 kg·hm-2,钙镁磷肥(CaO≥18%,MgO≥3%,P2O5≥12%)525 kg·hm-2。覆盖稻草处理花生播种后每个小区均匀覆盖干稻草4 500 kg·hm-2。种植制度为一季花生。1.3 样品采集及测定方法
于2014年9月(花生收获后一个月)在试验小区的上坡位采集距离香根草篱带上部边缘2 m内耕层0—15 cm土样,无篱小区沿等高线采集相同坡位原状土样,每个土样为采样区内10个采样点的混合样。采集的土样剔除可见的动、植物残体和石块等过2 mm筛,在4℃下保存用于土壤微生物PLFA和土壤酶活性、微生物生物量碳和可溶性有机碳测定。取出部分样品风干后磨碎过100目筛,用于测定土壤有机碳含量。1.3.1 土壤酶活性和有机碳 选取了与土壤碳代谢有密切关系的β-葡(萄)糖苷酶(β-Glucosidase,β-Glu)、多酚氧化酶(polyphenol oxidase,PPO)、纤维素酶(cellulase,CEL)和蔗糖酶(invertase,INV),其酶活性的测定参考关松荫等[20]的方法,其中β-葡糖苷酶采用对硝基酚比色法,以1 g土壤在30℃下培养1 h生成对硝基酚的mg数表示;多酚氧化酶活性采用邻苯三酚显色法,以1 g土壤在30℃下培养2 h分解邻苯三酚的mg数来表示;纤维素酶采用3,5-二硝基水杨酸比色法,以10 g土壤在37℃下培养72 h后生成葡萄糖的mg数表示;蔗糖酶活性采用3,5-二硝基水杨酸比色法,以1 g土壤在37℃下培养24 h后生成葡萄糖的mg数表示。
土壤有机质测定采用浓硫酸-重铬酸钾外加热法;土壤可溶性有机碳采用蒸馏水25℃ 恒温振荡浸提30 min(水土比为4:1)后,以4 000 r/min离心10 min,上清液用0.45 µm 滤膜抽滤, 滤液直接在岛津TOC- VCPH有机碳分析仪上测定;土壤微生物生物量碳采用氯仿熏蒸-0.5 mol·L-1硫酸钾浸提法[21]测定。
1.3.2 土壤微生物中的磷脂脂肪酸(PLFA) 本试验采用修正的Bligh-Dyer[22]方法。(1)提取:称取8 g冷冻烘干土壤,用色谱纯CHCl3、CH3OH、磷酸缓冲液按 2:1:0.8的比例混合浸提液在<25℃下振荡2 h(285 r/min)后离心10 min(3 500 r/min),收集上层离心液,如此反复一次提取后加入磷缓冲液和氯仿各12 mL,于黑暗中静置12 h,吸取下层溶液,在30—32℃水浴锅中用氮气吹干。(2)分离:取1 000 μL CHCl3转移浓缩的磷脂到萃取小柱,向小柱加5mL CHCl3用于洗去中性脂,加2次5 mL丙酮用于洗去糖性脂,用1 mL甲醇清洗萃取小柱底部,收集5 mL甲醇淋洗硅胶小柱溶液,加入4 μL C19:0 内标,在32℃水浴锅中用氮气吹干。(3)甲脂化:用1 mL甲醇:甲苯混合液(体积比为1:1)和1 mL 0.2 mol·L-1KOH溶解干燥的磷脂样品,在37℃水浴中加热15 min,加入0.3 mL 0.2 mol·L-1的醋酸溶液,2 mL正己烷,2 mL超纯水,低速振荡(120 r/min)10 min,吸取上层正己烷溶液,氮气吹干,加入200μL正己烷冲洗试管,将溶液吸入小柱保存。用Agilent 6850气相色谱仪(FID检测器)测定,仪器条件:色谱柱型号为Agilent 19091B-102E Ultra 25% Phenyl Methyl Siloxane 25.0 m×200 μm×0.33 μm;进样量为2 μL;进样口温度为250℃;载气气体为氦气;恒压模式为22 psi;分流比为100:1。
1.4 数据处理与分析
用多属性决策法对酶活性进行归一化处理,先对不同处理的同一种酶进行归一化处理,用相同处理中归一化的几种酶的几何平均数代表这几种酶的总活性。计算公式为xi′= xi/$\sum^{4}_{i}X_i)$i=1,2,3,4)。式中,xi为单一酶活性的测定值;xi′为归一化的酶活性;i为不同处理编号。微生物生物量碳酶强度(MBCE)为归一化酶活性与微生物生物量碳的比值(酶单位/g)。试验结果通过SPSS 17.0 进行单因素方差分析(one-way ANOVA)和相关分析。多重比较选择Duncan极值法,显著性水平为5%。绘图采用Origin8.0软件。为保证结果可靠性,减少误差,仅分析含量高于0.1%的脂肪酸。
2 结果
2.1 土壤有机质和活性有机碳含量变化
土壤有机质含量研究结果表明,与试验前相比,香根草篱(H)和稻草覆盖(M)措施下土壤有机质含量维持在相对稳定水平,且有上升趋势,在二者联合时具有叠加效应,但CK处理降低了土壤总有机碳含量。与CK相比, H、M和HM 3种水土保护措施土壤总有机碳含量分别显著增加7.1%、6.5%和9.9%。但3种水土保护措施间土壤有机质含量无明显差异(图1)。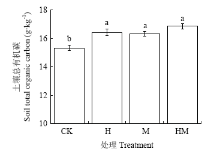 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1香根草篱和稻草覆盖对土壤总有机碳含量的影响图柱上不同小写字母代表处理间达5%差异显著,平均值±标准误差(n=3)。CK、H、M、HM分别表示花生常规耕作、花生+重根草篱、花生+稻草覆盖、花生+香根草篱+稻草覆盖。下同
-->Fig. 1The effect of Vetiver grass hedgerow and mulching on soil total organic carbon Different letters among the treatments mean significantly difference at 0.05 level. Mean ± SE (n = 3). CK, H, M, HM: Peanuts grown in high order, hedgerows+peanuts, mulching+peanuts, hedgerows+mulching+peanuts. The same as below
-->
土壤可溶性有机碳(DOC)和土壤微生物生物量碳(MBC)是土壤活性有机碳库的两个重要表征指标,对植物养分供应有最直接作用;微生物熵(微生物量碳与土壤总有机碳的比值,qMB)是评价土壤有机碳动态和质量的有效指标。从图2中可以看出,与CK相比,H、M和HM处理土壤中DOC含量分别提高15.1%、19.6%和13.6%, MBC含量分别提高100.7%、23.3%和160.0%,qMB分别提高87.4%、15.3%和135.9%,其提高幅度远远大于有机碳含量的提高幅度。双因素方差分析结果显示(表1),香根草篱和稻草等外源碳的投入均是增加土壤总有机碳、微生物量碳和微生物熵的主要影响因素。稻草覆盖对DOC含量的影响程度依赖于有无香根草篱种植。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2香根草篱和稻草覆盖下的土壤可溶性有机碳(A)、微生物生物量碳(B)含量和微生物熵(C)
-->Fig. 2Effects of vetiver grass hedgerow and mulching on soil dissolved organic carbon (A), microbial biomass carbon (B)and microbial quotient (C)
-->
2.2 土壤酶活性变化
由表2可知,除了多酚氧化酶(PPO)外,参与碳循环的蔗糖酶(INV)、纤维素酶(CEL)、β-葡糖苷酶(β-Glu)以及微生物生物量碳酶强度均受到香根草篱(H)的显著影响。INV和CEL的趋势一致,H和HM处理均能促进酶活性,分别是CK处理INV活性的1.51倍和1.53倍,CEL活性的1.61倍和1.44倍。与CK相比,H和HM处理均降低了土壤β-Glu活性和微生物生物量碳酶(MBCE)强度,单独香根草篱处理H对β-Glu的降低程度显著高于单独覆盖(M)处理。与CK相比,H和HM处理土壤β-Glu活性分别降低41.9%和73.1%。Table 1
表1
表1香根草篱和稻草覆盖对土壤有机碳、可溶性有机碳、微生物量碳和微生物熵影响的方差分析
Table 1ANOVAs results (F values and Probability level) on the effects of vetiver grass hedgerow(H) and mulching (M) on soil organic carbon, dissolved organic carbon, microbial biomass carbon and microbial quotient
| 变异来源 Sources of variation | 自由度 df | 有机碳 Organic carbon | 可溶性有机碳 Dissolved organic carbon | 微生物生物量碳 Microbial biomass carbon | 微生物熵 Microbial quotient |
|---|---|---|---|---|---|
| 香根草篱 H | 1 | 16.58** | 0.92 | 111.50** | 126.14** |
| 稻草覆盖 M | 1 | 12.97** | 3.55 | 13.51** | 12.10** |
| 交互 H×M | 1 | 2.01 | 4.87 | 2.57 | 3.21 |
新窗口打开
Table 2
表2
表2香根草篱和稻草覆盖对土壤酶活性的影响及方差分析
Table 2Effects of Vetiver grass hedgerow and mulching on soil enzymes activity and ANOVAs results
| 处理 Treatments | 蔗糖酶 Invertase (mg·g-1·24h-1) | 纤维素酶 Cellulase (mg·10g-1·72h-1) | 多酚氧化酶 polyphenol oxidase (mg·g-1·2h-1) | β-葡糖苷酶 β-Glucosidase (mg·g-1·h-1) | 微生物生物量 碳酶强度 Enzyme/MBC | 归一化酶 Normalized enzyme |
|---|---|---|---|---|---|---|
| CK | 86.21b | 74.44b | 0.17a | 14.48a | 0.0025a | 0.2259a |
| H | 130.04a | 120.12a | 0.19a | 8.42b | 0.0014b | 0.2542a |
| M | 93.84b | 84.27b | 0.25a | 13.34a | 0.0024a | 0.2550a |
| HM | 132.18a | 105.90a | 0.26a | 3.90c | 0.0010b | 0.2200a |
| 方差分析结果ANOVAs results (F values and probability level) | ||||||
| H | 27.75** | 42.16** | 0.31 | 52.95** | 33.99** | 0.063 |
| M | 0.39 | 0.18 | 4.35 | 7.07* | 1.64 | 0.038 |
| H×M | 0.12 | 5.38* | 0.03 | 2.51 | 0.63 | 5.660 |
新窗口打开
2.3 土壤微生物群落结构变化
2.3.1 土壤微生物磷脂脂肪酸(PLFA)含量变化及组成 PLFA 是活体微生物细胞膜的组分,特定的菌落PLFA数量变化可反映出原位土壤真菌(F)、细菌(B)活体生物量与菌落结构[23-24]。由表3可以看出,与CK相比,H、M和HM处理均增加了土壤微生物总PLFA(TB)的数量,不同水土保护措施下土壤各PLFAs含量具有明显差异,并且影响趋势和程度因微生物种类而不同(表3)。H处理的PLFAs 总量及细菌(B)、真菌(F)、革兰氏阳性菌(GP)、革兰氏阴性菌(GN)、放线菌(ACT)及AM真菌(AMF)含量均较高,且除了F和GN数量外,其他均显著高于CK处理;M处理主要增加了总PLFAs、B和ACT的PLFA量,较CK处理分别显著提高11.8%、13.4%和18.6%。Table 3
表3
表3不同水土保护措施下土壤微生物磷脂脂肪酸组成及含量(nmol·g-1)
Table 3Types and concentrations (nmol·g-1) of microbial PLFAs in soils with different soil and water conservation measures
| 处理 Treatment | ||||
|---|---|---|---|---|
| CK | H | M | HM | |
| 总磷脂脂肪酸 Total PLFAs | 26.39±1.30b | 29.75±0.31a | 29.51±0.82a | 29.18±0.29a |
| 细菌PLFA Bacterial PLFA | 10.81±0.43b | 12.78±0.44a | 12.26±0.33a | 12.23±0.45a |
| 真菌PLFA Fungi PLFA | 1.14±0.05a | 1.23±0.10a | 1.24±0.01a | 1.23±0.05a |
| 放线菌PLFA Actinomycetes PLFA | 2.15±0.13b | 2.63±0.06a | 2.55±0.08a | 2.67±0.09a |
| 革兰氏阳性菌PLFA GP PLFA | 5.78±0.42b | 7.02±0.30a | 6.79±0.26ab | 6.80±0.19ab |
| 革兰氏阴性菌PLFA GN PLFA | 4.71±0.23a | 5.37±0.23a | 5.12±0.25a | 4.92±0.32a |
| AM fungi PLFA | 0.46±0.04b | 0.65±0.04a | 0.61±0.05b | 0.64±0.03ab |
| 阴性PLFA/阳性PLFA GN PLFA/GP PLFA | 0.82±0.02a | 0.77±0.02ab | 0.75±0.01ab | 0.71±0.02b |
| 真菌PLFA/细菌PLFA F PLFA/B PLFA | 0.106±0.000a | 0.097±0.020a | 0.102±0.012a | 0.100±0.004a |
新窗口打开
真菌/细菌(F/B)及革兰氏阴性菌/革兰氏阳性菌(GN/GP)与微生物群落结构紧密相关,其中F/B比值可以表征农田土壤生态系统的稳定性[25]。由表3可以看出,不同水土保护措施间F/B无明显差异,说明稻草覆盖和香根草篱短期内对红壤坡耕地土壤生态系统稳定性无显著影响。稻草覆盖和香根草篱主要增加了土壤中GP菌的数量,进而减少了土壤中GN/GP比例,尤以HM处理减少幅度最大,较CK显著降低13.4%。
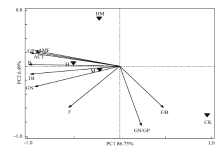
2.3.2 主成分分析 由PLFAs主成分分析可知(图3),第一主成分(PC1 )贡献率为86.75%,第二主成分(PC2)贡献率为6.49%,两者累积解释了微生物群落结构变化的93.24%。CK处理投射点分布在第四象限,H和HM处理投射点分布在第二象限,M处理投射点分布在第三象限,说明红壤坡耕地在稻草覆盖、香根草篱等水土保护措施下土壤磷脂脂肪酸组成结构与CK处理间产生较大的差异。不同水土保持措施处理中,H和M处理土壤微生物群落结构相近,与HM处理的磷脂脂肪酸的结构差别较大。由土壤PLFAs在主成分上的因子载荷分析表明(图3),对PC1贡献较大的为土壤总PLFAs生物量(TB)、B、AMF、ACT、GP和GN;对PC2贡献较大的为GN/GP和F/B。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3不同水土保护措施下土壤PLFA主成分分析
-->Fig. 3Principal component analysis of PLFAs in soils under different soil and water conservation measures
-->
2.4 各菌群PLFAs与土壤碳及参与碳循环的土壤酶活性的关系
冗余分析(RDA)表明,轴1和轴2解释了土壤菌落变异程度的95.37%。除了GN/GP和F/B外,土壤磷脂脂肪酸组成各指标之间及其和土壤MBC、TOC和DOC均呈正相关。在土壤酶活性指标中,土壤磷脂脂肪酸组成各指标与CEL和INV活性呈正相关,而与β-Glu活性及和碳循环酶有关的比例指标OCE和MBCE呈负相关。不同水土保护措施的影响差异明显,4个处理分别各占据1个象限,且H处理与除了β-Glu、OCE、MBCE、F/B和GN/GP外的所有指标呈正相关性(图4)。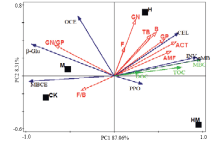 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4土壤微生物群落与土壤碳及参与碳循环酶活性的冗余分析排序图β-Glu:β-葡糖苷酶;INV:蔗糖酶;CEL:纤维素酶;PPO:多酚氧化酶;TOC:总有机碳;DOC:可溶性碳;MBC:微生物量碳;MBCE:单位微生物量碳酶活性;OCE:单位有机碳酶活性;GN:革兰氏阴性菌;GP:革兰氏阳性菌;TB:总生物量;B:细菌;F:真菌;ACT:放线菌;AMF:AM真菌
-->Fig. 4Redundancy analysis ordination showing the relationships among soil organic carbon, soil enzyme activity, and soil microbial communitiesβ-Glu: β-glucosidase; INV: Invertase; CEL: Cellulase; PPO: Polyphenol oxidase; TOC: Total organic carbon; DOC: Dissoved organic carbon; MBC: Microbial biomass carbon; MBCE: Microbial C-specific enzyme activities; OCE: Organic C-specific enzyme activities; GN: Gram negative bacteria; GP: Gram positive bacteria; TB: Total biomass; B: Bacteria;F: Fungi; ACT: Actinomycetes; AMF: AM fungi
-->
3 讨论
3.1 稻草覆盖和香根草篱对土壤碳及参与碳循环酶活性的影响
土壤有机碳组分及其含量是衡量土壤肥力的重要指标[26],农田系统中作物根系及其分泌物、根系残茬、有机肥投入和秸秆还田等均是增加土壤有机碳输入量的直接因素[12,27-28]。本研究中,与CK相比,H、M和HM外源碳投入均分别显著增加土壤总有机碳1.09、1.00和1.52 g·kg-1,另一方面,新鲜有机物的输入增加了土壤微生物活性,促进有机物的固定,进而增加有机碳的积累[29]。微生物生物量碳和可溶性有机碳是土壤有机碳中最活跃的组分之一,易被微生物分解利用、对植物养分供应有直接作用,是土壤潜在生产力的重要评价指标[30]。香根草篱和稻草覆盖通过提高土壤养分含量、改善土壤环境状况[31],为土壤微生物繁殖和生长提供了养料和水热条件,加速了土壤有机碳分解和转化,进而增加了土壤可溶性碳和微生物量碳的含量。相比于草篱根系的拦截作用,稻草覆盖在减弱雨滴对地表溅蚀的同时延缓了雨水在坡面地表的集聚速度和强度,进而减少了可溶性有机碳随雨水的淋失[32],以及稻草分解过程中产生大量水溶性有机碳均是其对增加土壤可溶性碳含量效果优于草篱的原因。相比稻草覆盖处理,草篱根际微生物群落结构的改变及其复杂的根系分泌物可能是其影响土壤可溶性有机碳含量改变的原因[33];同时,草篱处理因根系作用致使可溶性有机碳含量随雨水淋溶[34]以及本试验中为保证香根草篱长势,每年不定期刈割并移走,造成凋落物等可溶性有机碳的主要来源物减少是造成香根草篱有降低土壤可溶性有机碳含量趋势的又一原因。土壤酶活性是土壤生物活性和土壤肥力的重要指标,对土壤养分溶解、生物化学过程反应速率等都具有显著促进作用[35]。本试验中除了β-葡糖苷酶,其他3种参与碳循环的酶活性在不同水土保护措施下都有一定增加,主要是因为草篱和稻草覆盖直接或间接增加了土壤碳源,促进了微生物的繁殖,刺激了酶活性的增加,这与黄尚书等[36]对同一试验地的旱季土壤研究结果相似,但不同的是,本研究中香根草篱对4种参与碳循环的酶活性的影响显著,是因为本试验采集的是距离草篱2 m范围内的土壤样品。距离植物篱越近,植物篱对土壤理化性质及作物生长的影响越明显[12],同时,草篱发达的根系代谢也会释放大量的酶类[37]。H和HM处理降低了β-葡糖苷酶活性,这可能与各类酶的功能及来源有关,具体原因还有待进一步研究,但这也说明用单一酶活性评价土壤质量的局限性,土壤微生物数量的多少与土壤酶活性强弱不一定一致[38]。稻草覆盖对单一酶活性的影响不明显,但显著增加了单位微生物量酶活性(MBCE),说明了稻草覆盖条件下微生物对酶活性的贡献。3.2 稻草覆盖和香根草篱对土壤微生物各群落PLFA 含量的影响
本研究中,稻草覆盖、香根草篱和二者结合都增加了土壤中PLFAs总量,细菌、真菌和放线菌数量,此研究结果与前人研究结果一致[18]。土壤PLFAs主成分分析表明,H和M处理土壤微生物群落结构相似,而与HM处理的磷脂脂肪酸的结构差别较大,一方面说明香根草篱和稻草覆盖通过凋落物和根系带入了大量可被微生物分解利用的碳源和氮源,为微生物的繁殖提供了物质基础[16],而输入土壤的有机物抗分解能力较强,是造成土壤中真菌和放线菌丰富度较高的原因[39];另一方面也说明香根草篱和稻草覆盖相结合相比单一措施土壤理化性质的变异程度不同,影响了土壤微生物的数量及组成结构。AMF是土壤与植物根系关系最密切的微生物之一[40],广泛分布在根际土壤中,因此本研究中有草篱处理(H和HM)增加了AMF的数量。值得提出的是,主成分分析结果(图3)与各处理间单项指标单因素分析结果(表3)不一致的原因是因为主成分分析是将多项指标转化为少数几项综合指标,用综合指标来解释多变量的方差-协方差结构,是对原有变量的综合。不同种类微生物磷脂脂肪酸(PLFA)组成和含量间的差异,可用来直接评估其微生物的生物量及群落结构。真菌/细菌的比例是土壤生态系统缓冲能力的重要指标,比例越高则生态缓冲能力越高[25]。有研究表明秸秆还田显著提高了真菌/细菌比例[41],但本研究中真菌/细菌的比例并未有显著变化,可能是由于稻草覆盖和草篱等对细菌和真菌群落间丰度增加量相互抵消所致[41]。革兰氏阳性菌(GP)和革兰氏阴性菌(GN)是土壤细菌中重要组成部分,尤其对凋落物的分解起重要作用[15]。GN/GP值可以指示土壤营养状况,GN/GP越小,表示营养胁迫越强[42-43]。H、M及HM虽然都增加了GP和GN的数量,但GN/GP与CK相比呈下降趋势,且以HM处理GN/GP最小。H和M处理向土壤提供了丰富的有机碳源,但高碳氮比有机物的分解加剧了微生物和植物之间的养分竞争,因此H和M处理土壤微生物群落结构更倾向于向擅长竞争资源的革兰氏阳性细菌方向转变[44]。土壤理化性质的改变是影响微生物群落结构的主要原因,有机碳的种类和数量决定了微生物的种类和数量,并对微生物的生长繁殖及代谢过程产生影响。土壤酶作为土壤环境的重要组分,其与土壤微生物以及有机碳等理化性质之间存在密切的关系。而土壤微生物群落的动态变化反过来也影响着土壤系统的物质循环和能量流动[45]。冗余分析结果显示,土壤MBC、TOC,CEL、 INV、β-Glu活性及碳循环酶有关的比例指标OCE和MBCE均是影响土壤微生物数量和结构的主要因子。H处理中土壤MBC、TOC和CEL、 INV活性主要影响了B、ACT、AMF、GN、GP等微生物数量和总PLFAs量;M处理中β-Glu活性及OCE和MBCE主要影响了微生物结构指标GN/GP 和F/B,说明本实验中香根草篱和稻草覆盖由于不同密度、不同种类植物的凋落物和根系分泌物中输入碳的种类和数量不同[46-47],以及两种措施不同水土保护机制造成土壤养分及结构各异[12],进而影响土壤微生物的数量及组成结构,而香根草篱和稻草覆盖相结合加剧了与单一措施下土壤理化性质的变异程度,这也可能是导致HM处理土壤微生物结构不同于H和M处理的原因。同时,尽管PLFA技术常被用于研究土壤微生物群落结构,但该技术本身也有局限性。PLFA能定量描述环境样品中的微生物群体,而不能在种的水平精确的描述微生物的种类,且从土壤中提取出的各类脂肪酸种类仍十分有限[47],因此,要全面解析土壤微生物群落结构还需结合其他的研究手段。
4 结论
稻草覆盖和香根草篱通过秸秆、根系残茬和根系分泌物等直接或间接外源碳的投入均能显著增加土壤总有机碳含量及可溶性碳和微生物量碳等活性有机碳的含量。磷脂脂肪酸分析结果表明,香根草篱和稻草覆盖因不同密度、不同种类植物的凋落物和根系分泌物输入碳的种类和数量不同,以及两种措施不同水土保护机制造成土壤养分及结构各异,磷脂脂肪酸的种类及数量也会产生差异。同时,香根草篱和稻草覆盖降低了革兰氏阴性菌磷脂脂肪酸与革兰氏阳性菌磷脂脂肪酸的比值,且尤以二者结合处理显著。由此说明,香根草篱和稻草覆盖等生物水土保护措施在短期内是控制坡耕地水土流失,维持和提升耕地质量的有效措施,但长期应用要关注土壤碳氮养分均衡状态,以实现南方红壤坡耕地的可持续发展。(责任编辑 李云霞)
The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}