关键词:全球变暖; 玉米; 气孔特征; 气体交换; 华北平原 Responses of the Stomatal Traits and Gas Exchange of Maize Leaves to Climate Warming ZHENG Yun-Pu1,2,4, XU Ming2,*, WANG Jian-Shu3, QIU Shuai2, WANG He-Xin4 1 School of Water Conservancy and Hydropower, Hebei University of Engineering, Handan 056038, China
2Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographical Sciences and Natural Resources, Chinese Academy of Sciences, Beijing 100101, China
3 School of Agriculture, Hebei University of Engineering, Handan 056038, China
4Institute of Modern Agricultural Research, Dalian University, Dalian 116622, China
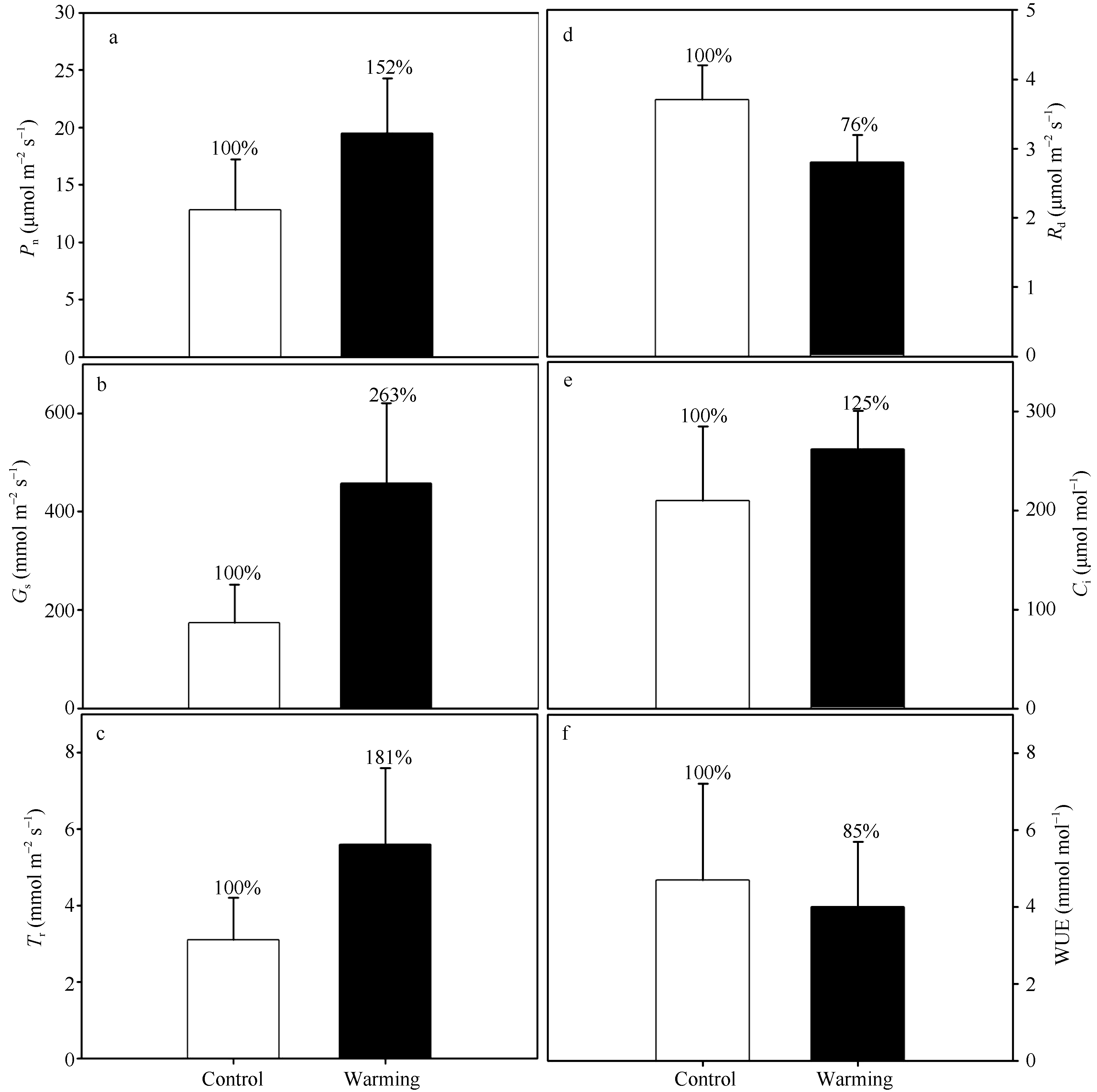
AbstractStomata are the pores on leaf surfaces controlling gas exchanges, mainly CO2 and water vapor, between the atmosphere and plants, and thus regulate carbon and water cycles in various ecosystems. This study investigated the effects of experimental warming on the stomatal frequency, stomatal aperture size and shape, and stomatal distribution pattern, and their relationships with the leaf gas exchange rates of maize ( Zea may L.) leaves through a field manipulative warming experiment with infrared heaters in a typical agriculture ecosystem in the North China Plain. Our results showed that experimental warming had little effect on stomatal density, but increased stomatal index by 12% ( P<0.05) due to the reduction in the number of epidermal cells under the warming treatment. Warming also decreased stomatal aperture length by 18% ( P<0.01) and increased stomatal aperture width 26% ( P<0.01). As a result, experimental warming increased the average stomatal aperture area by 31% ( P<0.01) and stomatal aperture circumference by 13% ( P<0.05), and resulted in a more regular stomatal distribution on both the adaxial and abaxial surfaces in leaves with an increased average nearest neighbor distance between stomata. In addition, experimental warming also affacted the gas exchange of maize leaves. Experimental warming significantly increased net photosynthetic rate ( Pn), stomatal conductance ( Gs), and transpiration rate ( Tr) by 52% ( P<0.05), 163% ( P<0.001), and 81% ( P<0.05), respectively. Meanwhile, experimental warming decreased the leaf dark respiration ( Rd) by 24% ( P<0.01), but had no significant effects on intercellular CO2 concentration ( Ci) and water use efficiency (WUE; P>0.05). In conclusion, the experimental warming may affect the gas exchange of maize leaves through the changes of the stomatal traits including stomatal frequency, stomatal aperture size and shape, and stomatal distribution on leaves.
Keyword:Global warming; Maize; Stomatal traits; Gas exchange; The North China Plain Show Figures Show Figures
表1 增温对玉米叶片气孔结构特征的影响 Table 1 Effects of experimental warming on stomatal traits of maize leaves
气孔参数 Stomatal parameter
对照Control
增温Warming
增量 Increase (%)
P值 P-value
近轴面 Adaxial
远轴面 Abaxial
近轴面 Adaxial
远轴面 Abaxial
气孔密度 SD (number mm-2)
56± 5 b
77± 3 a
58± 9 b
81± 1 a
—
—
67
70
4.5
P> 0.05
气孔指数 SI (%)
13.8± 0.1 c
19.4± 0.5 ab
16.5± 0.3 b
20.6± 0.8 a
—
—
16.6
18.6
11.7
P< 0.05
气孔长度 SAL (μ m)
36.8± 5.1 a
35.5± 2.1 a
30.8± 3.8 b
28.5± 3.7 b
—
—
36.2
29.7
-18.0
P< 0.01
气孔宽度 SAW (μ m)
3.7± 1.0 bc
3.2± 0.3 c
4.2± 1.0 ab
4.5± 0.9 a
—
—
3.5
4.4
25.7
P< 0.01
气孔面积 SAA (μ m2)
118± 43 bc
100± 15 c
150± 45 a
135± 31 ab
—
—
109
143
31
P< 0.01
气孔周长 SAC (μ m)
74± 10 b
70± 3 b
85± 10 a
77± 7 b
—
—
72
81
12.5
P< 0.05
气孔面积指数 SAI (%)
0.66± 0.24 c
0.77± 0.12 bc
0.87± 0.26 b
1.10± 0.25 a
—
—
0.71
0.99
39.9
P< 0.01
气孔形状指数 SASI (%)
13.8± 1.0 b
14.3± 0.7 b
14.4± 0.9 b
15.2± 0.9 a
—
—
14.0
14.8
5.7
P> 0.05
远/近轴面SD比率 Adaxial/abaxial
0.73± 0.09
0.72± 0.13
—
—
-1.4
P> 0.05
远/近轴面SI比率 Adaxial/abaxial
0.71± 0.09
0.80± 0.15
—
—
12.3
P< 0.05
Values are means ± standard deviation for SD, SI, SAAI, and SASI (75 subsamples and 3 replicates), and for SAL, SAW, SAA, and SAC (450 subsamples and 3 replicates). Means are compared by the one-way analysis of variance (ANOVA) at P< 0.05. Values followed by different letters are significantly different at P< 0.05 and the same letters indicate P> 0.05. SD: stomatal density; SI: stomatal index; SAL: stomatal aperture length; SAW: stomatal aperture width; SAA: stomatal aperture area; SAC: stomatal aperture circumference; SAI: stomatal area index; SASI: stomatal shape index. 所有数据为平均值± 标准偏差, 气孔密度、气孔指数、气孔面积指数、气孔形状指数为n = 75, 气孔长度、气孔宽度、气孔面积、气孔周长为n = 450。所有数据的平均值利用单因素方差分析在0.05水平上进行比较。同列标以不同字母的值在0.05水平上差异显著。SD: 气孔密度; SI: 气孔指数; SAL: 气孔长度; SAW: 气孔宽度; SAA: 气孔面积; SAC: 气孔周长; SAI: 气孔面积指数; SASI: 气孔形状指数。
表1 增温对玉米叶片气孔结构特征的影响 Table 1 Effects of experimental warming on stomatal traits of maize leaves
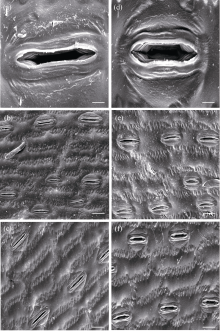
图3 玉米叶片气孔和表皮细胞的扫描电子显微照片 a、b、c为对照环境下的玉米叶片气孔特征, d、e、f为增温环境下的气孔特征。与对照相比, 增温使近轴面(b和e)和远轴面(c和f)的气孔均变得更短、更宽。同时, 增温还增加了表皮细胞的长度和宽度。标尺: 10 μ m (a和d)、40 μ m (b、c、e和f)。Fig. 3 Scanning electron microscopic photographs of stomata and epidermal cells in maize leaves Shorter and wider stomatal pores were observed on both the adaxial surface ( b and e) and abaxial surface (c and f) of maize leaves grown at elevated temperature than those grown at ambient temperature. In addition, elevated temperature also increased the width of epidermal cells. Bars, 10 μ m (a and d) and 40 μ m (b, c, e, and f).
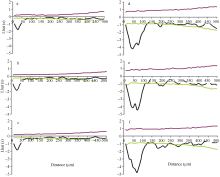
图4 玉米叶片近轴面气孔空间分布格局 对照环境下玉米叶片1、叶片2、叶片3近轴面(a~c)气孔的空间分布格局; 增温环境下玉米叶片1、叶片2、叶片3近轴面(d~f)气孔的空间分布格局。 上下两条包迹线代表95%的置信区间。Fig. 4 Spatial distribution pattern in the adaxial surface of maize leaves Point pattern analyses of stomata on the adaxial surface (a-c) in leaf 1, leaf 2, and leaf 3 of maize plants grown at ambient temperature and on those (d-f) at elevated temperature, respectively. The dotted lines give a 95% confidence envelope for complete spatial randomness. The data were given for three leaves from three ambient or warmed plots.
表2 增温对玉米叶片上不同位置气孔特征的影响 Table 2 Effects of warming on the stomatal traits among different sections of maize leaves
气孔参数 Stomatal parameter
气孔密度SD (Number mm-2)
气孔指数 SI (%)
气孔长度 SAL (mm)
气孔宽度 SAW (mm)
气孔面积 SAA (mm2)
气孔面积指数SAAI (%)
对照 Control
近轴面 Adaxal
上部Tip
51
13.4
38.9
6.9
112
0.57
中部Middle
61
14.0
49.8
6.8
117
0.71
下部Base
56
14.0
48.9
7.2
126
0.71
远轴面 Abaxial
上部Tip
73
19.2
38.7
5.8
84
0.61
中部Middle
79
19.7
49.0
6.6
97
0.77
下部Base
79
19.4
46.0
8.2
120
0.95
增温 Warming
近轴面 Adaxal
上部Tip
50
13.2
35.2
8.3
121
0.60
中部Middle
58
17.3
34.8
9.6
157
0.91
下部Base
67
19.3
34.8
8.2
168
1.14
远轴面 Abaxial
上部Tip
81
19.7
36.3
7.6
120
0.98
中部Middle
82
21.7
31.8
8.8
136
1.11
下部Base
80
20.6
31.1
9.6
156
1.25
温度 Temperature
< 0.050
< 0.001
< 0.001
< 0.001
< 0.001
< 0.001
叶面 Leaf surfaces
< 0.001
< 0.001
0.087
0.971
< 0.001
< 0.001
叶片部位 Leaf sections
< 0.001
< 0.001
< 0.010
< 0.010
< 0.001
< 0.001
温度× 叶面 Temperature × Leaf surfaces
0.536
0.059
0.587
0.714
0.330
< 0.050
温度× 部位 Temperature× Leaf sections
0.252
< 0.010
< 0.001
0.159
0.064
< 0.050
叶面× 部位 Leaf surfaces× Leaf sections
< 0.050
< 0.050
0.252
< 0.010
0.364
0.363
温度× 叶面× 部位 Temperature× surfaces× sections
< 0.050
0.068
0.630
0.661
0.086
< 0.001
Values are means ± standard deviation for SD, SI (25 subsamples and 3 replicates) and for SAL, SAW, and SAA (150 subsamples and 3 replicates). Means are compared by the ANOVA followed by Duncan’ s multiple range test. Values followed by different letters are significantly different at P< 0.05. Abbreviations are the same as those given in Table 1. 所有数据为平均值, 气孔密度和气孔指数均为n = 25, 气孔长度、气孔宽度及气孔面积为n = 150。所有数据的平均值利用多因素方差分析在0.05水平上进行比较, 对差异显著的组再进行多重比较(Duncan’ s multiple range test)。缩写同表1。
表2 增温对玉米叶片上不同位置气孔特征的影响 Table 2 Effects of warming on the stomatal traits among different sections of maize leaves
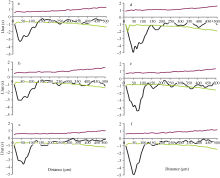
图5 玉米叶片远轴面气孔空间格局分析 对照环境下玉米叶片1、叶片2、叶片3远轴面(图4-a~c)气孔的空间格局; 增温环境下玉米叶片1、叶片2、叶片3远轴面(d~f)气孔的空间格局。 上、下2条包迹线代表95%的置信区间。Fig. 5 Spatial distribution pattern in the abaxial surface of maize leaves Point pattern analyses of stomata on the abaxial surface (a-c) in leaf 1, leaf 2, and leaf 3 of maize plants grown at ambient temperature and on those (d-f) at elevated temperature, respectively. The dotted lines give a 95% confidence envelope for complete spatial randomness. The data were given for three leaves from three ambient or warmed plots.
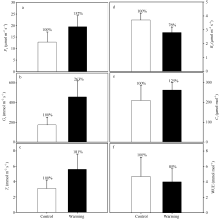
图6 实验增温对玉米叶片气体交换的影响 Pn: 净光合速率; Gs: 气孔导度; Tr: 蒸腾速率; Rd: 暗呼吸速率; Ci: 细胞间CO2浓度; WUE: 水分利用效率。Fig. 6 Effects of experimental warming on the gas exchange of maize leaves Pn: net photosynthetic rates; Gs: stomatal conductance; Tr: transpiration rates; Rd: dark respiration rates; Ci: intercellular CO2concentration; WUE: water use efficiency.
表3 Table 3 表3(Table 3)
表3 增温对玉米叶片表皮细胞和保卫细胞特征的影响 Table 3 Effects of experimental warming on the characteristics of epidermal and guard cells of maize leaves
参数 Parameter
对照 Control
增温 Warming
增量 Increase (%)
P值 P-value
近轴面Adaxial
远轴面Abaxial
近轴面Adaxial
远轴面Adaxial
表皮细胞Epidermal cells
表皮细胞密度 ECD (Number mm-2)
349± 27 a
321± 11 ab
296± 28 b
314± 16 ab
—
—
335
305
-9.0
P< 0.05
表皮细胞长度 ECL (µ m)
90.7± 5.4 a
74.5± 14.3 b
94.3± 19.0 a
82.6± 16.4 ab
—
—
81.7
89.3
9.3
P> 0.05
表皮细胞宽度 ECW (µ m)
24.0± 3.0 bc
21.9± 4.4 c
29.1± 4.4 a
25.6± 3.0 b
—
—
22.9
27.6
20.6
P< 0.01
表皮细胞面积 ECA (μ m2)
2104± 326 b
1496± 309 c
2638± 551 a
2051± 392 b
—
—
1766
2384
35.0
P< 0.01
表皮细胞周长 ECC (µ m)
247± 24 a
183± 26 b
293± 82 a
249± 54 a
—
—
212
274
29.6
P< 0.01
表皮细胞密度比率 Adaxial/abaxial
1.09
0.94
-13.3
P< 0.05
保卫细胞Guard cells
保卫细胞长度 GCL (μ m) #
52.4± 4.3 a
49.8± 4.0 b
47.1± 3.3 c
42.2± 2.6 d
—
—
51
45
-11.8
P< 0.05
保卫细胞宽度 GCW (μ m) #
6.2± 2.87 d
8.3± 1.24 c
11.6± 1.6 b
12.6± 1.2 a
—
—
7.3
12.1
66
P< 0.01
保卫细胞面积 GCA (μ m2)
453± 78 b
468± 77 ab
497± 70 ab
504± 72 a
—
—
461
501
8.0
P> 0.05
保卫细胞周长 GCC (μ m)
126± 11 a
129± 9 a
130± 15 a
134± 22 a
—
—
128
132
3.1
P> 0.05
Values are means ± standard deviation for ECD (75 subsamples, 3 replicates) and for other parameters (450 subsamples, 3 replicates). Means were compared by the ANOVA followed by Duncan’ s multiple range tests. Values followed by the different letters are significantly different at P< 0.05. #Guard cell length is the longest dimension, and the guard cell width is the widest dimension. ECD: epidermal cell density; ECL: epidermal cell length; ECW: epidermal cell width; ECA: epidermal cell area; ECC: epidermal cell circumference; GCL: guard cell length; GCW: guard cell width; GCA: guard cell area; GCC: guard cell circumference. 所有数据为平均值± 标准偏差, 表皮细胞密度为n = 75, 其他所有参数均为n = 450。所有数据的平均值利用单因素方差分析在0.05水平上进行比较, 对差异显著的组再进行多重比较(Duncan’ s multiple range test)。标以不同字母的值。#保卫细胞长度定义为保卫细胞长度方向上最长的距离, 保卫细胞宽度定义为保卫细胞宽度方向上最宽的距离。ECD: 表皮细胞密度; ECL: 表皮细胞长度; ECW: 表皮细胞宽度; ECA: 表皮细胞面积; ECC: 表皮细胞周长; GCL: 保卫细胞长度; GCW: 保卫细胞宽度; GCA: 保卫细胞面积; GCC: 保卫细胞周长。
表3 增温对玉米叶片表皮细胞和保卫细胞特征的影响 Table 3 Effects of experimental warming on the characteristics of epidermal and guard cells of maize leaves
4 结论未来全球气候变暖并不会对我国华北平原玉米叶片的气孔密度产生直接的影响, 但却可能通过改变表皮细胞的数目来影响气孔指数。同时, 未来气候变暖还会显著减小气孔开口的长度, 增加气孔开口的宽度, 从而增加气孔开口的面积和周长。另外, 全球气候变暖还可能使气孔在玉米叶片上的分布格局变得更加均匀。这些特征参数的改变导致华北平原玉米叶片气体交换过程显著变化, 从而可能最终影响到玉米的品质和产量。 The authors have declared that no competing interests exist.
Woodward FI. Stomatal numbers are sensitive to increases in CO2 from preindustrial levels. Nature, 1987, 327: 617-618[本文引用:3][JCR: 38.597]
[2]
Hetherington AM, Woodward FI. The role of stomata in sensing and driving environmental change. Nature, 2003, 424: 901-908[本文引用:3]
[3]
Franks PJ, Beerling DJ. Maximum leaf conductance driven by CO2 effects on stomatal size and density over geologic time. Proc Natl Acad Sci USA, 2009, 106: 10343-10347[本文引用:3][JCR: 9.737]
[4]
HaworthM, HeathJ, McElwain J C. Differences in the response sensitivity of stomatal index to atmospheric CO2 among four genera of Cupressaceae conifers. Ann Bot, 2010, 105: 411-418[本文引用:2]
[5]
Taylor SH, Franks PJ, Hulme SP. Photosynthetic pathway and ecological adaptation explain stomatal trait diversity amongst grasses. New Phytol, 2012, 193: 387-396[本文引用:2][JCR: 6.736]
[6]
Lake JA, Woodward FI. Response of stomatal numbers to CO2 and humidity: control by transpiration rate and abscisic acid. New Phytol, 2008, 179: 397-404[本文引用:1]
[7]
CiaisP, Denning AS, Tans PP. A three-dimensional synthesis of vegetation feedbacks in doubled CO2 climate experiments. J Geophys Res, 1997, 102: 5857-5872[本文引用:1][JCR: 3.021]
[8]
Buckley TN, Farquhar GD, Mott KA. Qualitative effects of patchy stomatal conductance distribution features on gas-exchange calculations. Plant Cell Environ, 1997, 20: 867-880[本文引用:1]
[9]
Apple ME, Olszyk DM, Ormrod DP. Morphology and stomatal function of douglas fir needles exposed to climate change: elevated CO2 and temperature. Int J Plant Sci, 2000, 161: 127-132[本文引用:3][JCR: 1.54]
[10]
Hovenden MJ. The influence of temperature and genotype on the growth and stomatal morphology of southern beech, Nothofagus cunninghamii (Nothofagaceae). Aust J Bot, 2001, 49: 427-434[本文引用:3][JCR: 1.204]
[11]
Kouwenberg L LR, Kürschner WM, McElwain JC. Stomatal frequency change over altitudinal gradients: Prospects for paleoaltimetry. Rev Mineral Geochem, 2007, 66: 215-241[本文引用:3][JCR: 3.119]
[12]
Fraser LH, GreenallA, CarlyleC. Adaptive phenotypic plasticity of Pseudoroegneria spicata: response of stomatal density, leaf area and biomass to changes in water supply and increased temperature. Ann Bot, 2009, 103: 769-775[本文引用:2][JCR: 0.657]
[13]
Beerling DJ, Chaloner WG. The impact of atmospheric CO2 and temperature change on stomatal density: observations from Quercus robur Lammas leaves. Ann Bot, 1993, 71: 231-235[本文引用:3][JCR: 0.657]
[14]
FerrisR, NijsI, BehaegheT. Elevated CO2 and temperature have different effects on leaf anatomy of perennial ryegrass in spring and summer. Ann Bot, 1996, 78: 489-497[本文引用:10][JCR: 0.657]
[15]
Reddy KR, Robana RR, Hodges HF. Interactions of CO2 enrichment and temperature on cotton growth and leaf characteristics. Environ Exp Bot, 1998, 39: 117-129[本文引用:5][JCR: 2.578]
[16]
Xu ZZ, Zhou GS, ShimizuH. Effects of soil drought with nocturnal warming on leaf stomatal traits and mesophyll cell ultrastructure of a perennial grass. Crop Sci, 2009, 49: 1843-1851[本文引用:5][JCR: 1.513]
[17]
Xu ZZ, Zhou GS. Effects of water stress and high nocturnal temperature on photosynthesis and nitrogen level of a perennial grass Leymus chinensis. Plant Soil, 2005, 269: 131-139[本文引用:3][JCR: 2.638]
[18]
张立荣, 牛海山, 汪诗平, 李英年, 赵新全. 增温与放牧对矮嵩草草甸4种植物气孔密度和气孔长度的影响. 生态学报, 2010, 30: 6961-6969Zhang LR, Niu HS, Wang SP, Li YN, Zhao XQ. Effects of temperature increase and grazing on stomatal density and length of four alpine Kobresia meadow species, Qinghai-Tibetan Plateau. Acta Ecol Sin, 2010, 30: 6961-6969 (in Chinese with English abstract)[本文引用:3]
[19]
Croxdale JL. Stomatal patterning in monocotyledons: Tradescantia as a model system. J Exp Bot, 1998, 49: 279-292[本文引用:4][JCR: 5.242]
[20]
BergerD, AltmannT. A subtilisin-like serine protease involved in the regulation of stomatal density and distribution in Arabidopsis thaliana. Genes Dev, 2000, 14: 1119-1131[本文引用:2][JCR: 12.444]
[21]
Croxdale JL. Stomatal patterning in angiosperms. Am J Bot, 2000, 87: 1069-1080[本文引用:3][JCR: 2.586]
[22]
Bergermann DC, LukowitzW, Somerville CR. Stomatal development and pattern controlled by a MAPKK Kinase. Science, 2004, 304: 1494-1497[本文引用:1]
[23]
JuarezM, TwiggR, TimmermansM. Specification of adaxial cell fate during maize leaf development. Development, 2004, 131: 4533-4544[本文引用:2][JCR: 6.208]
[24]
WangH, NgwenyamaN, LiuY. Stomatal development and patterning are regulated by environmentally responsive mitogen-actived protein kinases in Arabidopsis. Plant Cell, 2007, 19: 63-73[本文引用:1][JCR: 9.251]
[25]
Casson SA, Hetherington AM. Environmental regulation of stomatal development. Curr Opin Plant Biol, 2010, 13: 90-95[本文引用:1][JCR: 8.455]
[26]
Ciha AJ, Brun WA. Stomatal size and frequency in soybeans. Crop Sci, 1975, 15: 309-313[本文引用:2][JCR: 1.513]
[27]
Green RL, Beard JB, Casnoff DM. Leaf blade stomatal characterizations and evapotranspiration rates of 12 cool-season perennial grasses. HortScience, 1990, 25: 760-761[本文引用:2][JCR: 0.938]
[28]
Driscoll SP, PrinsA, OlmosE. Specification of adaxial and abaxial stomata, epidermal structure and photosynthesis to CO2 enrichment in maize leaves. J Exp Bot, 2006, 57: 381-390[本文引用:3][JCR: 5.242]
[29]
Salisbury EJ. On the causes and ecological significance of stomatal frequency, with special reference to the woodland flora. Philos Trans R Soc Lond B Biol Sci, 1927, 216: 1-65[本文引用:2]
[30]
Sharma GK, Dunn DB. Environmental modifications of leaf surface traits in Datura stramonium. Can J Bot, 1969, 47: 1211-1216[本文引用:2][JCR: 1.397]
[31]
TicháI. Photosynthetic characteristics during ontogenesis of leaves: 7. Stomata density and sizes. Photosynthetica, 1982, 16: 375-471[本文引用:2][JCR: 0.862]
[32]
ZacchiniM, MoriniS, VitaglianoC. Effect of photoperiod on some stomatal characteristics of in vitro cultured fruit tree shoots. Plant Cell Tissue Organ Cult, 1997, 49: 195-200[本文引用:2]
[33]
Stancato GC, Mazzoni-Viveiros S C, Luchi A E. Stomatal characteristics in different habitat forms of Brazilian species of Epidendrum (Orchidaceae). Nord J Bot, 1999, 19: 271-275[本文引用:2][JCR: 0.595]
[34]
TaoF, YokozawaM, XuY, HayashiY, ZhangZ. Climate changes and trends in phenology and yields of field crops in China, 1981-2000. Agric For Meteorol, 2006, 138: 82-92[本文引用:1]
[35]
LinE. Agricultural vulnerability and adaptation to global warming in China. Water Air Soil Pollut, 1996, 92: 63-73[本文引用:1][JCR: 1.748]
[36]
Mo XG, Liu SX, Lin ZH, Guo RP. Regional crop yield, water consumption and water use efficiency and their responses to climate change in the North China Plain. Agric Ecosyst Environ, 2009, 134: 67-78[本文引用:1]
[37]
Liu SX, Mo XG, Lin ZH, Xu YQ, Ji JJ, WenG, RicheyJ. Crop yield responses to climate change in the Huang-Huai-Hai Plain of China. Agric Water Manag, 2010, 97: 1195-1209[本文引用:2]
[38]
Tao FL, ZhangS, ZhangZ. Spatiotemporal changes of wheat phenology in China under the effects of temperature, day length and cultivar thermal characteristics. Eur J Agron, 2012, 43: 201-212[本文引用:2][JCR: 2.8]
[39]
Hou RX, Ou-YangZ, Li YS. Is the change of winter wheat yield under warming caused by shortened reproductive period?Ecol Evol, 2012, 2: 2999-3008[本文引用:2][JCR: 1.184]
[40]
Lobell DB, Field CB. Global scale climate-crop yield relationships and the impacts of recent warming. Environ Res Lett, 2007, 2: 1-7[本文引用:1][JCR: 3.582]
[41]
Ripley BD. The second-order analysis of stationary processes. J Appl Prob, 1976, 13: 255-266[本文引用:1][CJCR: 0.2331]
[42]
Lomax BH, Woodward FI, Leitch IJ. Genome size as a predictor of guard cell length in Arabidopsis thaliana is independent of environmental conditions. New Phytol, 2009, 181: 311-314[本文引用:1][JCR: 6.736]
[43]
DjanaguiramanM, Prasad P V V, Boyle D L. High-temperature stress and soybean leaves: leaf anatomy and photosynthesis. Crop Sci, 2011, 51: 2125-2131[本文引用:1][JCR: 1.513]
 , 王建书
, 王建书


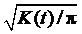





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}