 , 李伟
, 李伟*通讯作者(Corresponding author): 申家恒, E-mail:hsdshenjiaheng@163.com, Tel: 0535-6685930 收稿日期:2014-04-16 基金:
摘要
关键词:花生(
Double Fertilization and Duration of Phases in Peanut (
SHI Chun-Yan, SHEN Jia-Heng
, LI WeiFund:
Abstract
Keyword:
Show Figures
Show Figures
花生是豆科中重要的油料作物之一, 种子含油量约45%[ 1]。落花生通称花生, 是植物界被子植物门, 双子叶植物纲, 豆科, 蝶形花亚科, 落花生属, 落花生种。一年生草本, 根部有丰富的根瘤, 茎直立或匍匐, 长30~80 cm, 原产巴西。16世纪初叶或中叶, 由华侨引进福建、广东、山东等地区种植。我国花生年产量1500万吨以上, 居世界首位。花生种子内富含油脂、蛋白质和淀粉, 除食用外, 还有药用, 亦是制造生发油等化妆品的原料[ 2]。关于花生的大小孢子的发生, 雌雄配子体的发育以及胚胎的发育[ 3, 4, 5]均有研究, 对花生的受精过程国内外报道较少, 邢树平等[ 6]、Simth等[ 7]和何远康等[ 8]作了相关报道, 个别在时间上差异较大, 大部分结果差异不大。为进一步确定花生双受精过程及其经历的时间, 本研究以生长在哈尔滨地区的四粒红花生为试材, 以大量制片为基础, 将花生受精过程中雌雄性细胞融合的形态变化与相应经历的时间相结合, 获得了受精过程中更加详细准确的形态学信息, 确定受精过程经历的时间表, 丰富了花生胚胎学资料, 对花生育种、转基因操作等具有重要意义。
1 材料与方法1.1 试验材料落花生( Arachis hypogaea L .)“四粒红”种植于哈尔滨师范大学植物园, 普通大田管理, 生长良好。
1.2 试验方法从2010年到2012年每年的6月末至7月末取材, 取材时田间昼夜平均温度为20~30℃。早晨5:30左右花大量开放, 开花后6 h内, 每2 h取一次柱头和花柱, 开花后7~ 20 h, 每1 h取一次子房, 开花后20~36 h, 每2 h取一次子房, 开花后36~48 h, 每4 h取一次子房, 每次取50至100个试材。3年3个重复。
将材料放入卡诺固定液(无水乙醇与冰醋酸体积比为3∶1)固定2 h, 之后转入70%乙醇中保存在4℃冰箱中备用。爱氏苏木精染液(爱氏苏木精原液与45%醋酸体积比为1∶1)整体染色3~5 d。常规石蜡切片法制片, 切片厚度8 μm, 用Nikon ECLIPSE E600W型显微镜观察并摄影。实体解剖镜下观察花生的散粉。
2 结果与分析2.1 花粉的萌发及花粉管的生长2.1.1 精子的形成 花生的花朵由花萼、花瓣、雄蕊和雌蕊组成, 为上位花。闭花自花授粉, 开花前2~4 h就已经散粉(图版I-1)。花粉为二细胞型。花粉粒在柱头上萌发长出花粉管(图版I-2, 3, 4), 花粉管穿过柱头的表皮与薄壁组织的细胞间隙进入花柱道, 沿花柱道的表面向子房方向生长。大约开花1 h后在花粉管中可观察到蠕虫形的生殖细胞, 生殖细胞正处于分裂前期(图版I-5)。而后可观察到分裂中期(图版I-6), 此时染色体清晰可见。分裂后期(图版I-7)染色体平均分配到2个子细胞中。分裂末期时(图版I-8)在花粉管的挤压下精子呈椭球形, 2个精子保持一定的距离继续向子房方向移动。开花后5~6 h花粉管进入子房腔, 花粉管内的精子逐渐呈卵球形(图版I-9)。
2.1.2 精子的释放 花粉管到达子房腔后沿着胎座一侧向下延伸(图版I-9)到达胚珠。胚珠为双珠被, 蓼形胚囊。珠心细胞已经退化, 个别珠心残存着退化的痕迹, 有的珠孔端可以观察到少数珠心冠细胞。成熟胚囊的珠孔端由2个助细胞和一个卵细胞构成卵器(图版I-10, 11)。卵细胞呈近梨形, 卵细胞细胞核和细胞质被液泡挤到合点端, 液泡位于珠孔端, 占据卵细胞的大部分体积。此时反足细胞已经退化。助细胞大部分完好, 极少数有退化的迹象, 染色稍深, 有的2个助细胞都退化(图版I-12)。中央细胞有2个极核, 有的上下排列, 有的左右排列(图版I-13, 14), 有的2个极核融合成次生核(图版I-15)。花粉管从胚珠的珠孔进入胚囊。开花后8~9 h, 极核或次生核向卵细胞的合点端靠近; 花粉管的内容物释放到退化的助细胞中, 最初可见精子的细胞质, 很快脱去细胞质成为精核(图版I-16, 17)
2.2 精核与卵细胞融合开花后9 h可见精核与卵细胞、极核或次生核接触。当一个精核与极核或次生核融合时另外一个精核还贴在卵细胞膜上(图版II-18)。通常精核向卵细胞移动的速度慢于精核向极核及次生核移动的速度。
开花后10 h精核大部分贴合在卵核上(图版II-19), 与卵核渐渐融合(图版II-20, 21)。到开花后11 h, 精核在卵核上呈凸透镜形 (图版II-22)。开花后12 h精核染色质分散到卵核中 (图版II-23), 有雄性核仁出现(图版II-24)。此时作为雌雄性核融合结束的标志, 进入合子期。开花后13 h, 雄性核仁逐渐增大, 开花后15 h大量的合子中雄性核仁与雌性核仁等大, 成为2个等大核仁的合子(图版II-25)。雌雄核仁保持这样的状态可达开花后26~30 h, 而后两核仁开始靠近, 融合(图版II-26), 开花后约30 h两核仁融合成一个大核仁(图版II-27), 此时初生胚乳核已经分裂为2个胚乳游离核。
2.3 合子分裂合子的细胞质浓厚, 液泡消失。开花后约32 h合子中出现染色体 , 进入分裂的前期(图版II-28)。之后染色体横向(图版II-29)或纵向(图版II-30)排列在赤道板上, 横向排列居多, 进入合子分裂中期。开花后32~34 h进入合子分裂的后期, 染色体被平均分配到2个子细胞中, 出现新的核膜, 进入分裂末期(图版II-31, 32), 形成二细胞原胚(图版II-33), 合子的休眠期19 h。此时胚囊中有2~4个胚乳游离核。
2.4 精核与极核或次生核融合精核与极核或次生核的融合和精核与卵细胞融合基本上相似, 只是精核与极核或次生核接触早于精卵核的接触, 融合进程较快。开花后9 h精核附在极核或次生核上(图版III-34, 35)。开花后10 h精核紧紧贴附在极核及次生核上(图版III-36, 37), 精核呈透镜形。开花后11 h雌雄核的核膜进一步融合(图版III-38), 精核染色质完全分散到极核或次生核中并出现小的雄性核仁(图版III-39, 40)。此时作为极核或次生核与精核融合结束的标志, 初生胚乳核形成。初生胚乳核分裂前, 初生胚乳核内的雌雄性核仁存在3种情况, 其一为雌雄性核仁融合为1个较大核仁, 为三核合并的结果; 其二为1个大核仁与1个小核仁为次生核仁和雄性核仁; 其三为3个核仁同时存在, 为1个雄性核仁与2个雌性核仁。
2.5 初生胚乳核分裂初生胚乳核几乎没有休眠期, 或极短, 开花后13 h已经看到初生胚乳核的分裂相。开花后15 h分裂相比较集中, 分裂前期像(图版III-41)明显可见, 分裂的中期像位于胚囊的珠孔端, 有的在胚囊中央(图版III-42), 而后是分裂的后期(图版III-43)和末期像(图版III-44, 45), 进而形成2个胚乳游离核(图版III-46)。形成的胚乳游离核中有3个核仁的、2个核仁的和1个核仁的, 随后胚乳游离核进行下一次分裂。花生胚乳发育类型属于核型胚乳。
2.6 单受精和多精入胚囊现象开花后36 h的一个胚囊中观察到卵细胞受精后, 分裂成二细胞原胚, 没有受精的2个极核位于二细胞原胚的下方, 这个现象属于单受精(图版III-47); 此胚囊中还观察到多余花粉管的进入, 细胞质中存在2个精核(图版III-48, 49); 开花后36 h仍然有多余精子进入胚囊均属于多精入胚囊, 但没有观察到多精受精现象。
3 讨论3.1 精子的形成关于花粉管内生殖细胞的分裂Maheshwari将其总结为3个问题, 即是否有纺锤体形成; 有无中期板的形成; 是通过形成细胞板还是经细胞质的缢缩来完成胞质分裂[ 9]。邢树平等[ 6]通过半薄切片观察到花生花柱中生殖细胞分裂之后的2个精子, 观察到精子的细胞核和细胞质, 没有观察到生殖细胞的分裂过程。本试验通过石蜡切片可以清楚地观察到生殖细胞, 但看不到核仁, 分不清细胞质。可以观察到生殖细胞分裂成2个精子的前、中、后、末时期, 整个过程没有看到纺锤丝形成的纺锤体, 也没有观察到中期板。2个精子形成之后, 没有观察到细胞壁, 所以推测是由细胞质缢缩完成细胞分裂的。本试验表明花粉管在退化助细胞释放时精核周围有亮区。大豆精子被释放之后也可以看到透明区, 人们认为是精子的细胞质[ 10]。花生精子是否存在细胞质还有待进一步研究。
3.2 受精过程及其经历时间的确定四粒红花生的子房中通常有4个胚珠, 据观察胚珠受精的顺序是从上到下, 不是同时受精, 而且最下面的胚珠受精率较低, 所以本试验是按照最上面的胚珠受精时间确定的, 观察了将近3000个子房, 经过统计得到花生双受精过程各阶段经历的时间表(表1)。
表1
Table 1
表1(Table 1)
| 表1 花生双受精过程各阶段经历时间表 Table 1 Duration of double fertilization process in different stages in Arachis hypogea L. |
花生散粉大约在开花前的2~4 h, 这与邢树平等[ 6]的观察相同, 花粉随即萌发。开花后8 h左右, 花粉管已经进入胚囊释放内容物。中央细胞先于卵细胞受精, 但相差时间甚短, 以出现雄性核仁为受精结束的标志, 从开花后9 h融合开始到开花后13 h合子中出现雄性核仁为止受精结束, 开花后32 h合子分裂, 故合子的休眠期为19 h左右。极核及次生核在开花后9 h时与精核接触, 开花后13 h时进一步融合, 到开花后13 h时雄核仁在极核及次生核中出现, 形成初生胚乳核。开花后15 h时初生胚乳核大量分裂。故初生胚乳核的休眠期极短, 在0~2 h左右。有关花生雌雄性细胞融合发生的时间, 不同研究人员得出不同的结果, 根据邢树平等[ 6]的观察认为开花后12~20 h为雌雄配子融合阶段; 何远康等[ 8]认为如果早晨6:00至7:00时开花, 直到晚23:00时左右才进行受精作用; Smith等[ 7]认为授粉后10~18 h为双受精时间, 授粉后26 h合子分裂; 而“花生育种学”[ 11]与“作物栽培学”[ 12]中所记载的是授粉后12~18 h为受精时间。本试验表明, 开花后9~13 h为雌雄配子融合的时间, 与上述结果存在一定的差异。差异产生的原因, 很可能与研究者所用的花生品种、栽培环境、栽培条件以及生长温度相关。
3.3 单受精和多精入胚囊本试验在近3000个子房的切片中观察到一例单受精图像, 卵细胞受精并且已经横分裂成二细胞原胚, 而中央极核没有受精。此外还看到多例多精子进入胚囊的现象。有关文献[ 6, 7, 13]曾指出花生存在单受精现象, 但从未给出相应的照片。我们估计花生因其极核没有受精难以产生胚乳结构, 卵细胞受精并分裂产生的胚也很难发育成熟, 最终该胚珠可能败育。
3.4 合子休眠期的确定大量的研究表明无论是体内受精, 还是离体受精, 雌雄性配子融合(即受精作用)均发生在一个很短时间内, 而合子形成后, 大约经历较长休眠期才分裂。这其中涉及雌雄性配子融合结束的形态学标志是什么?早期的研究认为雌雄性配子融合后出现雄性核仁, 当雌雄性核仁融合为一个大核仁时, 标志着受精作用的结束, 邢树平等[ 6]报道即按此标准对花生雌雄配子融合的时间进行划分, 他认为开花后12~20 h为受精作用时间, 开花后21~26 h为合子休眠期(合子休眠期为5 h), 开花后27 h之后合子进行第一次分裂。申家恒等[ 14]在“玉米双受精过程的细胞学观察”一文提出, 以精核染色质均匀分散于雌性核内, 并出现已增大的雄性核仁作为受精终结的标志。本研究表明花生的雌雄配子融合并出现雄性核仁发生在开花后9~13 h, 而合子分裂则发生于开花后32 h, 故花生合子休眠期应为19 h, 对合子以及初生胚乳核的研究表明, 雄性核融入雌性核, 随着雄性染色质的分散, 出现一个小的雄性核仁, 继续增大雄性核仁的体积经历较长时间, 一旦雌雄性核仁融合, 预示该细胞即将分裂。
图版I、II和III中的缩写
Abbreviations in Plates I, II, and III
| Figure OptionViewDownloadNew Window | |
 | 图版I、II和III中的缩写 图版I、II和III中的缩写DSY: 退化助细胞; EC: 卵细胞; PN: 极核; PT: 花粉管; SY: 助细胞; ZY: 合子; SP: 精子; SN: 精核; SEN: 次生核; MN: 雄性核仁; PEN: 初生胚乳核; GC: 生殖细胞; TCP: 二细胞原胚; PSY: 宿存助细胞; FNE: 胚乳游离核; VN: 营养核; MP: 有丝分裂前期; MM: 有丝分裂中期; MA: 有丝分裂后期; MT: 有丝分裂末期。Abbreviations in Plates I, II, and III DSY: degenerated synergid; EC: egg cell; PN: polar nucleus; PT: pollen tude; SY: synergid; ZY: zygote; SP: sperm; SN: sperm nucleus; SEN: secondary endosperm nucleus; MN: male nucleolus; PEN: primary endosperm nucleus; GC: generative cell; TCP: two-cell proembryo; PSY: persistent synergid; FNE: the free nuclei of the endosperm; VN: vegetative nucleus; MP: mitosis prophase; MM: mitosis metaphase; MA: mitosis anaphase; MT: mitosis telophase. |
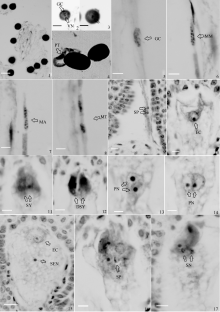
图版I
Plate I
| Figure OptionViewDownloadNew Window | |
 | 图版I 1: 花粉粒落在柱头上; 2: 成熟花粉, 箭头示生殖细胞和营养核; 3: 箭头示放大的成熟花粉营养核; 4: 花粉萌发, 长出花粉管(×800); 5: 生殖细胞进入花粉管, 在花粉管中向子房移动; 6: 生殖细胞分裂中期, 染色体清晰可见; 7: 分裂后期, 染色体平均分配; 8: 分裂末期, 形成2个子细胞; 9: 进入子房腔的2个精子; 10: 成熟的卵细胞形态; 11: 没有退化的2个助细胞; 12: 退化的助细胞, 染色较深; 13: 上下排列的2个极核; 14: 左右排列的2个极核; 15: 卵细胞, 次生核; 16: 精子在一个助细胞中释放; 17: 精核在卵细胞周围。(Bar=100 μm)Plate I Plate I 1: pollen grains on the stigma; 2: mature pollen, generative cell (GC) and vegetative nucleus (VN) (shown by arrow); 3: amplified vegetative nucleus of mature pollen (arrow); 4: pollen germination (×800), stretching from the pollen tube (PT); 5: generative cell coming into pollen tube and moving to the ovary in the pollen tube; 6: reproductive cell in mitosis metaphase (MM), pollen is clearly visible; 7: mitosis anaphase (MA), chromosome evenly distributed; 8: mitosis telophase (MT) forming two cells; 9: two sperms coming into ovary cavity; 10: egg cell(EC) shape; 11: two synergids (SY); 12: degenerated synergid (DSY); 13: two polar nucleus (PN) lined up and down; 14: two polar nucleus lined right and left; 15: egg cell, secondary nuclear; 16: sperms (SP) released; 17: sperm nucleus (SN) near the egg cell. (Bar=100 μm) |
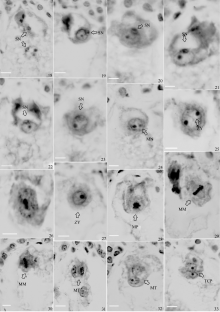
图版II
Plate II
| Figure OptionViewDownloadNew Window | |
 | 图版II 18: 精核分别贴在卵细胞和极核上;19: 精核和卵核贴合; 20~22: 精核与卵核渐渐融合; 23: 精核的染色质分散到卵核中; 24: 卵核中出现小的雄性核仁; 25: 2个等大核仁的合子, 雄性核仁渐渐长大, 和卵核仁等大; 26: 雌雄核仁靠近相融; 27: 雌雄2个核仁融合成一个核仁; 28: 合子分裂前期, 出现染色体, 核膜, 核仁消失; 29: 合子分裂中期(横分裂); 30: 合子分裂中期(纵分裂); 31~32:合子分裂末期; 33: 二细胞原胚。(Bar=100 μm)Plate II 18: sperm nucleus sticking to egg cell and polar nucleus; 19: sperm nucleus sticking to egg nucleus; 20-22: sperm nucleus and egg nucleus fused gradually; 23: sperm nucleus chromatin dispersed in egg nucleus; 24:small male nucleolus(MN) appeared in egg nucleus; 25: two same size and small nucleolus zygote; 26: female and male nucleolus close to fusing; 27: one nucleolus zygote fused from two nucleolus; 28: zygote mitosis prophase, appearing chromosome, disappearing nuclear membrane and nucleolus; 29: zygote mitosis metaphase(transverse division); 30: zygote mitosis metaphase (longitudinal division); 31-32: zygote mitosis telophase; 33: two-cell prombryo (TCP). (Bar=100 μm) |
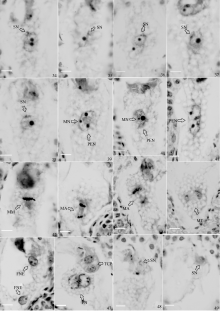
图版III
Plate III
| Figure OptionViewDownloadNew Window | |
| 图版III 34: 精核靠近极核; 35: 精核靠近次生核; 36: 精核贴在极核上; 37: 精核贴在次生核上; 38: 精核与次生核进一步融合; 39: 极核中出现小的雄性核仁; 40: 次生核中出现小的雄性核仁; 41: 初生胚乳核将要分裂前; 42: 初生胚乳核分裂中期: 43~44: 初生胚乳核分裂后期; 45: 初生胚乳核分裂末期; 46: 2个胚乳游离核; 47: 单受精, 中央极核没有受精, 卵细胞受精并且已经分裂; 48~49: 多精子进入同一个胚囊(47~49: 一个胚囊的不同切面)。(Bar=100 μm)Plate III 34: male nucleus close to polar nucleus; 35: male nucleus close to secondary endosperm nucleus; 36: male nucleus sticking to polar nucleus; 37: male nucleus sticking to secondary endosperm nucleus; 38: male nucleus and secondary endosperm nucleus further fused; 39: small male nucleus appearing in polar nucleus; 40: small male nucleus appearing in secondary endosperm nucleus (PEN); 41: primary endosperm nucleus before mitosis; 42: primary endosperm nucleus in mitosis metaphase; 43-44: primary endosperm nucleus in mitosis anaphase; 45: primary endosperm nucleus in mitosis telophase; 46: two free nuclei in the endosperm (FNE); 47: single fertilization phenomenon; 48-49: more sperms coming into embryonary sat (47-49: Different aspects of an embryo sac). (Bar=100 μm) |
致谢: 张仁庆、郑国赢、赵月婵同学帮助取材, 特此致谢。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
参考文献View Option
原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | |
| [2] | |
| [3] | |
| [4] | |
| [5] | |
| [6] | |
| [7] | |
| [8] | |
| [9] | |
| [10] | |
| [11] | |
| [12] | |
| [13] | |
| [14] |

{kind=link}
{kind=link}
{kind=link}