 , 吴存祥
, 吴存祥*通讯作者(Corresponding author): 韩天富, E-mail:hantianfu@caas.cn, Tel: 010-82105875 第一作者联系方式: E-mail:yanjiang416@163.com
** 对本文有相同贡献.
收稿日期:2014-03-04 基金资助:
摘要
关键词:大豆; 结荚习性; 顶端花序; 解剖学
Morphological and Anatomic Characteristics on Terminal Raceme Development of Soybean Varieties with Different Stem Termination Types
JIANG Yan
, WU Cun-XiangFund:
Abstract
Keyword:Soybean; Stem termination types; Terminal raceme; Anatomic
Show Figures
Show Figures


0 引言株型是指植株在空间上的分布态势, 是决定作物产量潜力的重要因素。良好的株型有利于作物充分利用光能, 适应肥、水条件及群体配置方式的变化, 进而获得高产[ 1, 2]。在大豆中, 结荚习性直接影响株高、节数、叶片大小等性状, 是最重要的株型性状之一, 其形成与地理、气候及土壤营养等生态条件均有关系[ 3, 4]。研究结荚习性的成因, 对改良和调节大豆株型, 提高大豆生态适应性均具有重要意义。
前人提出了多种大豆结荚习性的分类方法。有人将大豆的结荚习性划分为2种, 即有限结荚和无限结荚[ 5]; 有的将其分为3种, 即有限结荚、亚有限结荚和无限结荚[ 6, 7, 8]; 也有人将其分为5种, 即有限结荚、有限-亚有限结荚、亚有限结荚、亚有限-无限结荚和无限结荚[ 9]。另外, 有****通过大豆叶面积、叶片分布、茎粗、茎顶、花序等形态特点比较不同结荚习性类型的差异, 提出不同结荚习性的量化鉴别指标[ 10, 11, 12, 13, 14, 15], 但由于方法和指标较为复杂, 划分结果因人而异。目前普遍认同将大豆的结荚习性分为有限结荚、亚有限结荚和无限结荚等3类。
蒋青和李扬汉[ 16]通过解剖学方法观察不同结荚习性大豆品种的花芽分化过程认为, 无限型和有限型品种的顶端分生组织均可从营养生长转向生殖发育, 但无限型品种从植株基部开始花芽分化至主茎顶端开始花原基分化所经历的时间较有限型品种长。由此可推测, 不同大豆品种结荚习性的差异可能与顶端分生组织从营养生长向生殖发育转变的起始日期和历时长短有关, 但目前尚缺乏相应的形态学和解剖学方面的证据。此外, 在典型的有限结荚习性品种中, 顶端花序的长度也有明显的差异[ 17], 但其原因尚不清楚。
结荚习性是受少数基因控制的质量性状[ 10, 18, 19, 20], 并受外界环境因素的影响, 其中光周期是对大豆顶端花序影响较大的因素[ 4, 21, 22, 23, 24, 25]。对于光周期反应敏感的大豆品种, 长日照会导致顶端花序的发育时间延迟, 顶端花序长度增加[ 4], 甚至会出现花序部分逆转[ 4, 26, 27, 28, 29, 30]。本文利用解剖学手段, 研究不同光周期条件下不同结荚习性大豆品种顶端花序形态及分化过程的变化, 以期揭示大豆结荚习性和顶端花序长度差异形成的原因。
1 材料与方法1.1 试验材料选用中国农业科学院作物科学研究所育成的无限结荚习性品种中黄24、短花序有限结荚习性品种中黄13和辽宁省丹东市农业科学院育成的长花序有限结荚习性品种凤交66-12。这3个品种在北京地区的生育期基本相同。
1.2 光周期处理1.2.1 人工光周期处理 试验于2007年在中国农业科学院北京国家大豆改良分中心网室进行。短日照处理时数为12 h, 即7:00—19:00自然光照, 其余时间置暗室; 长日照处理光照长度16 h, 即6:00—22:00期间置室外, 日出前和日落后用白炽灯补足光照, 在植株顶端产生的光量子通量为28~50 µE s-1m-2。
于5月10日播种, 所用塑料桶高29 cm、上口直径32.5 cm。每个品种播种20桶(短日、长日处理各10桶)。播种前每桶装耕层土壤7.25 kg, 混施5 g (NH4)2HPO4。在同一光照条件下, 每个品种留6桶、每桶留5株进行生育时期的调查和考种, 其余材料用于解剖取样。人工光周期处理从植株子叶展开后(出苗后3 d)开始。
1.2.2 自然光照处理 于2007年5月8日在中国农业科学院作物科学研究所北圃场播种。试验采用随机区组设计, 行长3.5 m, 行距0.5 m, 3次重复, 出苗后人工间苗, 选留整齐一致豆苗, 其他栽培管理措施同大田。
1.3 测定项目与方法参考Fehr和Carviness[ 31]的大豆发育时期分期标准记载大豆植株生育时期, 包括营养生长期(V), 生殖生长期(R)。出苗期(Ve), 子叶期(Vc), 一节期(V1), 即单叶完全展开期, 二节期(V2), 即第一个三出复叶完全展开期, 余类推; 始花期(R1), 盛花期(R2), 始荚期(R3), 盛荚期(R4), 始粒期(R5), 满粒期(R6), 生理成熟期(R7), 完熟期(R8)。定期进行外部形态观察并照相, 成熟后收获、考种。
解剖学研究采用石蜡切片技术[ 32], 从Vc期至R1期隔日在固定时段取样, 每次取每品种顶芽20个。选取处在同一生长期、长势一致的大豆苗的顶芽, 用解剖针剥离顶芽, 只保留2~3个苞片, 用解剖刀切取大豆顶芽, 放入FAA固定液中固定24 h; 用50%、70%、85%的酒精溶液逐级脱水各30 min, 再经含0.5%伊红的95%酒精处理3 h, 无水酒精处理2次, 每次1 h; 依次用25%、50%、75%二甲苯的酒精溶液透明各30 min, 100%二甲苯处理3次, 每次30 min; 依次用50%二甲苯+50%石蜡、纯蜡在60℃烘箱内透蜡, 包埋; 用Leica RM 2235切片机切片, 然后在载玻片上粘片、在展台上展片、烘片; 用二甲苯、酒精、蒸馏水逐级脱蜡、复水; 用4%铁矾媒染30 min; 水洗后用0.5%苏木精染色; 在水洗后用2%铁矾分色至适度; 水洗返蓝后, 用酒精溶液逐级脱水, 在含1%伊红的95%酒精中复染, 透明、封片。最后用Olympus BX 51型光学显微镜对切片观察和拍摄, 采用Photoshop软件整理图片。
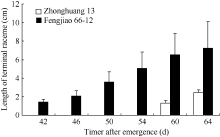
2 结果与分析2.1 不同结荚习性大豆品种顶端花序的发育过程在田间自然光照条件下, 供试的3个品种均于5月17日出苗。有限型短花序品种中黄13、长花序品种凤交66-12和无限型品种中黄24分别于6月24日(出苗后38 d)、6月26日(出苗后40 d)和7月2日(出苗后46 d)在主茎8~9节上出现第一朵开放的花朵。对茎顶形态的观察发现, 长花序品种凤交66-12在6月28日顶端花序即开始伸长, 中黄13在7月16日顶端花序开始伸长, 而中黄24到7月20日时仍未见顶端花序出现(图1)。不同结荚习性大豆品种的顶端花序形态和顶荚类型差别明显(图2)。
图1
Fig. 1
| Figure OptionViewDownloadNew Window | |
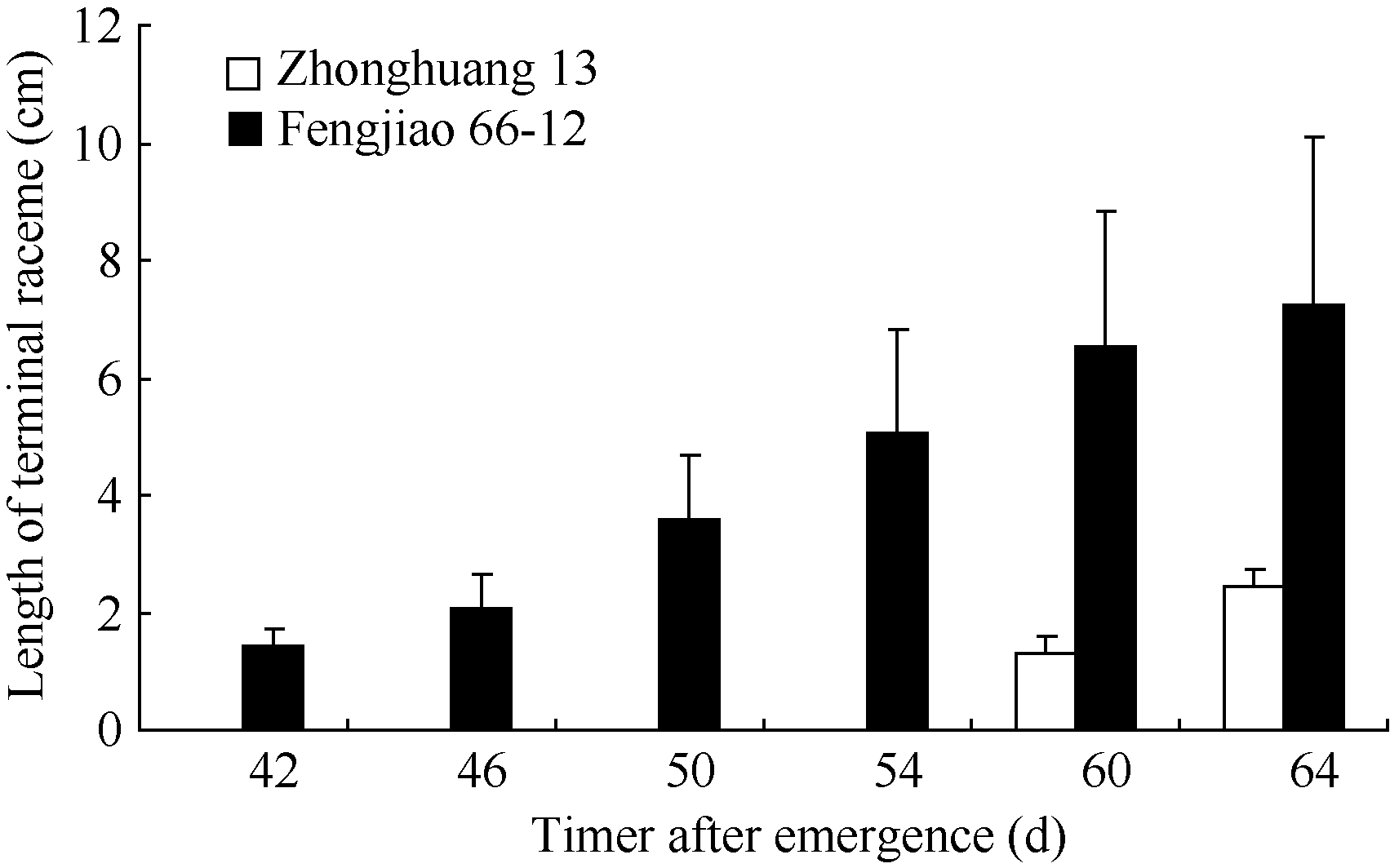 | 图1 自然光照条件下不同结荚习性大豆品种顶端花序 长度的变化动态Fig. 1 Changes of the terminal raceme length of soybean varieties with different stem termination types under the natural photoperiod |
图2
Fig. 2
| Figure OptionViewDownloadNew Window | |
 | 图2 自然光照条件下不同结荚习性大豆品种的顶端 花序的形态A、B和C分别为开花期中黄24、中黄13、凤交66-12的顶端花序形态(苗后60 d); D、E和F分别为结荚期中黄24、中黄13、凤交66-12的顶端豆荚形态(出苗后95 d)。Fig. 2 Terminal raceme morphological characteristics of soybean varieties with different stem termination types under the natural photoperiodA, B, and C are the terminal raceme of Zhonghuang 24, Zhonghuang 13, and Fengjiao 66-12 in flowering stage (taken at 60 days after emergence); D, E, and F are the terminal raceme of Zhonghuang 24, Zhonghuang 13, and Fengjiao 66-12 in pod bearing stage (taken at 95 days after emergence). |
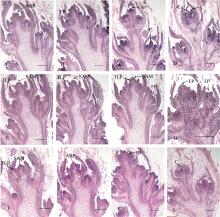
2.2 不同结荚习性大豆品种顶端花序的解剖学特征在田间自然光照条件下, 大豆品种中黄13、中黄24、凤交66-12的顶端花序发育过程分别见图3-A~D、E~H和I~L。出苗后24 d, 有限结荚习性品种中黄13 (V7期)植株茎顶已出现花序原基(图3-A), 而无限结荚习性品种中黄24的顶端分生组织仍处在叶原基的分化状态中(图3-E); 在出苗后30 d (V9期), 中黄13顶端分生组织已形成明显的花序结构, 在花序上着生许多小花原基(图3-B), 而中黄24顶端分生组织在叶腋处或产生分枝(图3-F), 或产生小花原基(图3-G), 茎顶端继续分化叶原基, 仍无花序原基出现(图3-F); 在出苗后36 d, 中黄13处于V11期, 顶端花序上着生7个小花, 最后一个小花已分化出萼片原基(图3-C), 而中黄24处于V12期, 其顶端分生组织仍然处于营养生长状态, 继续分化叶原基(图3-G); 在出苗后 40 d, 中黄13处于V12期, 其末端小花心皮原基已分化完全(图3-D), 而此时植株处于盛花期(V13期)的中黄24的顶端分生组织也形成单花(图3-H), 终止了器官分化。
在自然光照条件下, 中黄13顶端花序上第一个小花的苞片原基在出苗后24 d (V7期)产生, 最后一个小花的心皮原基在出苗后40 d (V12期)形成, 历时16 d (表1), 中黄24历时4 d, 凤交66-12历时12 d。有限结荚习性长花序品种凤交66-12茎顶端开始分化花序原基的时间较早, 在出苗后12 d (图3-I), 其茎顶芽处于营养生长阶段; 在出苗后20 d (图3-J), 茎顶产生花序组织, 比中黄13提前4 d; 在出苗后24 d (图3-K), 花序伸长, 其上的小花原基进一步分化; 在出苗后32 d (图3-L), 最后一朵小花的雌蕊原基出现, 历时12 d完成了整个花序的分化, 凤交66-12花序分化的时间与中黄13的相比少4 d (表1)。解剖学观察结果说明凤交66-12形成长花序的原因在于顶端花序分化的起始时间早, 且分化速度快。
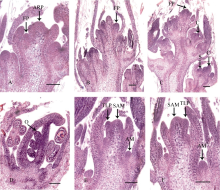
2.3 光周期对不同结荚习性大豆品种顶端花序分化的影响在短日照和长日照条件下, 中黄13顶端花序的分化过程差异明显。在出苗后12 d, 短日照处理的顶端分生组织开始分化出花序原基(图4-A), 并有小花原基的生成, 同期长日照处理的茎顶分生组织处在营养生长时期(图4-E), 不断分化出叶原基。中黄13顶端花序继续分化出小花原基(图4-B), 在出苗后20 d, 短日照处理的顶端花序上的小花已基本分化完毕(图4-C), 而长日照处理的茎顶分生组织仍处在营养生长状态(图4-F)。在出苗后28 d, 短日照处理下小花中的胚珠已形成(图4-D), 花粉母细胞处在四分体时期, 而长日照处理下茎顶分生组织还未见花原基的出现。中黄13在短日照条件下顶端花序发育历时为8 d (表2), 较自然光照条件下顶端花序的发育时间少8 d, 顶端花序形态发生了改变, 花序轴变短, 顶端荚数减少(图5-B), 部分植株从有花序轴的顶端花序变为无花序轴的多花, 类似短日照下的中黄24 (图5-A)。说明对同一品种来说, 短日照条件可使茎顶花序发育的起始日期提前, 且加快发育速度, 缩短发育进程, 导致植株提早成熟, 花序长度变短, 部分花序上小花出现败育, 顶端花序荚数减少。在可正常成熟的情况下, 日照的增加有利于大豆茎顶分生组织营养生长期的延长, 产生较多叶片, 利于光合产物的积累, 同时利于后期生殖发育时期养分供应, 为单株粒数和单株粒重的提高奠定基础[ 4]。
中黄24和凤交66-12在不同光照处理下茎顶发育状况与中黄13类似, 主要差别在于顶端花序分化持续时间不同。在短日照条件下, 凤交66-12的顶端花序分化持续日数比自然光照条件下少6 d (表2), 顶端花序明显缩短(图5-C), 部分植株与中黄13 (图5-B)类似, 顶端花序轴变短, 顶荚数减少。中黄24的顶端花序分化历时8 d (表2), 结荚习性从自然光照条件下的无限型转变为亚有限型, 茎顶从自然光照下的单花转变为短花序轴的多花(图5-A)。
图3
Fig. 3
| Figure OptionViewDownloadNew Window | |
 | 图3 自然光照条件下不同结荚习性大豆品种顶端花序发育过程的解剖学观察A: 出苗后24 d, 中黄13出现顶端花序原基; B: 出苗后30 d, 中黄13花序上出现小花原基; C: 出苗后36 d, 中黄13花序伸长, 小花原基进一步分化; D: 出苗后40 d, 中黄13花序上最后小花的雌蕊原基已分化完成; E: 出苗后24 d, 中黄24茎顶为营养生长阶段; F: 出苗后30 d, 中黄24茎顶仍为营养生长阶段; G: 出苗后36 d, 中黄24茎顶还处在营养生长阶段, 但花序下部已有花原基出现; H: 出苗后40 d, 中黄24茎顶出现小花原基; I: 出苗后12 d凤交66-12的茎顶处于营养生长阶段; J: 出苗后20 d, 凤交66-12已分化出顶端花序原基; K: 出苗后24 d, 凤交66-12的小花原基形成并分化; L: 出苗后32 d, 凤交66-12顶端花序上的小花已经基本分化完毕。ARP: 茎顶花序原基; C: 心皮; FP: 花原基; S: 雄蕊; SAM: 茎顶分生组织; TLP: 三出复叶原基。Bar: 200 μm。Fig. 3 Anatomic observations on the terminal raceme developmental process of soybean varieties with different stem termination types under the natural photoperiodA: the apical raceme primordium produced in the shoot tip of Zhonghuang 13 at 24 days after emergence; B: some flower primordia produced in the terminal raceme of Zhonghuang 13 at 30 days after emergence; C: the terminal raceme of Zhonghuang 13 elongated and the flower primordia further differentiated at 36 days after emergence; D: the carpel primordium of the terminal flower developed in the raceme of Zhonghuang 13 at 40 days after emergence; E: the shoot tip of Zhonghuang 24 was vegetative bud at 24 days after emergence; F: the shoot tip of Zhonghuang 24 was vegetative bud at 30 days after emergence; G: the shoot tip of Zhonghuang 24 was vegetative bud at 36 days after emergence, and flower primordium produced at the bottom of the inflorescence; H: the flower primordium produced in the terminal of Zhonghuang 24 at 40 days after emergence; I: the shoot tip of Fenjiao 66-12 was vegetative bud at 12 days after emergence; J: the apical raceme primordium produced in the shoot tip of Fenjiao 66-12 at 20 days after emergence; K: some flower primordia produced in the raceme of Fenjiao 66-12 at 24 days after emergence; L: some flowers developed in the raceme of Fenjiao 66-12 at 32 days after emergence. ARP: apical raceme primordium; C: carpel; FP: floral primordium; S: stamen; SAM: shoot apical meristem; TLP: trifoliate leaf primordium. Scale bar: 200 μm. |
表1
Table 1
表1(Table 1)
| 表1 在自然光照条件下不同结荚习性大豆品种顶端花序的分化持续时间 Table 1 Terminal raceme development duration of soybean varieties with different stem termination types under natural photoperiod |
顶端花序分化起始时间、顶端花序分化结束时间、顶端花序出现时间均为出苗后天数。
Beginning time of terminal raceme differentiation, ending time of terminal raceme differentiation, and the time to terminal raceme appearance on the plant were expressed by days after emergence.
图4
Fig. 4
| Figure OptionViewDownloadNew Window | |
 | 图4 光周期对中黄13茎顶发育的影响A: 出苗后12 d, 短日照处理的顶芽开始分化出花序原基, 并有小花原基的生成; B: 出苗后16 d, 短日照处理的花序上分化出许多小花原基, 呈现明显的花序结构; C: 出苗后20 d, 短日照处理的花序已分化完毕; D: 出苗后28 d短日处理的小花中的胚珠已形成, 花粉母细胞处在四分体时期; E和F: 长日照处理下, 在出苗后12 d和18 d, 茎顶芽仍处在营养分生组织阶段。AM: 叶腋分生组织; ARP: 茎顶花序原基; C: 心皮; FP: 花原基; O: 胚珠; SAM: 茎顶分生组织; TLP: 三出复叶原基。Bar: 100 μm.Fig. 4 Effects of photoperiod on the stem apex development of Zhonghuang 13A: the apical raceme primordium produced in the shoot tip at 12 days after emergence under short day treatment, and floral primordium produced in the axilla; B: many floral primordia produced in the terminal raceme at 16 days after emergence under short day treatment, to form terminal raceme structure; C: the terminal racemes developed at 20 days after emergence under short day treatment; D: the ovule developed at 28 days after emergence under short day treatment. the pollen mother cell was at the tetrad stage; E and F: the shoot tip was vegetative bud at 12 days and 18 days after emergence under long day treatment. AM: axillary meristem; ARP: apical raceme primordium; C: carpel; FP: floral primordium; O: ovule; SAM: shoot apical meristem; TLP: trifoliolate leaf primordium. Scale bar: 100 μm. |
表2
Table 2
表2(Table 2)
Beginning time of terminal raceme differentiation, ending time of terminal raceme differentiation, and the time to terminal raceme appearance on the plant were expressed by days after emergence. | 表2 短日照条件下不同结荚习性大豆品种顶端花序分化持续日数 Table 2 Terminal raceme development duration of soybean varieties with different stem termination types in short day condition | ||||||||||||||||||||||||
长日照下3个供试品种的茎顶发育进程均有不同程度的延迟, 在苗后60~75 d进入开花期。由于接近开花时植株高大, 切片较难, 其顶端花序分化的终止日期未通过解剖学观察加以确定。
3 讨论3.1 不同结荚习性大豆品种顶端花序发育过程的解剖学差异前人对大豆有限型和无限型结荚习性差异本质的研究结论不尽相同。Bernard[ 10]认为有限型大豆的茎生长到一定程度后会迅速终止生长, 而无限型大豆的茎则在相当长的时间内不断生长; 祝其昌[ 13]认为不同结荚习性大豆的本质差异在于其茎顶端花芽分化期的个体发育年龄不同; 蒋青和李扬汉[ 16]则认为不同结荚习性大豆的茎端都能转变为生殖茎端, 并分化出花原基, 主要差异表现在植株开始花芽分化后, 主茎端开始生殖发育的早晚不同。有限型大豆茎端的花原基能较好地发育, 并形成顶生花序; 而无限型大豆茎端的花原基大多数最后不能发育成荚, 植株一般只有少数几个甚至没有顶荚。本试验发现有限型品种茎顶形成花序原基的起始日期要比无限型品种早; 有限型品种的顶端花序分化时间长且发育充分, 并形成一定长度的顶端花序, 而无限型品种顶端分生组织花芽开始分化的时间晚, 茎顶最终未能形成明显的花序。这与祝其昌[ 13]、蒋青和李扬汉[ 16]的研究结果相符, 而与曹大铭[ 12]研究的无限结荚习性大豆茎端始终保持营养生长的观点不同。
图5
Fig. 5
| Figure OptionViewDownloadNew Window | |
 | 图5 短日照条件下不同结荚习性品种顶端花序的形态A、B和C分别为短日照下中黄24、中黄13、凤交66-12的顶端花序形态(出苗后37 d)。Fig. 5 Morphological characteristic of the terminal raceme in different stem termination types of soybean varieties in the short day treatmentA, B, and C were the terminal raceme of Zhonghuang 24, Zhonghuang 13 and Fengjiao 66-12 in the short day treatment at 37 days after emergence, respectively. |
本研究以大豆顶端花序上第一个小花的苞片原基产生到最后一个小花心皮原基出现之间的时间作为大豆顶端花序分化的时间指标。同样具有限结荚习性的凤交66-12和中黄13顶端花序长度不同的原因在于, 长花序品种凤交66-12开始形成顶端花序的时间比中黄13早, 且分化速度快, 花序上产生的小花原基多, 致使顶端花序延长。在小麦等作物中, 顶端花序(穗)的长度(小穗数)也由分化时间和分化速度共同决定[ 33]。
3.2 光周期对大豆结荚习性的影响大豆结荚习性除受自身遗传特性决定外, 还受环境因素的控制[ 4, 21, 22, 23, 24, 25]。为观察光周期对大豆不同结荚习性品种茎顶的影响, 本研究采用12 h和16 h光照进行全生育期处理, 发现中黄13、中黄24和凤交66-12从苗期到花期的时间和从苗期到始熟期的时间均随光照时间的延长而延长, 说明3个不同结荚习性大豆品种对光周期反应较为敏感。从开花期和成熟期的短日促进率来看, 中黄13对光周期的反应相对其他2个品种较钝感[ 34]。在短日照条件下, 有限结荚习性大豆品种的顶端花序分化时间较自然光照条件下减少, 花序长度缩短, 顶荚数减少, 结荚习性发生变化, 向亚有限或无限型转变, 而无限结荚习性大豆的顶端花序则向中间型-亚有限型转变。在长日照条件下, 不同结荚习性大豆品种的茎顶发育进程不同程度地延迟, 营养生长期延长, 开花期延后, 植株繁茂, 有利于后期养分供应及单株粒数和单株粒重的增加。
在小麦等作物中, 光照、温度及其光温互作[ 35]均对顶端花序的发育有重要影响。了解作物花序发育与光温环境的关系, 对因地制宜地选择品种和栽培管理均有指导意义。在大豆中, 不同结荚习性品种有一定的生态地理分布规律, 总趋势是北方大豆以无限结荚习性居多, 自北向南有限型品种逐渐增加, 至长江以南地区, 有限型品种占绝对优势[ 36]。我国南方及北方一些高温多湿、土壤肥沃地区适于栽培有限结荚习性大豆, 这种短日照和高气温的条件可促进大豆的生殖发育, 主要表现在顶端花序发育时间较早、分化进程快, 并能形成一定长度的顶端花序; 生长在我国东北中北部、西北地区、内蒙古以及晋北、陕北等地的大豆处于低温少雨、肥力较差条件, 适宜品种的顶端花序短, 顶荚数少[ 4], 故这些地区适于栽培无限结荚习性大豆, 其营养生长与生殖发育交错的时期长, 适应性较强; 亚有限结荚习性大豆适应于半湿润地区[ 37]。当然, 随着栽培条件的变化, 同一地区适宜大豆品种的结荚习性类型也会发生变化。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
参考文献View Option
原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | |
| [2] | |
| [3] | |
| [4] | |
| [5] | |
| [6] | |
| [7] | |
| [8] | |
| [9] | |
| [10] | |
| [11] | |
| [12] | |
| [13] | |
| [14] | |
| [15] | |
| [16] | |
| [17] | |
| [18] | |
| [19] | |
| [20] | |
| [21] | |
| [22] | |
| [23] | |
| [24] | |
| [25] | |
| [26] | |
| [27] | |
| [28] | |
| [29] | |
| [30] | |
| [31] | |
| [32] | |
| [33] | |
| [34] | |
| [35] | |
| [36] | |
| [37] |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}