
 , 何瑞亮1, 刘超1, 林保国2, 许大强2
, 何瑞亮1, 刘超1, 林保国2, 许大强21. 合肥工业大学资源与环境工程学院, 合肥 230009;
2. 安徽皖欣环境科技有限公司, 合肥 230601
收稿日期: 2021-01-25; 修回日期: 2021-03-18; 录用日期: 2021-03-18
基金项目: 国家自然科学基金项目(No.51579061);安徽省重点研究与开发计划项目(No.202004i07020005)
作者简介: 李如忠(1970—), 男, 教授(博士), E-mail: lrz1970@163.com
通讯作者(责任作者): 李如忠(1970—), 男, 教授, 博士, 主要从事水环境保护研究.目前已发表论文150余篇
摘要:为解析水库泄洪闸下溪流深潭氮磷营养盐滞留特征,选择NaBr为保守示踪剂,分别以NH4Cl和KH2PO4为添加营养盐,在合肥板桥河源头溪流开展野外示踪实验,据此估算氨氮(NH4+)、磷酸盐(PO43-)营养螺旋指标,识别主流区和暂态存储区NH4+、PO43-滞留贡献水平,模拟深潭地貌的氮磷吸收动力学特征.结果表明,深潭具有较好的氮磷滞留潜力,且对PO43-的滞留潜力超过NH4+;暂态存储区对NH4+的滞留贡献率平均为91.49%,表明NH4+滞留主要发生在暂态存储区;主流区对PO43-的滞留贡献率平均为96.09%,意味着主流区是PO43-滞留的主要场所;Michaelis-Menten(M-M)方程可以较好的模拟深潭氮磷滞留动力学效应,模拟得到的最大吸收速率Umax-NH4、Umax-PO4均值分别为0.48、0.08 mg·m-2·s-1,半饱和常数Km-NH4、Km-PO4均值分别为0.26、0.19 mg·L-1.
关键词:深潭氮磷滞留营养螺旋暂态存储动力学模拟
Retention characteristics and kinetics simulation of nitrogen and phosphorus in a stream pool under the sluice gate of a small reservoir
LI Ruzhong1
, HE Ruiliang1, LIU Chao1, LIN Baoguo2, XU Daqiang21. School of Resources and Environmental Engineering, Hefei University of Technology, Hefei 230009;
2. Anhui Wanxin Environmental Science and Technology Co. Ltd, Hefei 230601
Received 25 January 2021; received in revised from 18 March 2021; accepted 18 March 2021
Abstract: To investigate the retention characteristics of nitrogen and phosphorus in natural deep pool geomorphic pattern of streams,several field tracer experiments by taking NH4Cl and KH2PO4 as nutrient additions,and NaBr as a conservative tracer were performed in the headwater stream of Banqiao River,Hefei district. The nitrogen and phosphorus spiraling metrics were measured,and the contribution of main channel and transient storage zone to NH4+ and PO43- retention were estimated,respectively. Moreover,dynamics simulation for nitrogen and phosphorus retention was conducted as well. Results showed that the deep pool had a good potential for nitrogen and phosphorus retention,and retention potential for PO43- was higher than that of NH4+. NH4+ retention in transient storage zone was taken up to 91.49%,indicating that NH4+ retention mainly occurred in the transient storage zone. PO43- retention in main channel zone was up to 96.09%. In addition,Michaelis-Menten (M-M) equation could well simulate the dynamics of NH4+ and PO43- retention in the deep pool. The mean maximum uptake rates of Umax-NH4 and Umax-PO4 obtained from the M-M equation were 0.48 and 0.08 mg·m-2·s-1,respectively. And the half-saturation constants Km-NH4 and Km-PO4 were 0.26 and 0.19 mg·L-1,respectively.
Keywords: deep poolnitrogen and phosphate retentionnutrient spiralingtransient storagekinetic simulation
1 引言(Introduction)源头溪流不仅是陆生生态系统和水生生态系统物质循环的主要连接通道, 也是环境生态系统服务功能的重要提供者, 在汇流区域氮、磷等生源物质的滞留、传输和转化方面扮演着极为重要的角色(毛战坡等, 2015).究其原因, 一方面源头溪流因其数量庞大, 在河网水系总长度中所占比例较大, 蓄积的氮、磷营养盐量较为可观, 使得对下游水体水质影响不容轻视(Dollinger et al., 2015;李如忠等, 2018);另一方面由于源头溪流空间尺度较小, 自然和人为修复相对较为容易, 可有效提高生物地球化学反应速率(Claessens et al., 2010), 增加生物多样性(Swan et al., 2017), 并在非点源氮磷污染的过程阻断中发挥重要作用.目前, 有关源头溪流养分滞留的研究已成为环境科学、环境水文地球化学等学科领域的研究热点(李如忠等, 2017;2019;Gero et al., 2020;Martí et al., 2020).
深潭是溪流生态系统的一种特殊河床地貌形态, 是河道通过水流长期作用而逐渐形成的“急流-深潭-浅滩”结构单元的一部分, 常出现在河床比降相对较大的河段, 特别是溪流源头段.从成因看, 自然存在的深潭主要包括跌水冲淘、阶梯-深潭、河床坍塌、水溶侵蚀、侧向侵蚀等几种形式(王兵等, 2014).有研究发现, 阶梯、浅滩、深潭等河床地貌控制着溪流的暂态存储特征(Zarnetske et al., 2007; Baker et al., 2012; Weigelhofer et al., 2012), 阶梯-深潭结构的潜流交换强度甚至可以达到平坦河床的100倍(王伟, 2020).由于水动力条件弱, 水流流速低和水力停留时间长, 使得深潭内吸附在颗粒表面的营养盐易于沉降.李如忠等(2014b) 针对合肥城郊一条受生活污水影响的溪流沟渠氮磷滞留特征的研究表明, 溪流的坑状小水潭氨氮(NH4+)滞留速率较磷酸盐(PO43-)高出2~3个数量级, 而曲折渠段仅高出1~2个数量级, 表明在氮磷滞留效应的影响方面, 深潭和曲折渠段存在差异性.李如忠等(2017) 在对农田沟渠底部取土形成的坑槽状深潭地貌格局营养盐滞留的研究中, 发现深潭的氮磷滞留能力高于平直渠段, 且人为干扰措施可以进一步提高深潭的营养盐滞留潜力.一些研究也证明, 重塑溪流深潭-浅滩地貌格局, 对于提升河流水环境修复效果和环境生态功能大有裨益(Schwartz et al., 2015; Geoff et al., 2016), 因此在河流水环境修复中, 包括深潭在内的河床地貌结构重塑往往被推荐为经典的技术案例.
目前, 有关深潭或深潭-浅滩潜流交换作用的研究较多(Huang et al., 2018), 而对深潭格局营养盐滞留特征及其动力学模拟的研究则相对较少(Trauth et al., 2014), 对于深潭中主流区与暂态存储区营养盐滞留的相对贡献水平更是缺乏有效的分析与评估, 致使深潭营养盐滞留机制仍不是十分清楚.本研究拟以合肥市境内张桥水库泄洪闸下板桥河左侧源头支流为对象, 针对水库泄洪冲刷形成的代表性河床深潭地貌格局, 借助野外示踪实验和数学模拟, 分析深潭中主流区和暂态存储区的氮磷滞留的相对贡献水平, 并解析深潭氮磷滞留的动力学特征, 以期为科学评估溪流氮磷滞留能力提供依据, 也为小河流环境生态功能修复提供参考.
2 研究区概况(Site description)板桥河位于合肥城区北部, 为南淝河上游左岸重要支流.河流源头地处城市郊外, 左右两条一级支流分别位于小型水库张桥水库、蔡塘水库泄洪闸下.由于地处江淮分水岭南侧, 流域中上游低矮丘陵岗地地貌特征占主导.张桥、蔡塘水库周边及下游汇水区基本保持农村自然风貌, 农田、村落和林地用地占据主导地位.由于水库泄洪冲刷, 在闸下溪流段形成了多处明显的深潭-浅滩地貌格局, 而且溪流岸坡崩塌明显, 致使水面宽度、水深的分布极不均匀.
本研究的深潭位于张桥水库下方, 溪流长约4.5 km.选取的代表性深潭位于溪流中部, 两侧为岗坡, 左侧岗坡分布稀疏林地, 右侧岗坡杂草丛生.溪流深潭的实验段长约100 m, 包括了大小两个深潭, 小深潭位于大深潭的上方, 两者相距约20 m.其中, 小深潭长约10.0 m, 平均宽度2.0 m, 平均水深0.70 m;大深潭长约35 m, 平均宽度8.6 m, 平均水深0.65 m, 最大水深1.5 m.大深潭的两侧高坡坍塌显著, 形成了几近垂直的壁岸, 高度约3~7 m.由于泥沙被泄洪冲走, 在河床上遗留下大量的碎石和卵石.该深潭底部主要为砾石和粗砂, 淤泥和细沙很少, 整个河床无大型水生/湿生植物, 但在潮湿和浅水的碎石表面却附生着大量的苔藓状或丝状生物膜.溪流水体水质状况良好, NH4+浓度约为0.41~0.52 mg·L-1, SRP浓度约为0.030~0.051 mg·L-1.
3 材料与方法(Materials and methods)3.1 示踪实验设计选择NaBr为保守示踪剂, 以NH4Cl和KH2PO4为添加营养盐, 分别于2019年11月6、13日和12月6、20日, 采用瞬时投加的方式开展4次野外示踪实验.其中, 11月6日和12月6日为NH4Cl、NaBr混合投加实验, 其余两次为KH2PO4、NaBr混合投加实验.在示踪实验的前一天现场勘探水情, 分析测定Br-、NH4+、PO43-环境背景浓度.在此基础上, 根据预期达到的峰值浓度水平, 合理确定示踪剂和营养盐的投加量.
投加点O设置在小深潭前方25 m, 采样点A位于大深潭前端15 m, 靠近小深潭下端.考虑到大深潭后部的水面宽、水深浅特点, 为便于采样, 在大深潭后部的溪流渠段布设采样点B.其中, OA长度为40 m, AB长度为60 m.根据经验, 小深潭水力条件可以确保投加的示踪剂和营养盐在抵达采样点A时满足断面浓度均匀的要求.现场利用溪水将示踪剂与营养盐充分混合.为获得完整的示踪剂和营养盐浓度穿透曲线(break through curves, BTCs), 现场采用便携式电导率仪测试水样电导率, 待到电导率值稳定回到环境背景水平即停止采样.利用100 mL塑料瓶采集水样, 采样点A的采样间隔时间为20 s, 根据流速情况将采样点B的间隔时间设置为40 s和1 min两种情形.示踪实验结束后, 对OA、AB段内的多个断面, 分别测定水深、水面宽度和流速等基本信息.返回实验室后, 利用PXS-215离子活度计和溴离子选择性电极测定Br-浓度, NH4+、PO43-浓度分别采用纳氏试剂分光光度法和钼酸铵分光光度法测定.
3.2 模型与方法3.2.1 营养螺旋指标营养螺旋指标中营养盐吸收长度、吸收速度和吸收速率是小河流营养盐滞留潜力评估中较为常用的3项指标, 相应的数学表达式分别为:
 | (1) |
 | (2) |
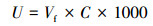 | (3) |
3.2.2 营养盐吸收贡献水平主流区和暂态存储区构成了溪流营养盐滞留的场所.其中, 主流区通常泛指溪流过水断面中流动性相对较为显著的部分, 而暂态存储区则主要包括水底潜流带、漩涡、滞水区(或死水区)等.根据示踪实验获得的BTCs, 利用选定的水质模型, 如OTIS模型(Runkel, 1998), 开展实验过程中溪流4种情景的水质模拟, 即不考虑主流区和暂态存储区生物滞留效应的情景1、考虑主流区和暂态存储区生物滞留效应的情景2、只考虑主流区营养盐生物滞留效应的情景3及只考虑暂态存储区营养盐滞留生物作用的情景4.这里, 不妨以Si(i=1, 2, 3, 4)分别表征上述4种情景的溪流深潭营养盐通量, 该值可以根据营养盐浓度模拟曲线围成的平面面积与溪流流量的乘积进行估算(McKnight et al., 2004), 即S=∫QCdt=Q∫Cdt.
基于上述4种情景的模拟结果, 估算评估主河道流动水体、暂态存储区对营养盐滞留的相对贡献水平(O′Connor et al., 2010; Argerich et al., 2011):
 | (4) |
 | (5) |
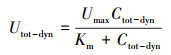 | (6) |
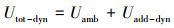 | (7) |
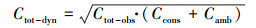 | (8) |
限于篇幅, 有关变量Uamb、Uadd-dyn和Ccons的具体计算方法, 参见文献(Covino et al., 2010; 李如忠等, 2018).
4 结果与分析(Results and discussion)4.1 深潭NH4+和PO43-浓度穿透曲线图 1为4次示踪实验中溪流深潭采样点A、B相应的NH4+和PO43-浓度穿透曲线.可以看出, 采样点A的NH4+和PO43-浓度曲线陡峭, 峰值两侧浓度梯度变化迅速, 而采样点B的浓度曲线则变化较为平缓, 且右侧拖尾现象较为显著.采样点A、B浓度穿透曲线峰值的巨大落差, 直观地体现了深潭地貌对于投加营养盐的显著调控效应, 表明自然深潭对氮、磷营养盐滞留的有效性.当然, NH4+和PO43-在深潭前后发生的浓度曲线的显著变化性离不开物理、化学和生物因素的共同作用.
2019年11月6日和12月6日示踪实验相应的自然深潭NH4+环境背景值分别为0.50、0.48 mg·L-1, 水温分别为15.4、10.2 ℃;2019年11月13日和12月20日的PO43-环境背景值分别为0.051、0.037 mg·L-1, 水温分别为16.9、8.6 ℃.
图 1(Fig. 1)
 |
| 图 1 NH4+和PO43-浓度穿透曲线 Fig. 1Breakthrough curves of simulated and observed NH4+ and PO43- concentrations |
4.2 深潭氮磷滞留潜力评估目前, 针对小河流暂态存储和营养盐滞留潜力评估的研究, 大多数都是依托OTIS模型, 利用Runkel (1998)开发的模型求解应用程序和OTIS-P参数自动优化包, 计算OTIS模型参数, 然后再利用其它有关模型进行定量化评估.2019年11月6日和12月6日示踪实验中, 采样点B的NH4+浓度模拟值与实测值的相对误差分别为4.12%、3.73%;2019年11月13日和12月20日示踪实验中, 该采样点的PO43-浓度模拟值与实测值的相对误差分别为5.28%、4.47%.据此, 模拟得到的深潭OTIS模型参数A、As、α、D以及营养盐吸收系数λ和λs, 见表 1.这里, A和As分别表示深潭主流区和暂态存储区断面面积(m2);α表示主流区与暂态存储区之间的交换系数(s-1);D表示深潭水流扩散系数(m2· s-1);λ、λs分别表示主流区和暂态存储区的营养盐一阶吸收系数(s-1).
4次示踪实验的交换系数α都处于10-4数量级, 与流动性较好的溪流沟渠相比, 基本上低1个数量级(李如忠等, 2014a;2016).As/A值常被用于定性评估溪流暂态存储区溶质滞留潜力的高低(Runkel, 2007).本研究中, 两次NH4+投加实验模拟得到的深潭As/A比值分别为0.52、0.59, 两次PO43-投加实验的比值分别达0.41、0.46, 较一般溪流沟渠水体高数倍(Weigelhofer et al., 2012;李如忠等, 2014b;2017), 表明就物理条件而言, 由水库泄洪冲刷形成的深潭具备了相当高的溶质滞留潜力.当然, 该深潭对于NH4+、PO43-的实际滞留效果, 可能还需要综合考虑其他因素的影响, 如营养盐理化特性、生物因素等.这里, 主流区与暂态存储区NH4+、PO43-一阶衰减系数基本处于10-4~10-3数量级, 较合肥地区其他源头溪流沟渠高出1~2个数量级(李如忠等, 2016).尽管4次实验在季节上都处于秋末冬初时节, 但实地调查仍发现, 在深潭浅水区块石、卵石及枯枝表面依旧不同程度地附着苔藓状或丝状生物膜, 且生长状况良好.由此, 可以推断生物因素的影响对于NH4+、PO43-滞留的贡献可能不容忽视.
在11月6日的NH4+滞留实验中λs为负值, 意味着暂态存储区存在NH4+释放现象, 可能与深潭中相对较高的NH4+浓度有很大的关系(李如忠等, 2015).事实上, 由于自然深潭NH4+的环境背景值基本上达到了生物吸收饱和水平, 在其他因素的综合作用下完全可能发生NH4+释放(Haggard et al., 2005; Sánchez-Pérez et al., 2009).
表 1(Table 1)
| 表 1 深潭OTIS模型参数 Table 1 Hydrological parameters of OTIS model in the deep pool | |||||||||||||||||||||||||||||||||||||||||||
表 1 深潭OTIS模型参数 Table 1 Hydrological parameters of OTIS model in the deep pool
| |||||||||||||||||||||||||||||||||||||||||||
根据表 1的相关参数值, 采用式(9)计算自然深潭营养盐综合衰减系数k(Runkel, 2007), 即:
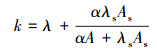 | (9) |
表 2(Table 2)
| 表 2 深潭氮磷营养盐螺旋指标 Table 2 Nutrient spiraling metrics and comprehensive degradation coefficients for NH4+ and PO43- | ||||||||||||||||||||||||||||
表 2 深潭氮磷营养盐螺旋指标 Table 2 Nutrient spiraling metrics and comprehensive degradation coefficients for NH4+ and PO43-
| ||||||||||||||||||||||||||||
4.3 深潭营养盐吸收贡献评估根据λ和λs的物理含义, 可以将式(4)和式(5)所对应的4种模拟情景分别表示如下, 即情景1: λ=0, λs=0;情景2: λ>0, λs>0;情景3: λ>0, λs=0;情景4: λ=0, λs>0.针对上述4种情景, 利用OTIS模型分别开展水质模拟, 据此计算各情景中采样点B的NH4+或PO43-通量, 进而得到深潭主流区和暂态存储区对NH4+和PO43-的滞留贡献水平(李如忠等, 2016), 结果见表 3.对于NH4+而言, 暂态存储区的平均滞留贡献率高达91.49%, 远高于主流区的8.51%, 表明该深潭的NH4+滞留主要发生在暂态存储区.而PO43-恰好相反, 主流区的滞留贡献率远高于暂态存储区, 平均值分别为96.09%和3.91%, 意味着深潭中PO43-滞留主要发生在主流区.也就是说, 水库泄洪闸下溪流深潭NH4+和PO43-滞留效应的主要发生场所不一致.
表 3(Table 3)
| 表 3 主流区和存储区对营养盐吸收贡献率 Table 3 Fraction of nutrients uptake between main channel and storage zones | ||||||||||||||||||
表 3 主流区和存储区对营养盐吸收贡献率 Table 3 Fraction of nutrients uptake between main channel and storage zones
| ||||||||||||||||||
McKnight等(2007) 在对冰川融雪溪流的研究中也发现, 似乎只有主流区对PO43-存在吸收滞留作用, 而潜流带的影响几乎可以忽略不计;李如忠等(2014b) 针对巢湖十五里河源头规则化、渠道化河段的研究, 得到主流区对PO43-损失的平均贡献率高达99.70%, 而暂态存储对于NH4+损失的贡献率则为93.82%, 该结果与本研究非常接近.当然, 主流区和暂态存储区对于NH4+、PO43-滞留的相对贡献率分配关系可能并不是绝对的、固定的, 如Argerich等(2011) 对不同底质人工沟渠的研究中, 主流区不仅可以解释92.0%左右的PO43-滞留, 也可以解释约96.1%的NH4+滞留, 即主流区是NH4+和PO43-滞留的主要发生场所.李如忠等(2016) 针对芦苇占优势农田溪流沟渠的研究, 得到主流区和暂态存储区对NH4+滞留的平均贡献率分别为43.12%和56.88%, 对PO43-的平均贡献率分别为50.13%和49.87%.
4.4 深潭营养盐吸收动力学模拟采用TASCC技术, 定量刻画4次实验的深潭NH4+或PO43-总动态吸收速率Utot-dyn与其总动态浓度Ctot-dyn的关系.图 2展示了经由M-M方程拟合得到的95%置信水平下2019-11-06和2019-12-20实验中Utot-dyn~Ctot-dyn关系曲线, 其他两次示踪实验也得到了类似的关系曲线图.需要指出的是, 在模拟计算过程中, 并不是BTCs曲线上所有样本数据均可以满足TASCC技术要求, 需要事先进行数据的筛选处理, 具体要求及操作办法参见文献(李如忠等, 2018).本研究中, 符合要求的样本点数据基本都落在95%置信水平区间或毗邻范围内.由图 2可见, 随着NH4+或PO43-总动态浓度的增加, 采样点A、B营养盐吸收速率的增幅变缓, 特别是2019-11-06实验表现得更为明显.
图 2(Fig. 2)
 |
| 图 2 在95%置信水平下NH4+和PO43-动态吸收过程模拟 Fig. 2M-M model fitting for NH4+ and PO43- retention dynamics under 95% confidence intervals |
拟合得到的4次实验M-M方程参数值, 见表 4.显然, 该深潭前、后两个采样点位的最大吸收速率Umax值悬殊较大, 尤其是PO43-, 意味着自然深潭对NH4+、PO43-滞留效果显著.半饱和常数Km是一个可以间接反映生物对营养盐亲和力水平的指标, Km越低意味着生物对其亲和力越高, 也就是生物对营养盐的响应速度越快(Piper et al., 2017).本研究中, 采样点B的Km-PO4略低于Km-NH4, 表明深潭对于PO43-的滞留效应超过NH4+, 这与根据营养盐螺旋指标得到的评价结果一致.总体上, 深潭下方采样点B的半饱和常数较深潭上方采样点A更为稳定.
表 4(Table 4)
| 表 4 深潭氮磷吸收动力学M-M模型拟合参数 Table 4 M-M model parameters for NH4+ and PO43- in the deep pool | ||||||||||||||||||||||||||||||||||||||||||||
表 4 深潭氮磷吸收动力学M-M模型拟合参数 Table 4 M-M model parameters for NH4+ and PO43- in the deep pool
| ||||||||||||||||||||||||||||||||||||||||||||
4.5 讨论一般认为, 湖泊、水库等水体氮滞留的主导机制是表层沉积物的反硝化脱氮反应, 而磷的主导机制则为颗粒态的沉降作用.同样, 溪流中氮磷滞留效率也可能因NH4+和PO43-两者滞留机理的不同而表现出较为明显的差异性.如Svendsendeng等(1993)、Russell等(1998) 针对农田溪流和排水沟渠中氮磷迁移转化过程的研究发现, 约70%以上的磷是通过颗粒态传输, 而氮则不足10%.尽管在外形上深潭与水塘较为类似, 但潭中相对明确的水体流动方向和较好的流动性特征, 可以为深潭源源不断地输送含氧量相对较高的上游来水, 使得深潭底部往往难以形成显著的厌氧或缺氧环境条件, 故此在水深不是很大的情况下, 反硝化脱氮不大可能成为溪流深潭氮滞留的主导机制.但若深潭水深较大, 在潭底部就可能产生缺氧水层, 这对反硝化作用的发生是有利的, 特别是当上游来水的水量和水动力条件不足时.就本研究而言, 受水库泄洪冲刷的影响, 河床掏蚀较为严重, 深潭最深处水深可达1.5 m.而且, 两侧土坡崩塌形成几近垂直的壁岸, 并在河床和深潭底部留下了大量碎石和粗砂, 但却很少有淤泥沉积物的存在.实地调查发现, 整个自然深潭都没有大型水生/湿生植物, 仅在深潭后部的河床浅滩存在一些大型水生植物和枯枝落叶残体, 也就是说深潭中几乎不具备发生反硝化作用的沉积物环境条件.一般地, NH4+易于被带负电荷的土壤颗粒所吸附, 而PO43-则可能通过与Al3+、Fe3+等发生化学反应而沉淀.而溪流深潭因自身特殊的地貌结构特征, 可以增加营养盐的水力停留时间, 从而更有利于氮磷颗粒物的滞留.
众所周知, 斜板沉淀池相对于常规的沉淀池可以更为有效地去除颗粒态污染物, 从而提升水处理设施的工作能力.笔者以为, 对于溪流深潭地貌格局的NH4+、PO43-滞留, 抛开生物因素的影响, 单纯从颗粒态氮磷营养盐沉淀或沉降去除的角度, 也可以通过对深潭采取一定的人工改造措施达到氮磷滞留能力提升的目的.李如忠等(2017) 针对人为取土形成的农田沟渠坑状深潭地貌格局, 按照一定的间距在深潭底部垂直水流方向平行放置一些装填泥土的编织袋, 从而在增大河床粗糙度的同时, 营造更多有利于暂态存储的涡流场, 结果发现NH4+吸收长度明显降低, 即Sw-NH4由331~3304 m下降为232~609 m.对于本研究水库泄洪冲刷形成的溪流深潭而言, 向潭中投放大块石或石笼或许也是NH4+滞留能力提升的可行方法.事实上, 投放石笼也可以为微生物提供更多的附着基质和界面, 对于发挥氮磷的生物滞留效应大有补益.但文献(李如忠等, 2017) 中的人工改造却造成PO43-滞留能力的下降, 即Sw-PO4由232~609 m上升为301~1100 m.因此, 对于磷富集相对较为突出的情形, 该技术或许并不合适.此外, 由于硝态氮带负电荷, 无法与同带负电荷的泥沙颗粒吸附而沉降, 在深潭地貌格局中很难通过这一途径实现滞留.不难看出, 如何通过调控深潭暂态存储潜力实现氮、磷滞留能力的同向提升, 仍需要深入探究.
毋庸讳言, 本研究中4次实验均安排在秋末冬初时节, 对于生物活性相对较高的夏、春季并未继续开展相关示踪实验, 因此无法对不同季节的研究结果进行比较.究其原因, 主要来自于两个方面, 一是受2020年上半年新冠肺炎大流行的影响, 自由出行受到限制, 导致春、夏季野外示踪实验无法正常进行;二是由于2020年合肥地区梅雨降雨量与历史同期相比偏高数倍, 张桥水库泄洪频次和强度显著高于以往年份, 导致本研究所在的溪流深潭损毁严重, 即便开展实验, 相关结果与本研究也很难具有可比性.实际上, 本研究已可以反映秋冬季节水库泄洪闸下溪流深潭NH4+和PO43-滞留潜力和动力学特征.可以断言, 从营养盐滞留的物理作用机制来看, 水库泄洪闸下溪流深潭与山区、丘陵岗地小河流深潭应该具有高度的相似性.至于化学作用和生物因素的影响, 则可能更多地取决于溪流营养盐环境背景浓度以及生物种群的差异.
5 结论(Conclusions)1) 水库泄洪闸下溪流深潭主流区与暂态存储区的交换作用较一般溪流水体弱, 但As/A比值却相对更高, 意味着深潭水体具有较大的暂态存储潜力.
2) 溪流深潭NH4+和PO43-滞留潜力较高, 且对PO43-的滞留潜力稍高于NH4+, 两种营养盐均可以在溪流长度内实现滞留.
3) 深潭对NH4+吸收主要发生在暂态存储区, 平均贡献率为91.49%;对PO43-的滞留贡献则主要发生在主流区, 平均贡献率为96.09%.
4) 采用M-M方程可以很好的模拟深潭的NH4+和PO43-吸收动力学特征, 且自然深潭段的最大吸收速率Umax-NH4、Umax-PO4平均值分别为0.48、0.08 mg· m-2· s-1, 半饱和常数Km-NH4、Km-PO4平均值分别为0.26、0.19 mg· L-1.
参考文献
| Argerich A, Martí E, Sabater F, et al. 2011. Influence of transient storage on stream nutrient uptake based on substrata manipulation[J]. Aquatic Sciences, 73(3): 365-376. DOI:10.1007/s00027-011-0184-9 |
| Baker D W, Bledsoe B P, Price J M. 2012. Stream nitrate uptake and transient storage over a gradient of geomorphic complexity, northcentral Colorado, USA[J]. Hydrological Processes, 26(21): 3241-3252. DOI:10.1002/hyp.8385 |
| Claessens L, Tague C L, Groffman P M, et al. 2010. Longitudinal and seasonal variation of stream N uptake in an urbanizing watershed: Effect of organic matter, stream size, transient storage and debris dams[J]. Biogeochemistry, 98(1/3): 45-62. |
| Covino T P, McGlynn B L, McNamara R A. 2010. Tracer additions for spiraling curve characterization (TASCC): Quantifying stream nutrient uptake kinetics from ambient to saturation[J]. Limnology and Oceanography: methods, 8(9): 484-498. DOI:10.4319/lom.2010.8.484 |
| Dollinger J, Dagès C, Bailly J S, et al. 2015. Managing ditches for agroecological engineering of landscape. A review[J]. Agronomy for Sustainable Development, 35(3): 999-1020. DOI:10.1007/s13593-015-0301-6 |
| Geoff J V, Rutherfurd I D, Fletcher T D, et al. 2016. Thinking outside the channel: Challenges and opportunities for protection and restoration of stream morphology in urbanizing catchments[J]. Land and Urban Planning, 145: 34-44. DOI:10.1016/j.landurbplan.2015.09.004 |
| Gero W, Ulrich H, Jochen K. 2020. Nitrate dynamics in springs and headwater streams with agricultural catchments in southwestern Germany[J]. Science of the Total Environment, 722: 137858. DOI:10.1016/j.scitotenv.2020.137858 |
| Haggard B E, Stanley E H, Storm D E. 2005. Nutrient retention in a point-source-enriched stream[J]. Journal of North American Benthological Society, 24(1): 29-47. DOI:10.1899/0887-3593(2005)024<0029:NRIAPS>2.0.CO;2 |
| Huang P, Chui T F M. 2018. Empirical equations to predict the characteristics of hyporheic exchange in a pool-riffle sequence[J]. Ground Water, 56(6): 947-958. DOI:10.1111/gwat.12641 |
| 李如忠, 曹竟成, 张瑞钢, 等. 2016. 芦苇占优势农田溪流营养盐滞留能力分析与评估[J]. 水利学报, 47(1): 28-37. |
| 李如忠, 丁贵珍. 2014a. 基于OTIS模型的巢湖十五里河源头段氮磷滞留特征[J]. 中国环境科学, 34(3): 742-751. |
| 李如忠, 董玉红, 钱靖, 等. 2015. 合肥地区不同类型源头溪流暂态存储能力及氮磷滞留特征[J]. 环境科学学报, 35(1): 128-136. |
| 李如忠, 耿若楠, 黄青飞, 等. 2017. 农田溪流深潭营养盐滞留及对人为干扰的响应[J]. 中国环境科学, 37(2): 720-729. |
| 李如忠, 杨继伟, 钱靖, 等. 2014b. 合肥城郊典型源头溪流不同渠道形态的氮磷滞留特征[J]. 环境科学, 35(9): 3365-3372. |
| 李如忠, 殷齐贺, 高苏蒂, 等. 2018. 农业排水沟渠硝态氮吸收动力学特征及相关性分析[J]. 环境科学, 39(5): 2174-2183. |
| 李如忠, 许大强, 阙凤翔. 2019. 源头溪流氮磷耦合吸收效应及动力学模拟[J]. 中国环境科学, 39(2): 648-656. DOI:10.3969/j.issn.1000-6923.2019.02.026 |
| 毛战坡, 杨素珍, 王亮, 等. 2015. 磷素在河流生态系统中滞留的研究进展[J]. 水利学报, 46(5): 515-524. |
| Martí E, Feijoo C, Vilches C, et al. 2020. Diel variation of nutrient retention is associated with metabolism for ammonium but not phosphorus in a lowland stream[J]. Freshwater Science, 39(2): 268-280. DOI:10.1086/708933 |
| McKnight D M, Runkel R L, Tate C M, et al. 2004. Inorganic N and P dynamics of antarctic glacial meltwater streams as controlled by hyporheic exchange and benthic autotrophic communities[J]. Journal of North American Benthological Society, 23(2): 171-188. DOI:10.1899/0887-3593(2004)023<0171:INAPDO>2.0.CO;2 |
| O'Connor B L, Hondzo M, Harvey J W. 2010. Predictive modeling of transient storage and nutrient uptake: implications for stream restoration[J]. Journal of Hydraulic Engineering, 136(12): 1018-1032. DOI:10.1061/(ASCE)HY.1943-7900.0000180 |
| Piper L R, Cross W F, Mcglynn B L. 2017. Colimitation and the coupling of N and P uptake kinetics in oligotrophic mountain streams[J]. Biogeochemistry, 132(1/2): 165-184. |
| Runkel R L. 1998. One-dimensional transport with inflow and storage (OTIS): A solute transport model for streams and rivers US Geological Survey Water-Resources Investigations Report, 98-4018[R]. Washington, D C: US Geological Survey |
| Runkel R L. 2007. Toward a transport-based analysis of nutrient spiraling and uptake in stream[J]. Limnology and Oceanography: Methods, 5: 50-62. DOI:10.4319/lom.2007.5.50 |
| Russell M A, Walling D E, Webb B W, et al. 1998. The composition of nutrient fluxes from contrasting UK river basins[J]. Hydrological Processes, 12(9): 1461-1482. DOI:10.1002/(SICI)1099-1085(199807)12:9<1461::AID-HYP650>3.0.CO;2-6 |
| Sánchez-Pérez J-M, Gérino M, Sauvage S, et al. 2009. Effects of wastewater treatment plant pollution on in-stream ecosystems functions in an agricultural watershed[J]. International Journal of Limnology, 45(2): 79-92. DOI:10.1051/limn/2009011 |
| Schwartz J S, Neff K J, Dworak F E, et al. 2015. Restoring riffle-pool structure in an incised, straightened urban stream channel using an ecohydraulic modeling approach[J]. Ecological Engineering, 78: 112-126. DOI:10.1016/j.ecoleng.2014.06.002 |
| Svendsen L M, Kronvang B. 1993. Retention of nitrogen and phosphorus in a Danish lowland river system: Implications for the export from the watershed[J]. Hydrobiologia, 251(1/3): 123-135. |
| Swan C M, Brown B L. 2017. Metacommunity theory meets restoration: isolation may mediate how ecological communities respond to stream restoration[J]. Ecological Applications, 27(7): 2209-2219. DOI:10.1002/eap.1602 |
| Trauth N, Schmidt C, Vieweg M, et al. 2014. Hyporheic transport and biogeochemical reactions in pool-riffle systems under varying ambient groundwater flow conditions[J]. Journal of Geophysical Research: Biogeosciences, 119: 910-928. DOI:10.1002/2013JG002586 |
| 王兵, 刘慧博, 李峪, 等. 2014. 深潭的功能及在城市河道治理中的构建模式[J]. 中国水土保持科学, 12(3): 107-112. |
| 王伟. 2020. 阶梯-深潭结构下的潜流交换过程试验研究[J]. 水电能源科学, 38(7): 34-36. |
| Weigelhofer G, Fuchsberger J, Teufl B, et al. 2012. Effects of riparian forest buffers on in-stream nutrient retention in agricultural catchments[J]. Journal of Environmental Quality, 41(2): 373-379. DOI:10.2134/jeq2010.0436 |
| Zarnetske J P, Gooseff M N, Brosten T R, et al. 2007. Transient storage as a function of geomorphology, discharge, and permafrost active layer conditions in Arctic tundra streams[J]. Water Resources Research, 43: W07410. DOI:10.1029/2005WR004816 |
