 ,1,2,3
,1,2,3A reappraisal of the Silurian galeaspids (stem-Gnathostomata) from Tarim Basin, Xinjiang
LIU Yu-Hai1, ZHU Min1,2,3, LIN Xiang-Hong1,4, LU Li-Wu5, GAI Zhi-Kun,1,2,3通讯作者: *gaizhikun@ivpp.ac.cn
收稿日期:2019-02-22网络出版日期:2019-10-20
| 基金资助: |
Corresponding authors: *gaizhikun@ivpp.ac.cn
Received:2019-02-22Online:2019-10-20

摘要
通过对新疆塔里木盆地志留系新一轮野外考察和对原来塔里木志留纪盔甲鱼类再观察,取得了以下认识:1) 依据新增加的材料, 张氏南疆鱼(Nanjiangaspis zhangi)被厘定为一新属西域鱼(Xiyuaspis), 并确定其系统位置隶属修水鱼科(Xiushuiaspidae); 2) 潘氏小瘤鱼(Microphymaspis pani)曾被归于Xiushuiaspidae, 系缘于观察上的失误,通过再观察得出M. pani当属大庸鱼科(Dayongaspidae), 同时与其产于同一产地和层位的天山宽头鱼(Platycaraspis tianshanensis)系前者的同物异名;3) 经重新研究,产于柯坪克兹尔塔格组曾被视为节甲鱼类的标本实属盔甲鱼类,并被鉴定为一新的属种:残缺角鱼(Jiaoyu imperfectus); 4) 产于柯坪塔塔埃尔塔格组的汉阳鱼未定种(Hanyangaspis sp.)的前腹片和产于柯坪塔塔埃尔塔格组及巴楚依木干他乌组的锅顶山汉阳鱼(H. guodingshanensis)具雪花状纹饰的骨片,可能均属柯坪南疆鱼(Nanjiangaspis kelpinensis); 5) 根据所产盔甲鱼残缺角鱼(J. imperfectus), 将克兹尔塔格组的时代修订为早泥盆世。
关键词:
Abstract
Some new galeaspid specimens have been recently found from the Silurian of Tarim Basin, Xinjiang. The description of these specimens and the new examination of the galeaspid collections render us the following understanding on the morphology and taxonomy of the Silurian galeaspids from Xinjiang. 1) Nanjiangaspis zhangi is removed from the Hanyangaspidae, and referred to a new genus of the Xiushuiaspidae, Xiyuaspis gen. nov.; 2) Platycaraspis tianshanensis is the junior synonym of Microphymaspis pani. According to the principle of priority, Microphymaspis pani is retained, but it is removed from the Xiushuiaspidae, and referred to the Dayongaspidae; 3) the specimen that was referred to the Arthrodira indet. from the Kezirtag Formation of Kalpin, is identified as a new genus and species of galeaspids, Jiaoyu imperfectus gen. et sp. nov.; 4) the fragments and the anterior ventral plate with snowflake-like ornaments previously referred to Hanyangaspis from the Tataertag and Yimugantawu formations probably belong to Nanjiangaspis; 5) the age of the upper member of the Kezirtag Formation is probably Early Devonian as indicated by Jiaoyu imperfectus and the invertebrate fossils from the lower member.
Keywords:
PDF (4934KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
刘玉海, 朱敏, 林翔鸿, 卢立伍, 盖志琨. 新疆塔里木盆地志留纪盔甲鱼类新知. 古脊椎动物学报[J], 2019, 57(4): 253-273 DOI:10.19615/j.cnki.1000-3118.190524
LIU Yu-Hai, ZHU Min, LIN Xiang-Hong, LU Li-Wu, GAI Zhi-Kun.
Galeaspida Tarlo, 1967
Hanyangaspidida P’an et al., 1975
Xiushuiaspidae P’an & Wang, 1983
Xiyuaspis gen. nov.
Etymology Xiyu, the Chinese pinyin (phonetic alphabet) of the ancient name of the Xinjiang Uygur Autonomous Region.
Type species Xiyuaspis zhangi (Lu et al., 2007).
Diagnosis Large-sized xiushuiaspid jawless fishes; subelliptic head-shield, about 110 mm in length; maximum width about equal to length; widest part positioned at about anterior 3/5 of head-shield; median dorsal opening close to rostral margin of head-shield, transversely oval in outline, width/length ratio about 2; orbits small, dorsally positioned, distance between two orbits about equal to distance from each orbit to lateral margin of head-shield; lateral dorsal sensory canals developed, with anterior end almost reaching rostral margin of head-shield; more than 7 pairs of lateral transverse canals, and 2 pairs of medial transverse canals; median dorsal canal absent; ornaments consisting of small round tubercles.
Xiyuaspis zhangi (Lu et al., 2007)
(Figs. 1-3A)
Nanjiangaspis zhangi Lu et al., 2007
Kalpinolepis zhangi Liu et al., 2015
Holotype An incomplete head-shield and its external mould, GMC V 2191a, b.
Paratype An incomplete head-shield, GMC V 2192.
New materials An incomplete head-shield, IVPP V 25445.1a and its external mould IVPP V 25445.1b; a nearly complete ventral rim IVPP V 25445.2; a nearly complete corner IVPP V 25445.3; two incomplete head-shields preserved together on the same rock, GMC V 2416.1, 2.
Type locality and horizon Tielikewatie village, Kalpin County, Xinjiang, China; Tataertag Formation, Telychian, Llandovery, Silurian.
Diagnosis As for genus (monotypic).
Measurements See Table 1.
Ordinis Incerti
Dayongaspidae Pan & Zeng, 1985
Microphymaspis Wang et al., 2002
Type species Microphymaspis pani Wang et al., 2002.
Diagnosis Large-sized dayongaspid jawless fishes; head-shield resembling lateral-view bamboo rain hat, about 200 mm in width, and about 130 mm in length; median dorsal opening close to rostral margin of head-shield, transversely oval in outline, with width/length ratio about 2; orbits round, dorsally positioned, and distance between two orbits larger than distance from each orbit to lateral margin of head-shield; lateral dorsal sensory canals very developed with anterior terminal almost reaching rostral margin of head-shield; lateral transverse canals more than seven pairs, medial transverse canal two pairs; ornaments consisting of fairly small round tubercles.
Microphymaspis pani Wang et al., 2002
(Figs. 3B, 4)
Platycaraspis tianshanensis Wang et al., 2002
Microphymaspis pani Liu et al., 2015
Holotype An incomplete head-shield, IVPP V 13332.
Paratype An incomplete head-shield, IVPP V 13333.
Type locality and horizon Tielikewatie village, Kalpin County, Xinjiang, China; Tataertag Formation, Telychian, Llandovery, Silurian.
Diagnosis As for genus (monotypic).
Order & Famly incerti
Jiaoyu gen. nov.
Etymology Jiaoyu, jiao, corner, the Chinese pinyin for corner; yu, the Chinese pinyin for fish, in reference to the head shield of the fish with a pair of corners.
Type species Jiaoyu imperfectus gen. et sp. nov.
Diagnosis Poorly known galeaspid jawless fish; head-shield about 70 mm in length, and widest part positioned at about 1/2 of head-shield; corner spine-like, issuing lateroposteriorly at about 1/2 of head-shield length; region of head-shield posterior to base of corner long; ornaments consisting of fairly small round tubercles.
Jiaoyu imperfectus gen. et sp. nov.
(Figs. 5A, 7)
Athrodira indet. Lu et al., 2007
Etymology Imperfectus (lat.), incomplete, referring to the poor preservation of specimen.
Holotype An incomplete head-shield GMC V 2195.
Type locality and horizon Kalpin County, Xinjiang, China; Upper Member of Kezirtag Formation, Early Devonian.
DiagnosisAs for genus (monotypic).
1 研究背景
在编写《中国古脊椎动物志·无颌类》有关盔甲鱼亚纲部分的过程中,通过对盔甲鱼类现有标本的系统整理和重新观察,我们获得了一些新的认识,特别是涉及新疆塔里木盆地志留纪的盔甲鱼类,由于标本保存原因,在鉴定上发现不少值得商榷的问题(Liu et al., 2015)。在《关于盔甲鱼类若干问题的讨论》一文中,我们曾对此有过说明(Liu et al., 2014)。本文是前文的续篇,着重讨论我们在对新疆盔甲鱼类的梳理中所获得的新认知,虽然其中个别的认知在上述志书中已有所涉及,但未曾展开讨论。2018年9月,课题组针对新疆柯坪地区的志留系红色岩系又组织了一次新的野外考察,并采获一批盔甲鱼类的新材料,从而进一步佐证了我们一些认知,在此一并描述。在新疆塔里木盆地西北边缘的柯坪、巴楚一带分布有厚达2500 m的中古生代红色岩系,由下而上分为4个岩石地层单位:柯坪塔格组、塔塔埃尔塔格组、依木干他乌组和克兹尔塔格组。曾经由于缺少可靠的化石证据,该红色岩系的地质时代归属一直众说纷纭,莫衷一是。总的来说,在脊椎动物化石发现之前该红色岩系大部归于泥盆纪的不同时期(Liu, 1995; Zhao et al., 2009)。1988年,王朴等报道了柯坪塔塔埃尔塔格组的中华棘鱼(Sinacanthus)棘刺,这是脊椎动物化石在塔里木红色岩系的首次发现(Wang et al., 1988), 据此他们将该组归于中-晚志留世。此前Sinacanthus曾发现于湖北武昌坟头组(原锅顶山组)、皖北坟头组,并被归入棘鱼纲(P’an, 1957, 1959; Pan, 1986a, b), 后来朱敏根据组织学等方面的研究认为Sinacanthus及形态上与其相近的Neosinacanthus和Tarimacanthus均属软骨鱼纲,同时朱敏将这3个属归入一新科——中华棘鱼科Sinacanthidae (Zhu, 1998)。
继王朴等之后,刘时藩分别在1993和1995年对产自塔里木红色岩系的Sinacanths进行了观察和研究,发现它们也存在于依木干他乌组,从而认为该组时代亦属志留纪温洛克世;与此同时,刘氏更进一步指出Sinacanthus在塔里木红色岩系的出现,更重要的地质意义在于其所透露出的志留纪时期的中国古动物地理信息(Liu, 1993, 1995)。刘时藩(1995)总结了中国地区性鱼类中华棘鱼类的地史分布,和其相伴出现的盔甲鱼类汉阳鱼(Hanyangaspis)、大庸鱼(Dayongaspis)同样也是中国地区性的鱼类,二者广泛分布于扬子地区志留纪温洛克世和兰多维列世地层中。结合当时王俊卿等对塔里木红色岩系中脊椎动物化石的初步研究结果——塔塔埃尔塔格组也产盔甲鱼类Hanyangaspis (其报告发表于1996年), 刘氏认为塔里木红色岩层中的脊椎动物Hanyangaspis-Sinacanthus组合与扬子地区志留纪时期的脊椎动物组合是相似的,当时这两个地区是彼此邻近或相连的同一个生物地理区系。为了佐证这点,刘氏将构成中国大陆的主要构造单元的地层架构进行了比较,认为其中扬子地区和塔里木地区在古生代时期的地层框架最为相近,同样表明当时在地理上它们是临近或相连的;至于今天两地相隔遥远,则是后来构造错动的结果(Liu, 1995)。王俊卿等(1996)依据对塔里木红色岩系的脊椎动物化石,特别是盔甲鱼类的研究,也认为这些化石对塔里木中古生代古地理的分析意义重大,“华南与塔里木在志留纪时有可能同属一个陆块”。此后,赵文金等(2005, 2009)先后分别主要根据当时的盔甲鱼类研究成果或侧重于新疆塔里木地区志留纪时期含鱼层序列所作分析,同样认为在中古生代塔里木板块与华南板块属于同一个生物区系且彼此有着密切联系。此后越来越多的研究佐证了这一说法,盖志琨等(2018)报道了在这一地区塔塔埃尔塔格组中发现的最早的多鳃鱼类锯齿宽腹鱼(Platylomaspis serratus), 该种与华南地区的曾氏南宁鱼(Nanningaspis zengi)存在较近的亲缘关系,再次印证了塔里木板块与华南板块关系密切。综上所述,塔里木地区的盔甲鱼类对含鱼层时代的确定和该地区古地理的恢复具有重大意义。然而,在系统古生物学方面,由于迄今所获得的塔里木盔甲鱼类标本大多破碎、残缺不全,很难对其准确鉴定,仍在在某些方面存在很多令人困惑的问题,致使有些鉴定需要重新厘定。
2 标本记述
盔甲鱼亚纲 Galeaspida Tarlo, 1967汉阳鱼目 Hanyangaspidida P’an et al., 1975
修水鱼科 Xiushuiaspidae P’an & Wang, 1983
西域鱼属 Xiyuaspis gen. nov.
词源 Xiyu, 新疆古称,西域的汉语拼音。
属型种 张氏西域鱼Xiyuaspis zhangi (Lu et al., 2007) (图1-3A)。
图1
 新窗口打开|下载原图ZIP|生成PPT
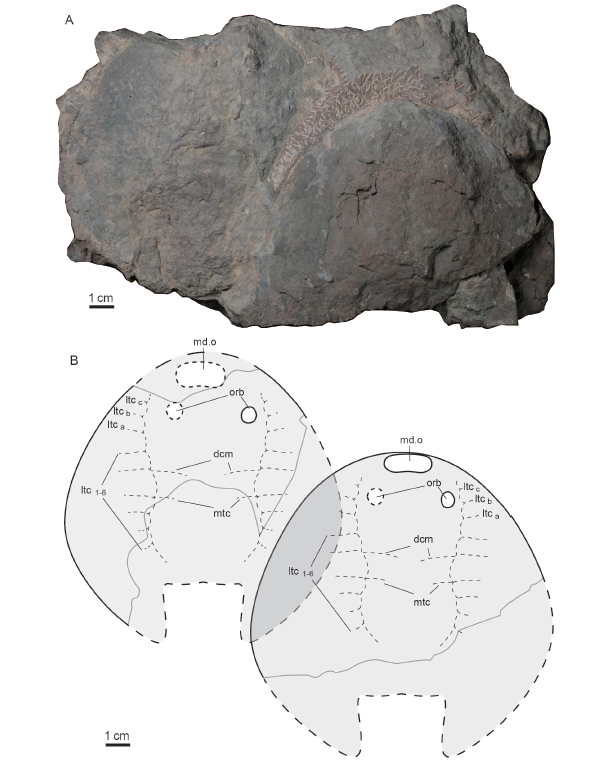
新窗口打开|下载原图ZIP|生成PPT图1张氏西域鱼新增标本IVPP V 25445.1-3照片及其解释性素描
Fig. 1Photographs of Xiyuaspis zhangi IVPP V 25445.1-3 and their interpretative drawingA. an incomplete head-shield (V 25445.1): A1, A2. internal mould and its interpretative drawing
(V 25445.1a), A3. close-up of granulous ornamental tubercles (box region of A1), A4, A5. external mould and its interpretative drawing (V 25445.1b); B. a nearly complete ventral rim (V 25445.2);C. a nearly complete corner (C1) and its interpretative drawing (C2) (V 25445.3)
Abbreviations: c. corner 角;ltca-c. the first to third lateral transverse canals issuing from the infraorbital canal 从眶下管发出的第一至三对侧横管;ltc1-3. the first to third lateral transverse canals issuing from the lateral dorsal canal 从侧背管发出的第一至三对侧横管;md.o. median dorsal opening 中背孔;orb. orbital opening 眶孔;vr. ventral rim 腹环
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2张氏西域鱼新增标本GMC V 2416.1, 2照片(A)及其解释性素描(B)
Fig. 2Photograph of Xiyuaspis zhangi GMC V 2416.1, 2 (A) and its interpretative drawing (B)
图3
 新窗口打开|下载原图ZIP|生成PPT
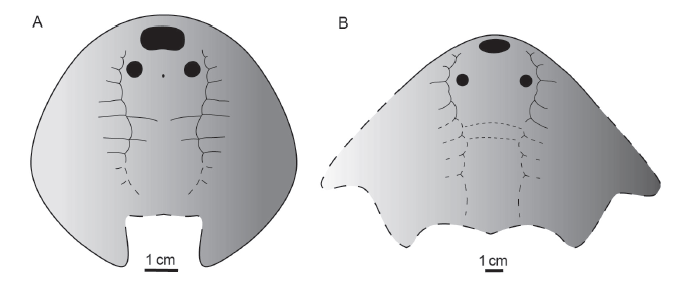
新窗口打开|下载原图ZIP|生成PPT图3张氏西域鱼(A)与潘氏小瘤鱼(B)复原
Fig. 3Restorations of Xiyuaspis zhangi (A) and Microphymaspis pani (B)
特征 体形大的修水鱼类,头甲呈亚椭圆形,长约110 mm, 最宽部位约位于头甲前3/5处,与头甲长大致相等;中背孔贴近头甲前缘,呈横置椭圆形,宽约为长的两倍;眶孔背位,眶间距与眶孔至头甲侧缘的距离相等;侧背纵管发达,前端几乎达头甲吻缘,侧横管7对以上,中横管2对,中背纵管缺失;纹饰由细小的粒状突起组成。
比较与讨论 属型种Xiyuaspis zhangi是西域鱼属目前所知的唯一种。由于该种建立时所依据的头甲边缘大部缺失,因而关乎其系统位置至关重要的头甲形状不能确定,以致该种最初归于汉阳鱼科(Hanyangaspidae)的南疆鱼属(Nanjiangaspis, Lu et al., 2007)。然而,其头甲纹饰是由细小粒状突起组成,而Nanjiangsapis的纹饰不仅大,而且呈雪花状,由于这方面的显著不同,因而后来被归入纹饰同样为粒状突起的柯坪鱼属(Kalpinolepis, Liu et al., 2015), 后面这个属也归属汉阳鱼科(Wang et al., 1996)。现在新增加的材料证实该物种的头甲呈椭圆形确定无疑。目前汉阳鱼目包括汉阳鱼科和修水鱼科(Xiushuiaspidae)两个科,二者的区别主要在于前者头甲呈前窄后宽的梯形,后者头甲为竖置的椭圆形(Zhu and Gai, 2006; Liu et al., 2015), 因此该种无疑当归于修水鱼科,并为其建立新属西域鱼属Xiyuaspis gen. nov.。目前所知,修水鱼科包括两个属:修水鱼(Xiushuiaspis)和长兴鱼(Changxingaspis), 分别产自华南的志留纪兰多维列世特列奇期的江西西坑组和浙江茅山组(P’an and Wang, 1983; Wang, 1991)。虽然出自塔里木红色岩系的小瘤鱼属(Microphymaspis)也曾被归入修水鱼科(Wang et al., 2002), 但经厘定被归入大庸鱼科(Dayongaspidae, Liu et al., 2015, 本文)。新属西域鱼与修水鱼、长兴鱼之不同是显著的:首先是体型硕大,头甲长达110 mm, 而在后二者则分别为25和35 mm; 在头甲的长与宽比例上,西域鱼宽约与长相等甚或宽略大于长,而后二者则宽远小于长;就中背孔而言,虽然该孔在三者都是接近头甲吻缘并且宽大于长,但在西域鱼中该孔为椭圆形,宽与长之比为2:1, 而在修水鱼和长兴鱼中,该孔近于裂隙状,宽与长之比分别为4:1和5:1。西域鱼在头甲后部的轮廓可能更趋向于长兴鱼,两者的角均较大,而修水鱼的角非常小,近于消失。
张氏西域鱼 Xiyuaspis zhangi (Lu et al., 2007)
(图1-3A)
Nanjiangaspis zhangi Lu et al., 2007
Kalpinolepis zhangi Liu et al., 2015
正模 一件头甲的内、外模。中国地质博物馆脊椎动物化石编号GMC V 2191a, b。
副模 一件不完整头甲内模。中国地质博物馆脊椎动物化石编号GMC V 2192。
新增标本 一件不完整头甲的内、外模,一件右半部分保存头甲腹环和一件近于完整保存的角,中国科学院古脊椎动物与古人类研究所脊椎动物化石编号IVPP V 25445.1a, b, V 25445.2, V 25445.3。一件近于完整保存的头甲内模和一件右侧边缘大部保存的头甲内模,中国地质博物馆脊椎动物化石编号:GMC V 2416.1, 2, 两者共同保存在同一标本上,后者轻微覆压在前者之上。
产地与层位 新疆柯坪铁热克阿瓦提村,志留系兰多维列统特列奇阶塔塔埃尔塔格组。
特征 唯一的种,特征从属。
测量 见表1。
Table 1
表1
表1张氏西域鱼标本测量
Table 1
| Holotype | GMC V 2416.1 | GMC V 2416.2 | IVPP V 25445.1 | |
|---|---|---|---|---|
| Maximum length of preserved head-shield | 76.0 | 111.0 | 90.5 | 92.0 |
| Maximum width of preserved head-shield | 72.0 | 115.0 | 115.0 | 101.6 |
| Length of short axis of orbital opening | 7.0 | 6.4 | 7.2 | 6.0 |
| Length of long axis of orbital opening | 10.0 | 7.1 | 7.8 | 7.6 |
| Distance between orbital openings | 24.0 | — | 21.0 | 22.5 |
| Length of short axis of median dorsal opening | 11.0 | — | 11.0 | 10.6 |
| Length of long axis of median dorsal opening | 20.0 | — | 22.5 | 21.0 |
| Length of short axis of pineal fossa | 1.3 | — | — | — |
| Length of long axis of pineal fossa | 2.0 | — | — | — |
新窗口打开|下载CSV
描述 新增标本IVPP V 25445.1a眶孔两侧保存完好(图1A), GMC V 2416.1头甲于眶孔以前的部分大部缺失,眶孔以后头甲左侧部分基本保存,GMC V 2416.2则头甲眶前部保存完整(图2), 三者共同显示头甲呈亚椭圆形,而非原来描述的梯形。头甲长约110~115 mm, 其最大宽度约与长度相等,位于头甲的前3/5处。头甲吻缘在IVPP V 25445.1a, GMC V 2191a, GMC V 2192和GMC V 2416.2均有保存,较圆钝;头甲侧缘在标本IVPP V 25445.2和GMC V 2416.1保存甚好,呈深度弓形弯曲,于头甲的前3/5处达到弯曲的顶点,即达到头甲最宽处,宽110~115 mm。头甲侧缘后段如同修水鱼和长兴鱼,由最宽处顶点向后,逐渐向内弯曲,暗示内角弯曲(图1-3)。内角在标本IVPP V 25445.3中独立保存(图1C), 长度约30 mm, 为自然边缘,表明西域鱼的内角与长兴鱼的比较相似,较大,呈肥大叶状,且有一定的弧度。
中背孔在正型标本和新增标本IVPP V 25445.1a和GMC V 2416.2中均有不同程度的保存,而以V 25445.1a保存最完好,贴近头甲吻缘,略呈横置亚椭圆形,向前凸,后缘在GMC V 2192保存较好,微凹、近于平直,长轴长20~23 mm, 短轴长约11 mm。眶孔在IVPP V 25445.1a, GMC V 2191a, V 2192保存清晰而未变形,背位,眶孔呈椭圆形,长轴长7~10 mm, 短轴长6~7 mm, 两眶孔的眶间距21~24 mm, 略大于中背孔之宽,眶孔前缘连线与中背孔后缘之距离约等于眶孔的直径,头甲于眶孔内侧隆起为低矮的短脊,从而眶间区相对低平,而眶孔朝向侧背方。V 2191a显示出松果穴,位于眶孔之间。
侧线感觉管在正型标本GMC V 2191a与新增标本IVPP V 25445.1a, b保存很好(图1A1, A4), GMC V 2192亦有部分保存,其中侧背管发达,前端始于中背孔侧面靠近吻缘,保存有最前面7支侧横管;中横管2支(对), 其中前面的1支与其他盔甲鱼类的背联络管(dcm)相当,因此6支侧横管位于联络管之前;但是GMC V 2191a头甲保存部分长仅76 mm, 而GMC V 2416.1头甲长达约110 mm, 因此在GMC V 2191a头甲后部缺失可能长达约30 mm。与此相关,现有保存的最后一支侧横管之后,可能大约有2支侧横管因头甲的缺失而未保存;中背管不存在。
头甲腹面了解甚少,仅标本IVPP V 25445.2近于完整保存了头甲右侧腹环(图1B), 标本GMC V 2416.1头甲左侧的腹环有部分出露(图2A), 腹环由前向后增宽,至头甲前2/3处,该侧腹环所暴露出的部分,其宽超过40 mm, 超过头甲宽的1/3, 这指示西域鱼可能像修水鱼和长兴鱼那样,两侧腹环后端向中线凑合,形成封闭的口鳃窗,口鳃区局限在头甲的前部,而鳃后区伸长。
纹饰由粒状瘤点组成,瘤点细小、密集、分布均匀、互不融合,每平方毫米约有5~6个瘤点(图1A3)。
目未定 Incerti Ordinis
大庸鱼科 Dayongaspidae Pan & Zeng, 1985
小瘤鱼属 Microphymaspis Wang et al., 2002
属型种 潘氏小瘤鱼Microphymaspis pani Wang et al., 2002。
特征 体形硕大的大庸鱼科鱼类,头甲呈侧视斗笠状,宽约达200 mm, 长估计约130 mm。中背孔近头甲吻缘,呈横置的椭圆形,宽约为长的2倍;眶孔圆形、背位,位于中背孔后侧方,眶间距大于眶孔至头甲侧缘的距离;侧线系统中,侧背管发达,前端几乎达到头甲吻缘,侧横管7支以上,其中6支位于背联络管之前;纹饰为粒状突起,细小而密集。
比较与讨论 潘氏小瘤鱼(Microphymaspis pani)建立时归入修水鱼科(Wang et al., 2002), 刘玉海等(2015)厘定其隶属大庸鱼科;原作者对该属种系统位置的误判,缘于误认为正型标本在右侧眶孔之外尚保存有左侧眶孔,导致对头甲侧缘走向和头甲形状的误判,从而认为头甲为椭圆形,实则是像大庸鱼科的大庸鱼(Dayongaspis)那样,头甲呈侧视的斗笠形。小瘤鱼虽然和大庸鱼同样头甲呈侧视斗笠状,但前者远大于后者,就头甲宽而言前者约为后者的2倍;小瘤鱼的中背孔呈横置的椭圆形,而大庸鱼的中背孔则小而圆;小瘤鱼的眶孔远离头甲中线,因此眶间距大于眶孔至头甲侧缘的距离,大庸鱼的眶孔则小而靠近中线,与中背孔一起形成紧凑的“品”字形。
潘氏小瘤鱼 Microphymaspis pani Wang et al., 2002
(图3B, 4)
Platycaraspis tianshanensis Wang et al., 2002
Microphymaspis pani Liu et al., 2015
正模 一件不完整的头甲,IVPP V 13332。
副模 一件头甲残片的外模,IVPP V 13333。
产地与层位 新疆柯坪铁热克阿瓦提村,志留系兰多维列统特列奇阶塔塔埃尔塔格组。
特征 唯一种,特征从属。
描述 本文纳入潘氏小瘤鱼的标本,除了这个属种初建时所依据的正模IVPP V 13332之外(图4A), 另一件是被本文归入的标本IVPP V 13333 (图4B), 该标本曾被作为天山宽头鱼(Platycaraspis tianshanensis)正型标本,并被归入大庸鱼科(Wang et al., 2002)。将标本V 13333归入潘氏小瘤鱼的根据有四:其一,V 13333与潘氏小瘤鱼的正模V 13332出自同一产地与层位;其二,二者头甲均显示异于寻常的宽大,尤其是头甲侧缘走向有着相似的轨迹——侧展、微弯曲;其三,二者的纹饰相同,均由细小粒状瘤点组成;其四,二者现有保存的侧线相互吻合。因此依据标本V 13333所建立的宽头鱼(Platycaraspis)与依据标本V 13332所建立的小瘤鱼(Microphymaspis)应属于同一个属。虽然天山宽头鱼(P. tianshanensis)和潘氏小瘤鱼(M. pani)二者建立时刊出于同一篇论文,但前者排序在后,故天山宽头鱼应是潘氏小瘤鱼的次异名。
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4潘氏小瘤鱼化石照片及其解释性素描
Fig. 4Photographs of Microphymaspis pani and their interpretative drawings
A. photograph of an incomplete head-shield (A1) and its interpretive drawing (A2), IVPP V 13332, holotpye;B. photograph of an incomplete head-shield (B1) and its interpretive drawing (B2), IVPP V 13333, paratype
Abbreviations: ldc. lateral dorsal canal 侧背管;ltc. lateral transverse canal 侧横管;md.o. median dorsal opening 中背孔;mtc. medial transverse canal 中横联络管;orb. orbital opening 眶孔
正型标本V 13332仅保存头甲的右侧前部,包括头甲右侧的部分前缘和侧缘、中背孔的右侧部分、右侧的眶孔及部分侧线。头甲的吻缘和侧缘走向作侧向伸展并呈略微弯曲的弧形,而非原作者认为的作后向伸展、呈深弧形弯曲(Wang et al., 2002)。在侧缘后部缺失的情况下,头甲右侧保存宽度,即现存侧缘末端至头甲中线,已达65 mm, 可以想象完整头甲之宽必定可观;标本V 13333为头甲右侧的部分自然外模,包括头甲右侧的部分侧缘、侧线感觉管和残缺的眶孔。就侧缘而言,如同V 13332, 侧向伸展、微弯曲,虽然也未达其自然末端,但其后部较V 13332保存得较多,其宽度很可能超过200 mm, 因此与已知体形最大的盔甲鱼类如汉阳鱼(Hanyangaspis, Pan, 1986a, b)、班润鱼(Bannhuanaspis, Janvier et al., 1993)、东方鱼(Dongfangaspis, Liu, 1975)和箭甲鱼(Antiquisagittaspis, Liu, 1985)等可以匹敌甚或过之。然而,由于现有标本中头甲后部的缺失,因此头甲长度、后缘和内角的形状均不详。
中背孔仅部分右侧在V 13332中有所保存(图4A1, A2), 接近吻缘,横置椭圆形,估计宽约为长的2倍;眶孔中等大小,背位,在中背孔的侧后方,与中背孔侧端的垂直线之距约等于眶孔的直径,与中背孔后缘的水平线之距亦约等于眶孔的直径,眶间距大于眶孔与头甲侧缘间的距离。
侧线感觉管在标本V 13332仅侧背管的前部保存,但甚清晰,其前端止于中背孔侧方,未达头甲吻缘,由此向侧后方伸延,至眶孔侧面则折向中后方,由于侧背管后部的缺失,因此仅保存其最前面的4支侧橫管,由前而后长度递增,其中前2支位于眶孔之前,而第3支位于眶孔侧方,其出发点正处于侧背管走向由侧后方折向中后方的转折点,第4支则位于眶孔之后;见之于大庸鱼中的前眶上管在这里不存在。在标本V 13333中感觉管也很清晰,但是由于头甲前部和后部均缺失,侧背管只保存中段部分。由这段侧背管发出的最前面的1支侧横管仅残留近端部分,值得注意的是,其位置与标本V 13332中的第3支侧横管相似,也恰在眶孔侧面,并处于侧背管的走向由侧后方折向中后方之转折点上,据此,该侧横管当与标本V 13332中的第3支侧横管相对应。在此侧横管之前,标本V 13333由于头甲前部缺失,侧背管仅有小段保存;在此侧横管之后,尚有2支侧橫管保存下来,再稍后,可观察到侧背管的内侧残存一段中横管(mtc)。综合上述两件标本的侧线管的描述可以归纳为潘氏小瘤鱼在中横管之前发育5支侧横管,其中2支位于眶孔之前,1支位于眶孔之侧,2支位于眶孔之后,至于中横管之后,则因头甲缺失而未有侧横管保存。在湖南大庸鱼(Dayongaspis hunanensis)、锅顶山汉阳鱼(Hanyangaspis guodingshanensis)以及顾氏长兴鱼(Changxingaspis gui)等盔甲鱼基干类群中(Liu et al., 2015: 图79C, 66C, 78A), 通常具有2支前后相邻的中横管,其中后1支与多鳃鱼类(Liu et al., 2015: 图109A, 114D)和华南鱼类(Liu et al., 2015: 图125C, 126B)的背联络管(dcm)同源,而侧横管则从位于背联络管之前的2支起,由前而后依次与多鳃鱼类和华南鱼类第1, 第2, 第3......等侧横管(ltc1, 2, 3......)同源;在汉阳鱼和大庸鱼等盔甲鱼基干类群里,在第1侧横管(ltc1)之前尚具若干侧横管,数目因物种不同而异,这些侧横管排序则由后向前名之为侧横管a, b, c......(ltca, b, c......)。依据以上侧线同源关系的陈述,可以推测潘氏小瘤鱼的背联络管在现有标本中没有保存,而中横管之前的1支侧横管,即V 13333中现保存的最后1支,当与第1侧横管同源,而V 13332中所保存的4支侧横管,则由后而前依次为ltca-d。而在上述3个盔甲鱼基干类群里,自背联络管向后尚具侧横管2支以上,多达4支,其中湖南大庸鱼头甲相对宽但短,仅约2支,潘氏小瘤鱼的情况与其近似,可能也约2支。
头甲纹饰在两件标本中均有保存,由粒状瘤点组成,突起细小,每平方毫米有6~7个瘤点,排列紧密但彼此不融合。
盔甲鱼亚纲 Galeaspida Tarlo, 1967
目和科未定 Incerti ordinis et incertae familiae
角鱼属 Jiaoyu gen. nov.
词源 jiaoyu, 角鱼,汉语拼音,有角的鱼。
属型种 残缺角鱼 Jiaoyu imperfectus gen. et sp. nov.。
特征 了解甚少的盔甲鱼;头甲长约70 mm, 最宽部位在头甲中部;角呈狭长棘状,指向后侧方,角的位置靠前,头甲角后区长;头甲前侧缘微弓曲,向后侧伸延,后侧缘走向则向后但略朝中线倾斜;纹饰由细小突起组成。
比较与讨论 本文建立的残缺角鱼(Jiaoyu imperfectus gen. et sp. nov.)所依据的标本GMC V 2195 (图5), 曾被认为属节甲类或长胸节甲类的前腹侧片+棘片(avl+sp), 并引起关于含鱼层时代的争论(Lu et al., 2007)。此标本骤然看起来与包括节甲类在内的某些盾皮鱼类的avl+sp颇为相似(图6A, B), 但仔细观察乃是盔甲鱼类发育有角的头甲左侧碎片,即头甲的角(被误判为棘片)和与角相连的部分头甲本体碎片(被误判为前腹侧片)。事实上,现有标本的头甲本体部分除了其侧缘为自然边缘外,前缘、内缘和后缘均参差不齐,并不是自然边缘,表明它只不过是头甲左侧腹环及角的一部分,与盾皮鱼类前腹侧片的某种相似只是巧合。众所周知,盔甲鱼类头甲的角乃是头甲侧缘向外突伸部分,是头甲本体的延续,因此在纹饰分布上,二者间也是连续无间的,显然上述标本正是这一情况。然而前腹侧片和棘片则是构成盾皮鱼类的躯甲的诸多甲片里的两件,和其他甲片一样,这两件甲片也是各自分别产生的,因此它们之间虽然连接紧密(与其他甲片相比更经常牢固连接在一起), 但其间的界线通常依然可寻。现以盾皮鱼类中的节甲类(图6A, B)、瓣甲类(图6C)为例,三者的棘片+前腹侧片在形态上与GMC V 2195标本看起来最为相似而易于混淆。从图中可以看出,前腹侧片的纹饰无论是粒状疣突还是条状脊纹,都是以前腹侧片生长中心为中心,作不同程度的同心圆或放射状分布(图6B, C), 而棘片的纹饰则是作大致与棘片边缘平行分布(图6B)。因此,从纹饰上这两件甲片的界线依稀可见。另一方面,在没有受到严重挤压的情况下,前腹侧片通常存在一个以该甲片生长中心为顶角、胸窦部位为底边的三角形凹陷。显然,这两个特征在标本GMC V 2195中均不具备,因此,以上证据足以说明该标本排除属于盾皮鱼类的avl+sp的可能性。
图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5残缺角鱼(新属新种,正型标本GMC V 2195) (A)和让氏昭通鱼(IVPP V 25965) (B)化石照片
Fig. 5Photographs of Jiaoyu imperfectus gen. et sp. nov., holotype, GMC V 2195 (A) and Zhaotongaspis janvieri, IVPP V 25965 (B)
Abbreviations: c. corner 角;ltc. lateral transverse canals 侧横管;vr. ventral rim 腹环
图6
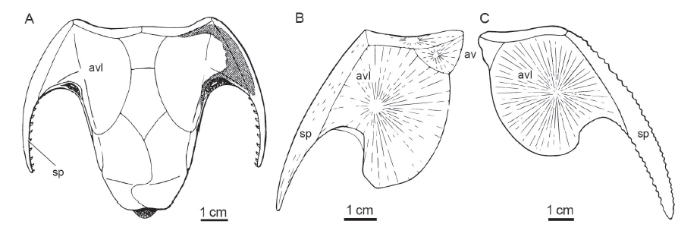 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6盾皮鱼类中的前腹侧片+棘片(avl+sp)
Fig. 6The anterior ventrolateral plate (avl) plus spinal plate (sp) in placoderms, ventral view
A. arthrodire Dicksonosteus arcticus from Early Devonian of Spitsbergen (modified from Goujet, 1984);B. arthrodire Kujdanowiaspis sp. (modified from Denison, 1958);C. petalichthyid Quasipetalichthys haikouensis (modified from Liu, 1991). av. anterior ventral plate 前腹片
由于保存的原因,角鱼在盔甲鱼类中的分类位置目前还不明确。依据角呈长棘状、位置甚是靠前和头甲具长的角后区等特征,首先当将其排除在盔甲鱼类基干类群汉阳鱼目(包括汉阳鱼科和修水鱼科)和繁盛于早泥盆世的多鳃鱼目之外,因为二者均不存在角。除上述两个目之外,角广泛出现在盔甲鱼的其余类群中。首先就大庸鱼科而言,如大庸鱼(Pan and Zeng, 1985)、小瘤鱼(图3B)的角不但极短而壮,而且硕大内角紧随其后并构成头甲后端,因此角鱼与以上两属迥然不同,排除了隶属大庸鱼科的可能性。至于真盔甲鱼目(Eugaleaspdiformes), 其现有两个科之一的中华盔甲鱼科(Sinogaleaspidae)目前只发现于志留系的茅山组,所含4个属中华盔甲鱼(Sinogaleaspis)、曙鱼(Shuyu)、安吉鱼(Anjiaspis)、煤山鱼(Meishanaspis)形态分异度甚小,其角不但处于头甲最后部位、头甲不具角后区,而且内角形如短棘,附于角的内侧(Pan and Wang, 1980; Wang, 1991; Gai and Zhu, 2005; Gai et al., 2005, 2011)。因此,角鱼与该科所有已知属明显有别,无疑无缘隶属该科。至于出现于志留系罗德洛统至下泥盆统的真盔甲鱼科(Eugaleaspidae), 形态分异度较大,可分为3种情况:其中如真盔甲鱼(Eugaleaspis)和盾鱼(Dunyu), 其角呈宽大的三角形、且处于头甲末端,从而完全无角后区(Zhu et al., 2012; Liu et al., 2015: 图88, 91), 角鱼显然与之相距甚远;另一情况见之于云南盔甲鱼(Yunnanogaleaspis)和憨鱼(Nochelaspis), 角与内角由外而内并列、构成头甲后端(Pan and Wang, 1980; Zhu, 1992), 这一情况显然也不为角鱼所有;第三种情况见之于布拉格期的翼角鱼(Pterogonaspis)和三尖鱼(Tridensaspis) (Liu, 1986; Zhu, 1992), 与角鱼类似,它们的角也呈长棘状,并具明显的角后区,因此角鱼是否与它们有较近的关系也是须考虑的可能性之一。除以上分析过的盔甲鱼成员外,在现有的盔甲鱼类分类系统中,余下的尚有华南鱼目(Huananaspiformes)和华南鱼目外类群昭通鱼科(Zhaotongaspidae), 二者也皆普遍发育有角。就华南鱼目而言,所含种类繁多,形态分异度大,其中三岐鱼科(Sanqiaspidae)和鸭吻鱼科(Gantarostrataspidae)成员的头甲狭长、两侧缘近于平行,角则处于头甲的极后端、完全无角后区 (Liu, 1975; Wang and Wang, 1992; Wang et al., 1996), 因此角鱼与这两个科存在较近关系的可能性不大;大窗鱼类(Macrothyraspinae)则由于在头甲背面存在一对窗(Pan, 1992), 从而排除其与角鱼具较近关系的可能,因为无迹象显示后者存在该构造,而且大多成员角的位置甚为靠后且接近头甲后缘。华南鱼目中只有部分分子类似角鱼,具有位置靠前的角和长的角后区,其中包括三岔鱼科(Sanchaspidae) (如三岔鱼Sanchaspis和箭甲鱼Antiquisagittaspis) (Pan and Wang, 1981; Liu, 1985)和南盘鱼科(Nanpanaspidae) (如南盘鱼Nanpanaspis) (Liu, 1975; Liu et al., 2018); 但是从属一级阶元来说,角鱼与这些属明显有别,从而不属于它们中的任何属,例如箭甲鱼和南盘鱼的角呈既短又壮的三角形,三岔鱼的角短而弯曲。从角的形态、伸出位置判断,角鱼与昭通鱼科的昭通鱼最为接近,也是角前位、头甲角后区长(图5A, B)。两个属种在保存相同部位的两块标本中,均保存了同样的两条侧横管(ltc, 图5A, B)。另外,两者的纹饰也较为相似,均为细小的粒状瘤点。但两者仍有非常明显的差异,昭通鱼的角内缘饰以齿状小刺,并且其角的内缘与头甲后侧缘的夹角为55°~65°, 而在角鱼则为80° (图5A, B)。因此,Jiaoyu作为一个新属是成立的。
至于角鱼与哪个属亲缘关系最近,由于现有标本保存方面的局限,对于确定其系统位置至关重要的特征,诸如吻突存在与否、中背孔形状、眶孔位置和侧线等均不详,只能作某些可能性的推测。角鱼的中背孔如果呈纵长裂隙状,则可能与翼角鱼比较相似,而归到真盔甲鱼目真盔甲鱼科(图7C); 角鱼的中背孔如果呈椭圆形,同时具有吻突,则可能与三岔鱼比较相似,可能隶属华南鱼目三岔鱼科(图7D); 如果不具吻突,则与昭通鱼较近,从而隶属昭通鱼科(图7B)。从现有材料看其可能的归属应是三岔鱼科或昭通鱼科,究竟是二者中的前者还是后者,则有待新的材料证明角鱼是否具有吻突。
图7
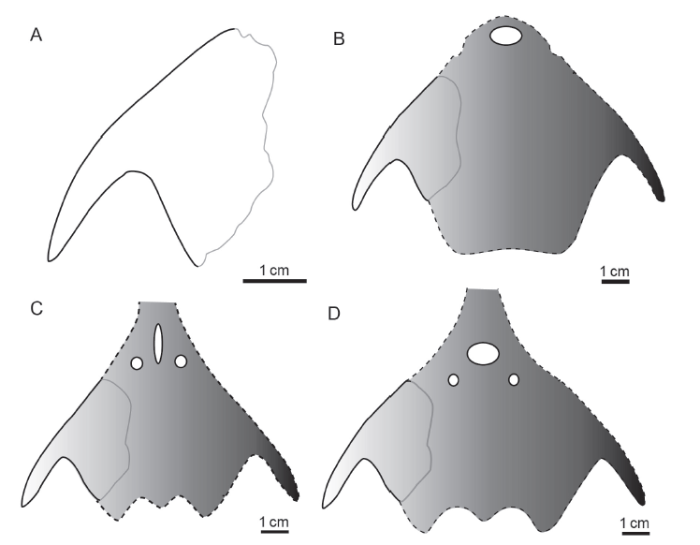 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7残缺角鱼(新属新种)及其可能的头甲复原
Fig. 7Jiaoyu imperfectus gen. et sp. nov. and possible restorations
A. interpretative drawing of holotype, GMC V 2195; B. restoration of the head-shield based on zhaotongaspid Zhaotongaspis, in dorsal view; C. restoration of the head-shield based on eugaleaspid Pterogonaspis, in dorsal view; D. restoration of the head-shield based on sanchaspid Sanchaspis, in dorsal view
残缺角鱼 Jiaoyu imperfectus sp. nov.
(图5A, 7)
Athrodira indet. Lu et al., 2007
词源 imperfectus (第一、第二变格法形容词), 不完全的,残缺的,归因于该鱼保存不完整。
正模 头甲右侧局部自然外模,中国地质博物馆标本登记号GMC V 2195。
产地与层位 新疆柯坪铁热克阿瓦提村,下泥盆统克兹尔塔格组上段。
特征 唯一种,特征从属。
描述 正模GMC V 2195乃现有唯一标本,系头甲右侧的部分外模残片,包括右侧完整保存的角及角之前和之后头甲的部分侧缘,除此之外,其余边缘皆系头甲破碎所致,而非自然边界。头甲保存长度40 mm, 估计其长度约达60 mm。角呈狭长的棘状,指向侧后方,由基部至末端,角长25 mm。角之前头甲前侧缘仅部分保存,保存长度约25 mm, 该侧缘微弓曲,向后侧方伸延;角之后的头甲后侧缘也只部分保存,但长度已达20 mm, 表明头甲具有明显长的角后区,换言之,角的位置明显靠前。头甲后侧缘的走向是向后而略朝中线倾斜,其与头甲前侧缘呈近75°夹角;从头甲现有保存部分和后侧缘的走向推测,头甲应不具内角;头甲边缘保存了两条侧横管的印痕(ltc, 图5A); 纹饰由粒状突起组成,突起细小,分布均匀,每平方毫米有6~7个瘤点。
3 讨论
3.1 塔里木红色岩系是否产汉阳鱼尚无实证
王俊卿等(1996)将产自柯坪塔塔埃尔塔格组和巴楚依木干他乌组的盔甲鱼类头甲散落的若干镶嵌片,以其具雪花状纹饰,而鉴定为锅顶山汉阳鱼(Hanyangaspis guodingshanensis)。锅顶山汉阳鱼最初发现于湖北汉阳锅顶山组(现已归并到坟头组)。因此,锅顶山汉阳鱼镶嵌片的发现成为当时盔甲鱼类在华南与塔里木两个地区之间唯一的共有属种。然而,王俊卿等(2002)在塔塔埃尔塔格组则发现了同样具雪花状纹饰的头甲,并被订为汉阳鱼科一新属种柯坪南疆鱼(Nanjiangaspis kalpinensis), 因此将上述镶嵌片归于该新属种,从而否认了在产上述镶嵌片相同或相当层位找到过汉阳鱼头甲。实际上,迄今所知雪花状纹饰在盔甲鱼类中除了存在于锅顶山汉阳鱼和柯坪南疆鱼外,还见之于湖北京山纱帽组(=坟头组)的锅顶山汉阳鱼相似种(Hanyangaspis cf. H. guodingshanensis, Pan, 1986a, b)、安徽巢湖坟头组的巢湖宽吻鱼(Latirostraspis chaohuensis, Wang et al., 1980; Pan, 1986a, b)、陕西紫阳吴家河组顶部C. lapworthi笔石带的一具雪花状纹饰、属种未定的汉阳鱼类的残余头甲(Fu and Song, 1986)。由此可见,在当前的研究尚未达到在不同属种间的雪花状纹饰作出区分的情况下,单独凭借雪花状镶嵌片判断其所属属种是靠不住的,除非同层位还存在具雪花状纹饰的头甲。因此当前来说,我们认为王俊卿等将前述鉴定为锅顶山汉阳鱼的镶嵌片更正为归属于柯坪南疆鱼是可取的。另一方面,虽然并不是所有汉阳鱼科成员都具有雪花状的纹饰,如在大眼锥角鱼(Konoceraspis grandoculus)则为粒状疣突(Pan, 1992), 但尚未发现雪花状纹饰存在于汉阳鱼科以外的盔甲鱼。归纳以上所列具雪花状纹饰的汉阳鱼类,就其层位来说,在塔里木出现于塔塔埃尔塔格组至依木干他乌组,在华南则产自坟头组至吴家河组顶部C. lapworthi带。因此,在一定范围内雪花状镶嵌片对于其产出层位的时代鉴定还是有帮助的。此外,还有一件产自柯坪塔塔埃尔塔格组的前腹片,也具雪花状纹饰,该片被卢立伍等(2007)鉴定为Hanyangaspis sp.。目前,我们对盔甲鱼类头甲腹面的情况了解并不充分,就现有的资料所知,一共可以区分出三种类型。其一,见之于廖角山多鳃鱼(Polybranchiaspis liaojiaoshanensis), 口鳃窗仅为一件腹片所覆盖,腹片远小于口鳃窗,略呈矩形,而前、后缘凹进(Liu, 1965, 1975); 其二,见之于盾状五窗鱼(Pentathyraspis pelta), 腹片与腹环愈合,从而完全覆盖了口鳃窗,仅留有口孔和鳃孔(Pan, 1992), 上述两属种均隶属繁盛于早泥盆世的多鳃鱼目;其三,见之于锅顶山汉阳鱼和巢湖宽吻鱼(Pan, 1986a, b; Wang, 1986), 其口鳃窗由前后相邻的前腹片和腹片覆盖,前者覆盖口鳃窗的口区,后者覆盖鳃区,这两个属种均产于志留纪兰多维列世坟头组,隶属于盔甲鱼类基干类群汉阳鱼目汉阳鱼科。
在这三种类型中,前两种应是由后一种衍生而来,柯坪塔塔埃尔塔格组发现的前腹片无疑当属后一种类型。该前腹片与锅顶山汉阳鱼和巢湖宽吻鱼的前腹片相比,在形状上介于后二者之间(Lu et al., 2007)。因此,卢立伍等将其命名为Hanyangaspis sp.。此处,有两点应引起注意:其一,迄今非但在柯坪塔塔埃尔塔格组,甚至在整个塔里木红色岩系都未有Hanyangaspis或Latirostraspis的头甲发现,而在柯坪该组中出现的则是汉阳鱼科的另一属种柯坪南疆鱼,且其头甲也具雪花状纹饰(Wang et al., 2002)。其二,上述柯坪的前腹片宽为35 mm, 在尺寸上则明显小于锅顶山汉阳鱼和巢湖宽吻鱼,在后二者该片宽约为45 mm。而就头甲大小来讲,柯坪南疆鱼头甲长约55 mm (Wang et al., 2002), 同样也明显小于锅顶山汉阳鱼和巢湖宽吻鱼,在后二者其长分别约为100和110 mm (Pan, 1986a, b; Wang, 1986)。柯坪塔塔埃尔塔格组发现的前腹片和柯坪南疆鱼的头甲二者均小于锅顶山汉阳鱼和巢湖宽吻鱼的前腹片和头甲;另外,从系统分类上判断,南疆鱼与汉阳鱼、宽吻鱼同属盔甲鱼类基干类群中的同一个科,有很近的亲缘关系,其头甲腹面很可能与后两属一样共同属于同一原始类型,即口鳃窗由前腹片和腹片所覆盖。因此,将柯坪塔塔埃尔塔格组发现的前腹片归于柯坪南疆鱼,在当前是更为可取的选项。
3.2 塔里木红色岩系含鱼层位及其时代讨论
塔里木红色岩系自下而上包括柯坪塔格组、塔塔埃尔塔格组、依木干他乌组和克兹尔塔格组,其中仅有后三个组有鱼化石的发现,目前所产鱼类包括盔甲鱼类、中华棘鱼科的棘刺及软骨鱼类新疆鱼(Xinjiangichthys)的鳞片(Wang et al., 1998; Zhao and Zhu, 2014)。赵文金、朱敏(2014)在总结截至当时包括塔里木在内所有中国志留纪时期的各种鱼类资料的基础上,并参考了含鱼层中相应的无脊椎动物和植物等生物地层学的研究成果,提出了中国志留纪含鱼层的时代和对比方案,认为塔里木红色岩系中含鱼化石的塔塔埃尔塔格组与华南的溶溪组的层位相当,二者时代均为特列奇早期,而依木干他乌组则与华南的坟头组鱼群对比,其时代为特列奇中期。综合鱼群面貌来看,赵文金、朱敏提出的对比方案是可以接受的。但是有两个现象值得注意:其一是修水鱼科的成员如张氏西域鱼在塔塔埃尔塔格组的出现。在华南地区,修水鱼科分子如修水鱼、长兴鱼均出现于特列奇晚期的茅山组(Zhao and Zhu, 2014; Liu et al., 2015)。然而如同汉阳鱼科,修水鱼科作为盔甲鱼基干类群之一,其有些分子出现在特列奇早期是完全可能的。其二是巴楚假都匀鱼(Pseudoduyunaspis bachuensis)在依木干他乌组中的出现。正如王俊卿等(2002)所指出的,尽管由于保存的原因,假都匀鱼某些特征尚不清楚,从而其科级位置未定,但隶属多鳃鱼目应无问题。此前化石记录显示多鳃鱼目的成员广泛分布于中国华南和越南的安明(Yen Minh)、河江(Ha Giang), 除了产自广西博白中泥盆统艾菲尔阶“信都组”近中显眶鱼(Clarorbis apponomedianus)外(Pan and Ji, 1993), 其时代绝大多数为早泥盆世,从洛霍考夫期至埃姆斯期(Liu et al., 2015; Si et al., 2015)。盖志琨等(2018)报道了依木干他乌组之下的塔塔埃尔格组中所产的多鳃鱼新属种——锯齿宽腹鱼(Platylomaspis serratus), 代表了多鳃鱼目的最原始类型,从而将多鳃鱼类的最早出现由早泥盆世提前至志留纪兰多维列世,同时,将多鳃鱼类的古地理分布从原来的华南板块扩展到塔里木板块。因此,志留纪特列奇期出现其他多鳃鱼目成员应无问题。塔里木红色岩系各个组的时代归属问题,除了克兹尔塔格组,目前大部分得到了牙形动物生物地层学的支持(Rong et al., 2019)。克兹尔塔格组的时代由于缺少可靠的化石证据,有关该组的时代见解分歧较大,由晚泥盆世到志留纪温洛克世不一而足(Zhao et al., 2009)。20世纪90年代以来化石陆续有所发现,克兹尔塔格组可分为上、下两段,化石主要产自下段。其中下段以细砂岩、泥质粉砂岩为主,耿良玉(1993)以所产胞石Cingulochitina wronai和Eisenackituna granulate等为依据,认为其时代为罗德洛世晚期至普里道利世早期;而张师本等(1996, 2001)以所产疑源类Lophosphaeridium, Leiosphaeridium和Leiofusa等及孢子化石?Ambitisporites, Synorissporites, Apiculiretusispora, Leiotriletes, Archaeozonotriletes?和Tetrahedraletes等为依据,认为其时代为普里道利世。然而,赵文金等(2009)指出上述孢子化石中的Archaeozonotriletes?与兰多维列世晚期-艾菲尔早期的Archaeozonotriletes chulus具有相似之处,考虑到依木干他乌组和克兹尔塔格组之间连续沉积,因此并不排除本组部分地层属于兰多维列世特列奇晚期的可能。该组上段则主要为砂岩和含砾砂岩,目前所知仅产有盔甲鱼类残缺角鱼(Jiaoyu imperfectus), 之前曾被误认为是节甲类。赵文金、朱敏(2014)根据该件节甲类化石建立了以盾皮鱼亚纲节甲鱼类繁盛为特征的克兹尔塔格组合,并将其与滇东岳家山组(原关底组下部)和湘西北重新厘定的小溪组发现的节甲鱼类对比,认为克兹尔塔格组上段的时代可能为罗德洛世。戎嘉余等(2019)认为该观点尚需要进一步的野外调查求证,特别是克兹尔塔格组上段可能需要重新命名一个新的地层单位,在没有新的化石证据之前暂时将克兹尔塔格组的红层厘定为上红层,并与华南的茅山组层位相当,其时代暂定为兰多维列世特列奇期晚期。但是这一时代归属似乎又与无脊椎动物胞石、疑源类和孢粉等方面的证据相矛盾(Zhang et al., 1996, 2001)。同时,我们需要指出的是塔里木红色岩系迄今尚未发现“上红层”特有的中华盔甲鱼科分子,表明塔里木红色岩系很可能缺失志留纪兰多维列世特列奇晚期的沉积,因为该科分子广泛出现在华南地区特列奇晚期的茅山组,其成员多达5个属种:山口中华盔甲鱼(Sinogaleaspis shankouensis), 西坑“中华盔甲鱼” (‘Sinogaleaspis’ xikengensis), 浙江曙鱼(Shuyu zhejiangensis), 雷曼煤山鱼(Meishanaspis lehmani)和网状安吉鱼(Anjiaspis reticularis) (Pan and Wang, 1980; Wang, 1991; Gai and Zhu, 2005; Gai et al., 2005, 2011)。我们重新研究发现克兹尔塔格组合中的唯一的节甲类化石,实际上属于无颌类盔甲鱼类——残缺角鱼(Jiaoyu imperfectus)。在前面关于残缺角鱼的讨论中,该属种分类上的隶属有三种可能:华南鱼目中的三岔鱼科抑或目未定的昭通鱼科或者与真盔甲鱼科中具吻突的分子翼角鱼(Pterogonaspis)相近;三者现有的化石记录年代只限于早泥盆世,早自洛霍考夫期,繁盛于布拉格期,个别晚至埃姆斯期(Liu et al., 2015)。据此可将产残缺角鱼的克兹尔塔格组上段的时代暂定为早泥盆世。
早泥盆世是盔甲鱼类第二个繁盛期,其成员包括真盔甲鱼目中的真盔甲鱼科和多鳃鱼目、华南鱼目以及华南鱼目的外类群昭通鱼科,它们大量生存在早泥盆世洛霍考夫期至埃姆斯期,其中早于早泥盆世的仅有3例:其一为真盔甲鱼科的秀山盾鱼(Dunyu xiushanensis)和长孔盾鱼(D. longiforus), 出现在志留纪兰多维列-罗德洛世(Zhu et al., 2012); 另一例是秀甲鱼(Geraspis), 出现在兰多维列世特列奇期坟头组,这个属的归属尚有争议,不排除其与盾鱼有较近的关系(Pan and Chen, 1993); 第三例就是塔塔埃尔格组中所产的多鳃鱼类锯齿宽腹鱼(Platylomaspis serratus)和依木干他乌组所产的巴楚假都匀鱼(Wang et al., 1996; Gai et al., 2018)。可以肯定,角鱼与这三例中的任何成员都相距甚远。至于中晚泥盆世的盔甲鱼类,实属凤毛麟角,至今也仅有两例,而且均为残片,其一为显眶鱼(Clarorbis)可能属多鳃鱼类,出自广西博白“信都组”,可能属于中泥盆世艾菲尔期(Pan and Ji, 1993); 另一例乃是发现于宁夏中宁的上泥盆统法门阶中宁组,因过于破碎,只能认为其属盔甲鱼类(Pan et al., 1987)。显然这两例的成员也不会与角鱼有较近关系。总之, 含盔甲鱼类残缺角鱼的克兹尔塔格组的上段为早泥盆世,这与上述依据所产胞石、孢子和疑源类而将该组下段时代定为志留纪晚期沉积是并行不悖的。当然,从整个盔甲鱼亚纲系统发育的角度来看,最原始的多鳃鱼类锯齿宽腹鱼(Platylomaspis serratus)在塔塔埃尔塔格组的发现,将多鳃鱼目与华南鱼目及其外类群昭通鱼科的最早分异时间从早泥盆世提前至志留纪兰多维列世(Gai et al., 2018), 因此后续野外工作中在更古老的地层中发现残缺角鱼(Jiaoyu imperfectus)类型的盔甲鱼类是完全有可能的。
既然克兹尔塔格组下段的时代为志留纪罗德洛世晚期至普里道利世,而其下伏的依木干他乌组为志留纪兰多维列世,因此,塔里木红色岩系很可能也缺失志留纪温洛克世时期的沉积。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOIURL [本文引用: 2]
DOIURL [本文引用: 4]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 4]
[本文引用: 1]
[本文引用: 4]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 15]
[本文引用: 1]
[本文引用: 12]
[本文引用: 6]
[本文引用: 6]
[本文引用: 3]
[本文引用: 1]
[本文引用: 2]
[本文引用: 3]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 2]
DOIURL [本文引用: 2]
[本文引用: 2]
[本文引用: 3]
[本文引用: 1]
[本文引用: 5]
[本文引用: 16]
[本文引用: 2]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 4]
[本文引用: 4]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}