 ,1,2,3, 李春晓1,2,4, 张晓晓1,2,4,5, 江左其杲1,2,4, 叶捷1,2, 李录1,2,4, 李福昌6
,1,2,3, 李春晓1,2,4, 张晓晓1,2,4,5, 江左其杲1,2,4, 叶捷1,2, 李录1,2,4, 李福昌6A record of the early Protanancus and Stephanocemas from the north of the Junggar Basin, and its implication for the Chinese Shanwangian
WANG Shi-Qi,1,2,3, LI Chun-Xiao1,2,4, ZHANG Xiao-Xiao1,2,4,5, JIANGZUO Qi-Gao1,2,4, YE Jie1,2, LI Lu1,2,4, LI Fu-Chang6收稿日期:2018-06-6网络出版日期:2019-04-20
| 基金资助: |
Corresponding authors: *wangshiqi@ivpp.ac.cn
Received:2018-06-6Online:2019-04-20

摘要
中国新近纪哺乳动物年代序列的研究已经有了很大的进展,然而,山旺期早期(20~18 Ma)的哺乳动物化石记录却非常缺乏。报道了新疆准噶尔盆地北缘吃巴尔我义东地点索索泉组上部山旺期早期(18.6 Ma)的化石,包括早期欧亚大陆象类和早期鹿科动物。其中,嵌齿象的M2第二脊具有发育良好的副齿柱三叶,因此可以鉴定为原互棱齿象未定种(Protanancus sp.), 属于铲齿象科(Amebelodontidae), 形态上比山旺期晚期的短吻原互棱齿象(P. brevirostris)和托氏原互棱齿象(P. tobieni)原始。而鹿科的鹿角化石具有比较小但高的掌状部分,因此被鉴定为皇冠鹿未定种(Stephanocemas sp.), 属于柄杯鹿亚科(Lagomerycinae)。通过形态学对比认为,该鹿角在系统演化上非常接近皇冠鹿和柄杯鹿(Lagomeryx)的分化位置,并与原始的卢瓦鹿(Ligeromeryx)相去不远。以往的研究证明,长鼻类和鹿科动物在中国出现在谢家期和山旺期之交,因此,巴尔我义东地点原互棱齿象和皇冠鹿的发现首次提供了在这一地区寻找长鼻类和鹿类的重要线索,并且有可能建立山旺阶的下界层型。新材料的层位过去在中国境内很少发现化石,因此有助于深入理解中国山旺期早期的哺乳动物面貌。
关键词:
Abstract
The Chinese Neogene mammalian fauna sequence has been well established, except for a gap in the early Shanwangian (~20-18 Ma) for which there are few known records of fossil mammals in China. Hereby we report on a newly discovered fossiliferous horizon from the upper part of the Suosuoquan Formation (18.6 Ma), Chibaerwoyi East locality, northern Junggar Basin, Xinjiang Uyghur Autonomous Region. In this locality, the occurrences of an early Eurasian proboscidean and an early cervid clearly indicate an early Shanwangian Age of the Early Miocene. The proboscidean check teeth are characterized by the well-developed posttrite trefoils on the second lophs. It was identified as Protanancus sp. (Amebelodontidae), which is morphologically more primitive than the late Shanwangian P. brevirostris and P. tobieni. The cervid antler is characterized by the relatively small yet high palm of the antler and relatively short tines. It was identified as Stephanocemas sp. (Lagomerycinae), which is phylogenetically very close to the dichotomy of Stephanocemas and Lagomeryx, and is morphologically close to the more ancestral Ligeromeryx. Previous studies have shown that both proboscideans and cervids occurred at the Xiejian/Shanwangian transition period. Therefore, the new materials of Protanancus sp. and Stephanocemas sp. give important clues to the first appearance data of proboscideans and cervids in this area, which facilitates establishing the lower stratotype of the Chinese Shanwangian. In conclusion, the new findings represent a rarely known stage of mammalian biostratigraphy in China and are helpful for understanding the faunal succession during the early Chinese Shanwangian.
Keywords:
PDF (7207KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
王世骐, 李春晓, 张晓晓, 江左其杲, 叶捷, 李录, 李福昌. 准噶尔盆地北缘的早期原互棱齿象和皇冠鹿的化石记录及其对中国山旺阶的地层学意义. 古脊椎动物学报[J], 2019, 57(2): 133-154 DOI:10.19615/j.cnki.1000-3118.181108
WANG Shi-Qi, LI Chun-Xiao, ZHANG Xiao-Xiao, JIANGZUO Qi-Gao, YE Jie, LI Lu, LI Fu-Chang.
In Eurasia (except the Indian subcontinent), proboscideans (elephants and their relatives) and cervids (deer and their fossil relatives) first occurred in the middle Early Miocene (G?hlich, 1999; Gentry et al., 1999). Proboscideans appeared during the Paleogene of the Ethiopian Realm (Gheerbrant and Tassy, 2009), they subsequently first reached the Indian Subcontinent during the Late Oligocene (Antoine et al., 2003), and then northern China during the Early Miocene (Wang et al., in press). There are several early records of proboscideans in China. The Shanwang fauna yields Gomphotherium sp. (Yan et al., 1983), which was recently dated to ~17 Ma (He et al., 2011). In the Zhangjiaping Loc., Lanzhou Basin, proboscidean fragments were identified and the horizon was estimated to ~19 Ma (Qiu et al., 2001). Recently, proboscidean fragments were recovered from the Yangping, Wushan Basin, with the estimated aged between 20-21 Ma (Sun et al., 2018). Outside of China, G. annectens from Hiramaki Formation, Mino, Japan, was dated to ~19.5 Ma (Tomida et al., 2013). In Europe, there are several records of early proboscideans in MN 3-4 (Tassy, 1990).
By contrast, the Cervidae are a family endemic to the Eurasian group virtually throughout their evolutionary history. This group is characterized by their periodically casted cranial appendages (i.e. antlers) (Groves, 2007). In Europe, cervids occurred in MN 3, represented by Procervulus and Ligeromeryx, with two- and multi-pronged antlers respectively (Gentry et al., 1999). However, the earliest occurrence of cervids in China has been only vaguely appraised. Ligeromeryx (formerly attributed to Lagomeryx) and Heterocemas were recorded in the Shanwang fauna at ~17 Ma (Young, 1937; Telhard de Chardin, 1939). Zhang et al. (2011) reported Lagomeryx and Stephanocemas from the Early Miocene Damiao fauna. They correlated this fauna to the Xiejian Age, and then it was dated to 20~21 Ma (Kaakinen et al., 2015). Wang et al. (2009a) reported Ligeromeryx or Lagomeryx from the upper Aoerban fauna. A detailed study of the material and paleomagnetic dating are still underway. However, this fauna should be correlated to the Xiejian/Shanwangian transition period.
Abundant fossils have been discovered from the Cenozoic sediments from the Ulungur River drainage area, northern Junggar Basin, Xijiang Uyghur Autonomous Region, especially from the Early Miocene eolian reddish thin-clasted Suosuoquan Formation and the Middle Miocene fluviolacustrine greenish thick-clasted Halamagai Formation (Ye et al., 2001a, b, 2003, 2012; Meng et al., 2006, 2013) (Fig. 1A). Three fossil assemblages, Suosuoquan I, II, and III, have been recognized from the lower part of the Suosuoquan Formation, and the latter two were correlated to the Xiejian Age (Suosuoquan I is in Oligocene) (Meng et al., 2006, 2013). The Halamagai fauna from the Halamagai Formation was correlated to the Tunggurian Age, which is characterized by the occurrence of Platybelodon (Ye et al., 2001a, b). However, fossils were rarely discovered from the upper part of the Suosuoquan Formation, which should be correlated to the Shanwangian Stage (except for the top sandstones of the Duolebulejin section, which continuously transforms to the Halamagai Formation, see Ye et al., 2012).
Fig. 1
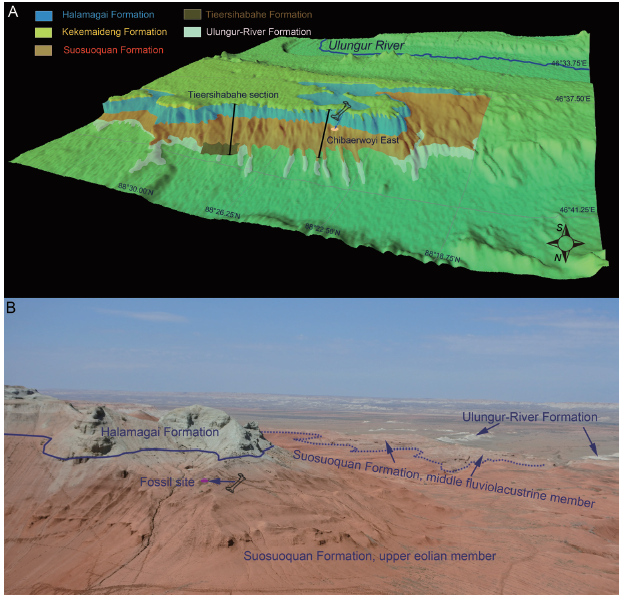 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1Geomorphologic and geographic information of the study area
A. 3D topology map of the Tieersihabahe Terrace with the Tertiary geologic units along the north cliff of the Tieersihabahe Terrace; B. the photo of the fossil site showing the subdivision of the Chibaerwoyi East section
Recently, a new fossiliferous horizon has been discovered from the upper Suosuoquan Formation at the Chibaerwoyi East locality (Figs. 1B, 2A, B) on the Tieersihabahe Terrace, Ulungur River drainage area. The screening of the samples of small mammals is still incomplete at this time. However, this fossil assemblage is characterized by several large vertebrates, such as Protanancus sp., Stephanocemas sp., and the Trionychidae gen. et sp. indet. The fossils mark the first appearance of proboscideans and cervids in this depositional series and clearly indicate the Shawangian Age of the assemblage. Therefore, the new materials are of quite considerable importance for studying the early evolution and dispersal of proboscideans and cervids in the Far East, and for further understanding of the Chinese Shanwangian.
Abbreviations IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing; MNHN, Musée National d’Histoire Naturelle, Paris; NMB, Naturhistorisches Museum, Basel; SW, Shandong Linqu Shanwang Paleontological Museum, Linqu; MN, European Neogene mammal zone; NMU, Chinese Neogene Faunal Units.
1 Materials and methods
The new materials reported are currently housed at the IVPP. Comparative materials referable to Gomphotherium cf. G. annectens are housed in SW. Information regarding comparative materials was obtained from previous publications (Borissiak and Belyaeva, 1928; Borissiak, 1929; Colbert, 1936; Bohlin, 1937; Stehlin, 1937; Young, 1937, 1964; Tassy, 1983, 1984 , 1986, 1994, 2014; Ye and Jia, 1986; Ye, 1989; Azanza and Menendez, 1989 -1990; Azanza and Ginsburg, 1997; Wang et al., 2009b; R?ssner, 2010; Markov and Vergiev, 2010; Sanders et al., 2010, Wang et al., 2013, 2015, 2016; Deng et al., 2014; Suraprasit et al., 2014; Yang et al., 2017).Fig. 2
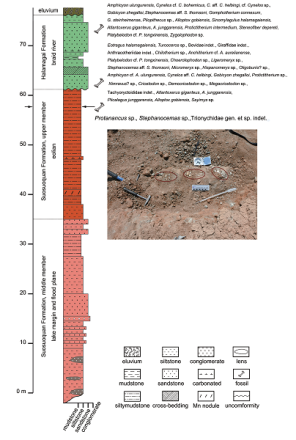 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Stratigraphic column of the Chibaerwoyi East section
Note the fossil horizon at the top of the Suosuoquan Formation (indicated by arrows) The insert panel shows the fossils exposed on the ground surface
The terminology adopted here to describe the occlusal structures of gomphotheriid cheek teeth follows Tassy (1996) (Fig. 3A), and that for the lagomerycine antler was modified from Colbert (1936) (Fig. 3D). Gomphotheriid tooth measurement protocols follow Tassy (2014) (Fig. 3B, C) and lagomerycine antler measurements follow Wang et al. (2009b) (Fig. 3E, F). All measurements were taken using digital calipers (in mm). Micrography of the lagomerycine antler sample for histological analysis was taken by a Leica MC170 HD microscope.
Fig. 3
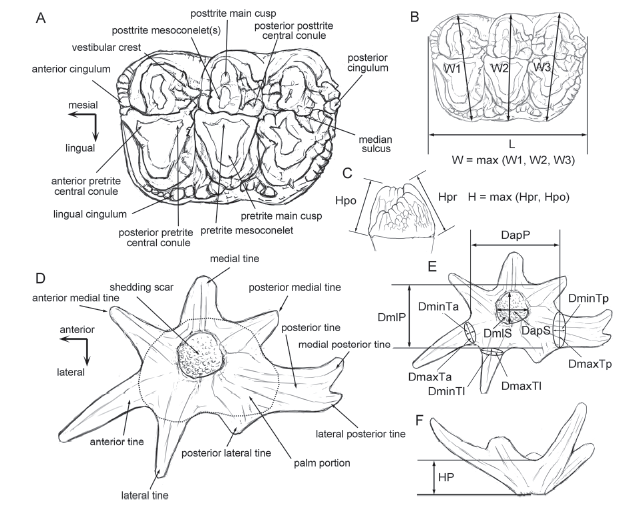 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3Terminology and measurements of gomphotheriid cheek tooth and lagomerycine antler
A. terminology of the gomphotheriid cheek tooth, the sketch is based on a left M2 of Sinomastodon praeintermedius; B, C. occlusal (B) and height (C) measurements of the gomphotheriid cheek tooth; D. terminology of the lagomerycine antler (in ventral view), the sketch is based on a right antler of Stephanocemas aff. S. thomsoni; E, F. ventral (E) and lateral (F) measurements of the lagomerycine antler Abbreviations: DapP. anteroposterior diameter of the palm portion; DapS. anteroposterior diameter of the shedding scar; DmaxTa, DmaxTl, and DmaxTp. maximal diameter of the anterior, lateral, and posterior tines; DminTa, DminTl, and DminTp. minimal diameter of the anterior, lateral, and posterior tines; DmlP. mediolateral diameter of the palm portion; DmlS. mediolateral diameter of the shedding scar; H. hight; HP. height of the palm portion; Hpo. height at the posttrite side; Hpr. height at the pretrite side; L. length; W. width; W1, 2, and 3. width of the 1st, 2nd, and 3rd loph
The measurements of the thickness of strata were undertaken using Jacob’s staff (for thick layers, > 1 m) and tapeline (for thin layers, < 1 m). Descriptions of rock coloration were based on the Munsell Color Chart (Landa and Fairchild, 2005); and determinations of granule sizes were based on the Wentworth Grain Size Chart (Krumbein and Aberdeen, 1937).
2 Geological settings
The Tieersihabahe Terrace is situated between 88°20′E and 88°40′E west-east, and between 46°35′N and 46°40′N south-north (Fig. 1A). Cenozoic deposits are well exposed along the north cliff of the Tieersihabahe Terrace. The sedimentary sequence consists bottom-up of the Eocene Ulungur-River (= Ulunguhe) Formation, the Late Oligocene Tieersihabahe Formation, the Late Oligocene and Early Miocene Suosuoquan Formation, the early Middle Miocene Halamagai Formation, and the late Middle Miocene Kekemaideng Formation (Ye et al., 2001a, b, 2012; Meng et al., 2006, 2013). Unconformities occur between most stratal units in contact except for the conforming contact between the Tieersihabahe and the overlying Suosuoquan Formations (Ye et al., 2001a, b, 2003, 2012; Meng et al., 2006, 2013) (Fig. 1A). In these units, the Suosuoquan Formation comprises of a suite of reddish eolian fine-clasts occasionally intercalated by thick, fluviolacustrine clasts. The Halamagai Formation comprises of several greenish, upwardly finning, fluvial sedimentary cycles. Abundant fossils have been discovered from these two formations, including Suosuoquan I, II, and III assemblages from the former and Halamagai fauna from the latter (Ye et al., 2001a, b, 2003).The Chibaerwoyi East section is located at the western part of the north cliff of the Tieersihabahe Terrace (Fig. 1A). In this region, the Ulungur-River and Tieersihabahe formations are poorly exposed, and the Kekemaideng Formation is entirely absent due to erosion. In the Chibaerwoyi East section, the lower part of the Suosuoquan Formation is directly overlaid by Quaternary wash-downs, and the section only consists of the upper part of the Suosuoquan Formation and the Halamagai Formation (Fig. 1B).
Chibaerwoyi East section (starting point: 46°39′48.75″N, 88°23′56.79″E; ending point 46°39′24.19″N, 88°24′7.84″E; up-bottom) (Fig. 2)
20. eluvium soil
~~~~~ unconformity ~~~~~
Middle Miocene Halamagai Formation
19. massive, dark-greenish-yellow (10Y6/6) siltstone 1.5 m
18. massive, pale-olive (10Y6/2), fine-grained, rock-fragment quartz-sandstone containing fine-grained quartz debris, poorly bedded; fossils: Amphicyon ulungurensis, Cynelos cf. C. bohemicus, C. aff. C. helbingi, cf. Cynelos sp., Gobicyon zhegalloi, Stephanocemas aff. S. thomsoni, Gomphotherium connexum, G. steinheimense, Pliopithecus sp., Alloptox gobiensis, Sinomylagulus halamagaiensis, Atlantoxerus giganteus, A. junggarensis, Protictitherium intermedium, Steneofiber depereti, Platybelodon cf. P. tongxinensis, Zygolophodon sp. 6 m
17. medium-layered, light-olive-gray (5Y5/2) and pale-brown (5YR5/2) carbonate-cemented, fine-grained, ferruginous quartz-sandstone, with horizontal beddings 0.5 m
16. massive, dark-greenish-yellow (10Y6/6) siltstone with several intercalated thin-layered, moderate-yellow (5Y7/6), moderate-yellow-green (5GY7/4), and dusky-yellow (5Y6/4) mudstones on the upper part, occasionally containing black ferromanganese 3 m
15. massive, pale-olive (10Y6/2), fine to medium grained rock-fragment quartz-sandstone containing fine-grained quartz debris, with medium-dimensional planar cross-beddings, occasionally intercalated by thin-layers or lenses of pale-greenish-yellow (10Y8/2), fine-grained conglomerates; fossils: Eotragus halamagaiensis, Turcocerus sp., Bovidae gen. indet., Giraffidae gen. indet., Anthracotheriidae gen. indet., Chilotherium sp., Anchitherium cf. A. aurelianense, Platybelodon cf. P. tongxinensis, Choerolophodon sp., Ligeromeryx sp., Stephanocemas aff. S. thomsoni, Micromeryx sp. Hispanomeryx sp., Oligobunis? sp., Amphicyon cf. A. ulungurensis, Cynelos aff. C. helbingi, Gobicyonzhegalloi, Protictitherium sp., Nimravus? sp., Cricetodon sp., Democricetodon sp., Megacricetodon sp., Tachyoryctoididae gen. indet., Atlantoxerus giganteus, A. junggarensis, Plicalagus junggarensis, Alloptox gobiensis, Sayimys sp. 1.5 m
14. massive, pale-olive (10Y6/2), medium to coarse grained quartz-sandstone containing fine grained quartz debris, porous cemented, disseminated by <Φ 1 cm grained moderate-yellow (5Y7/6) carbonated muddy angular gravel 3 m
~~~~~ unconformity ~~~~~
Early Miocene Suosuoquan Formation, upper eolian member
13. massive, light-brown (5RY6/4), silty mudstone, poorly bedded, with black ferromanganese on the broken surfaces; longitudinal cleavages present on the top, which were filled with ~Φ 1 cm grained, moderate-greenish-yellow (10Y7/4), muddy angular gravel; 6.5 m to bottom producing fossils: Protanancus sp., Stephanocemas sp., and Trionychidae gen. et sp. indet. 10 m
12. thick-layered, dark-yellowish-orange (10YR6/6), carbonated siltstones with Φ 1 cm moderate-greenish-yellow (10Y7/4) stains indicating pedogenesis 0.7 m
11. massive, light-brown, (5RY6/4) non-bedded, silty mudstone, occasionally containing fine-grained quartz debris, with black ferromanganese on the broken surfaces 6 m
10. massive light-brown (5RY6/4) non-bedded silty mudstone, occasionally with fine-grained quartz debris, containing Φ 1 cm rod-like black manganese nodules 1.5 m
9. massive, light-brown (5RY6/4), non-bedded, silty mudstone, occasionally containing fine-grained quartz debris, with black ferromanganese on the broken surfaces 1.5 m
8. massive, light-brown (5RY6/4), non-bedded, muddy siltstone, occasionally containing fine-grained quartz debris, with black ferromanganese on the broken surfaces, small particles of carbonated silty nodules spread on the ground surface 3 m Early Miocene Suosuoquan Formation, middle fluviolacustrine member
7. thick-layered, grayish-orange-pink (5YR7/2), medium to fine porous cemented arkosic sandstone, intercalated by thick-layered 5YR6/4 siltstones containing fine quartz debris 3 m
6. massive, light-brown (5RY6/4), silty mudstone containing fine-grained quartz debris, with black ferromanganese on the broken surfaces 4.5 m
5. massive, moderate-brown (5YR4/4) siltstone containing a lot of fine-grained quartz debris, with black ferromanganese on the broken surfaces 7.5 m
4. massive, moderate-orange (10R6/4), coarse-grained arkose containing fine-grained quartz debris 4.5 m
3. thick-layered, pale-yellowish-brown (10YR6/2), coarse-grained conglomeratic sandstone or fine grained sandy conglomerate, debris with maximum diameter of Φ 2 cm 1 m
2. thin- to medium-layered, moderate-brown (5YR4/4) slitstones interbedded with thin-layered, pale-yellowish-brown (10YR6/2) horizontally bedded arkosic sandstones; thickness of sandstones increase upwardly; plenty of fine-grained quartz debris throughout the layer 5.5 m
1. massive, light-brown (5RY6/4), conglomeratic, muddy siltstone intercalated by lenses of pale-yellowish-brown (10YR6/2), medium to fine grained conglomerate, debris mainly consisted of subangular and well-sorted quartz with maximum diameter of Φ 3 cm > 9 m, not to bottom
0. Quaternary washes
3 Systematic paleontology
Order Proboscidea Illiger, 1811Family Amebelodontidae Barbour, 1927
Protanancus Arambourg, 1945
Type speciesProtanancus macinnesi Arambourg, 1945.
Diagnosis (revised after Tassy, 1986) Amebelodontids with large upper tusks that are strongly ventrally curved. Mandibular tusk more dorsally concave than Archaeobelodon with concentric dentine layers. Pretrite and posttrite half-loph(id)s with slightly alternative arrangement and with a tendency of pseudo-anancoidy. Posttrite trefoils moderately developed. Cementum heavier than that in Archaeobelodon.
Distribution Early and Middle Miocene of East Africa (Protanancus macinnesi: Tassy, 1986) and East Asia (P. brevirostris and P. tobieni, respectively, see Wang et al., 2015); Middle and Late Miocene of the Indian Subcontinent (P. chinjiensis, see Tassy, 1983); probably in Eastern Europe (cf. Protanancus sp. from Bulgaria: Markov and Vergiev, 2010) and Southeast Asia (Protanancus from Thailand: Saegusa et al., 2005).
Protanancus sp.
(Fig. 4A-F)
New material IVPP V 25230 [field number, XJULG1709 (SSQ)-1], a nearly complete right M2, an incomplete left M2, the posterior-most loph of a right M3, and other dental fragments belonging to the same individual.
Locality Chibaerwoyi East [IVPP locality, XJULG1709; GPS, 46°39′26.91″N, 88°23′59.30″E, 753H], top of the Suosuoquan Formation, Early Miocene.
Description The specimens are heavily weathered, with plenty of postmortem cracks that were infilled with thin sediments throughout the specimen. The right M2 (Fig. 4A-C) is the most complete single tooth, slightly broken only in the first two posttrite half-lophs and in the anterior cingulum. The tooth has a brachyodont crown, the main cusps and conelets are inflated, and the interlophs are compressed. The pretrite central conules are relatively strong and high in lingual or buccal views, and posttrite central conules are also present (especially in the second loph). The cingulum is pronounced except along the buccal margin. The cementum is undeveloped. In the first pretrite half-loph, the anterior pretrite central conule extends anteriorly to merge with the anterior cingulum. The posterior pretrite central conule is moderately developed and the pretrite mesoconelet is unclear. The enamel on the first posttrite half-loph is considerably defective, but a weak posterior posttrite central conule is present (exiting as an enamel bulging). In the second pretrite half-loph, the anterior and posterior central conules are equally developed, and both are quite prominent. The anterior pretrite central conule displays tendency to extend to the first ectoflexid, and a subdivision of the posterior pretrite central conule is pronounced. The pretrite mesoconelet is large. In the second posttrite half-loph, the anterior and posterior central conules are well developed, especially for the posterior one, which is subdivided into three conules. The posttrite mesoconelet is moderately developed. In the third pretrite half-loph, the pretrite mesoconelet is strongly anteriorly positioned to merge with the anterior central conule; this element is irregularly subdivided and extends anterobuccally, showing a tendency of anancoid contact with the posterior posttrite central conule of the second half-loph. An additional conule that is lingual to the central conule is present on the anterior enamel wall of the third pretrite half-loph. The posterior pretrite central conule is absent. The third posttrite half-loph is structurally rather simple, with a main cusp and a mesoconelet without central conules. Three enamel cuspules rise from the posterior cingulum, and the height increases from the buccal (posttrite) to lingual (pretrite) ones. The lingual-most cuspule is close to the main cusp of the third pretrite half-loph. Measurements: L, 106.6; W1, 54.13; W2, 56.96; W3, 57.84; Hpo, 39.97; Hpr, 41.17.
The left M2 (Fig. 4D, E), although broken in the first two posttrite half-lophs, the third pretrite half-loph, and the posterior cinglum, retains diagnostic Protanancus features, such as strong pretrite central conules, and strong posttrite central conules with subdivision in the second posttrite half-loph. All these features are almost identical to those of the right M2. In the second pretrite half-loph, the posterior central conule is slightly larger than the pretrite ones, which is a feature convergent with the “Gomphotherium angustidens group” (see below).
Several dental fragments were also collected from the same quarry, but are difficult to identify, except one fragment indicating the distal-most loph of the right M3 (Fig. 4F). The crown is brachyodont. The pretrite main cusp is broken, and the mesoconelet is strongly anteriorly positioned to merge with the anterior central conule. An enamel plate that is subdivided from the mesoconelet is crowded in the median sulcus. The posttrite half-loph is composed of a main cusp and a mesoconelet, showing a tendency of subdivision. A row of strong cuspules rises along the posterior cingulum. Two additional conules are present between the loph and the cingulum.
Comparison and discussion The Chibaerwoyi East specimen lacks the critical diagnostic elements with respect to trilophodont gomphotheres, i.e. the features from mandible and lower tusks. However, the dentition alone can provide sufficient taxonomic information in this case. In the Chibaerwoyi East specimen, an important feature is the presence of posttrite central conules, forming secondary trefoils. Among longitrostine trilophodont gomphotheres, posttrite central conules more frequently exist within the Amebelodontidae than within the Gomphotheriidae and Choerolophodontidae (Tassy, 1986; Sanders et al., 2010). For example, Gomphotherium angustidens, a typical member of the Gomphotheriidae, may also have posttrite central conules (Tassy, 2014), but these conules are weak, generally showing enamel bulking along the anterior and posterior walls of the posttrite mesoconelets. Subdivision of posttrite central conules is even more rare. It should be mentioned that in the left M2 of Chibaerwoyi East specimen, the posterior pretrite central conule of the second loph is larger than the corresponding anterior one. This feature is not only present in the G. angustidens group, but also occasionally occurs in Protanancus macinnesi (see, Tassy, 1986). Posttrite central conules are also present in the Early and Middle Miocene choerolophodontids, albeit very infrequently so (unpublished observations).
In the trilophodont Amebelodontidae, the Eurasian genera include Archaeobelodon, Platybelodon, Protanancus, and Aphanobelodon. Archaeobelodon were only reported from Western Europe and Africa (Tassy, 1984, 1986; Sanders et al., 2010). In this taxon, the posterior central conules are rarely developed (Wang S Q, personal observation in the MNHN collections). In Platybelodon, only one posttrite central conule is developed on the second loph(id) (posterior central conule in upper teeth and anterior in lower teeth) (Wang et al., 2013). Both anterior and posterior posttrite central conules are present only in Protanancus and in Aphanobelodon. Aphanobelodon is only recovered from the Middle Miocene of Tongxin, with a highly specialized feature that is eponymous of the genus—loss of upper tusks, and the check teeth are fully cementodont (Wang et al., 2017). Therefore, we attributed the Chibaerwoyi East specimen to Protanancus with a strong degree of assurance.
Previously, two species of Protanancus have been reported from China, the primitive P. brevirostris and the more derived P. tobieni (Wang et al., 2015). The discrimination is mainly based on the morphology of the mandibule and lower tusks. The former was correlated to Europe MN4 and the latter MN5 (Wang et al., in press). However, the Chibaerwoyi East locality is dated no later than 18 Ma, which is earlier than any previously known occurrence of Protanancus (see below). The Chibaerwoyi East specimen is therefore probably the earliest record of the genus Protanancus. The sample is fairly small-sized with respect to what is previously known for the genus. The size is only slightly larger than the primitive Gomphotherium annectens and some G. angustidens, and is smaller than any of the other Protanancus (Fig. 5A). From Tongxin, Wang et al. (2015) reported the replacement of Protanancus tobieni by Platybelodon tongxinensis (originally identified by Wang et al. [2015] as Platybelodon danovi). This event was also recently recognized in the Shanggou section, Linxia Basin (unpublished data), and might represent the transition between the Shanwangian and Tunggurian Stages. In the Chibaerwoyi East section, Pl. tongxinensis shows its first appearance in the lower part of the Halamagai Formation (See Ye et al., 2001a, b), and Protanancus had not been identified from this region prior to the current contribution. Therefore, one may plausibly infer that most of the upper part of Shanwangian Stage is missing in the Chibaerwoyi East section (also missing in the entire Teersihabahe Terrace), due to the erosion of the overlying fluvial Halamagai Formation.
Fig. 4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 4Early Shanwangian proboscideans from the west and east part of northern China
A-F. Protanancus sp. (IVPP V 25230) from the top of the Suosuoquan Formation, Chibaerwoyi East Section, Junggar Basin; A-C. right M2, in occlusal (A), buccal (B), and lingual (C) views; D, E. left M2, in occlusal (D), and lingual (E) views; F. posterior fragment of right M3, in occlusal view; G, H. Gomphotherium cf. G. annectens (SW S700063), left M3, in occlusal (G) and buccal (H) views
Proboscidean records are rare in the Chinese early Shanwangian Stage. Among the previously disseminated records, a left M3 of Gomphotherium sp. (SW S700063) (Fig. 4G, H) from the Xiejiahe locality, the only published proboscidean fossil from the Shanwang region (Yan et al., 1983), is worth mentioning here. The first loph as well as the third and fourth posttrite lophs were broken down. The interlophs are compressed by the adjacent lophs, the crown is not high, and the subdivision of lophs is not prominent. The posterior pretrite central conule is poorly developed (a small posterior central conule is present on the second loph and absent on the third loph), the pretrite mesoconelet is triangular and is poorly separated from the anterior pretrite central conule (in the third loph), and posttrite central conules are absent (seen on the second and third lophs). These features are very similar to Gomphotherium annectens from the Hiramaki Formation, Japan (Tassy, 1994) (dating to ~19 Ma, see Tomida et al., 2013). Due to the lack of further evidence, here we refer the Shangwang material to G. cf. G. annectens. Possibly, Protanancus sp. and G. cf. G. annectens represent two types of early Shanwangian proboscideans inhabiting eastern and western China, respectively.
Order Artiodactyla Owen, 1848
Family Cervidae Goldfuss, 1820
Subfamily Lagomerycinae Pilgrim, 1941
Stephanocemas Colbert, 1936
Type speciesStephanocemas thomsoni Colbert, 1936.
Diagnosis Antlers showing a strong tendency of horizontal palmation in central part, i.e. tines short and small relative to palm portion (compared to Ligeromeryx and Heterocemas). Large antlers relative to pedicle (compared to Lagomeryx). Shedding scar generally present (compared to Ligeromeryx, Heterocemas, and Lagomeryx). Pedicle stout and medially curved (compared to Ligeromeryx, Heterocemas, and Lagomeryx). No dorsal ridges between anterior and posterior tines (compared to Paradicrocerus).
Distribution Early and Middle Miocene in East Asia (Stephanocemas thomsoni, S. chinghaiensis, S. palmatus, and S. guangheensis from China, see Colbert, 1936, Wang et al., 2009b, Deng et al., 2014), Central Asia (Stephanocemas sp., S. aralensis, and S. actauensis, from Kazakhstan, see Beliajeva, 1949, 1974, Tleuberdina et al., 1993, respectively), and Southeast Asia (S. rucha from Thailand, see Ginsburg and Ukkakimapan, 1983).
Stephanocemas sp.
(Fig. 6)
New material IVPP V 25231.1 (field number, XJULG1709 (SSQ)-2), a left casting antler with some breakage in the median part; V 25231.2 (field number, XJULG1709 (SSQ)-5), a broken tine of an antler.
Locality Chibaerwoyi East (IVPP locality, XJULG1709; GPS, 46°39′26.91″N, 88°23′59.30″E, 753H), top of the Suosuoquan Formation, Early Miocene.
Description V 25231.1, the left antler (Fig. 6A-D), broke down its medial side of the dorsal part of the palm. Therefore, two or more medial tines were missing. Because of its relatively small size (relative to V 25231.2, a single broken tine), it potentially belongs to a young individual. On the surfaces of the antler, deep grooves emitt from the palm and run along the directions of the tines. The palm is relatively high (HP in Fig. 3F), considering its dimension in horizontal plane (DapP and DmlP in Fig. 3E). In lateral view, the palm steeply shrinks, showing a bowl-like profile. In dorsal view, the palm portion is rather small relative to most of the other Stephanocemas specimens, but larger than that of Lagomeryx. Three tines are retained. The anterior tine is the longest and most slender one. It extends anterolaterally, with a nearly circular cross-section. The posterior tine is the shortest and thickest one. It is posterolaterally oriented with a rounded-triangular cross-section. The depression of the tip may indicate a potential bifurcation of this tine. The length and thickness of the medial tine is intermediated between the anterior and posterior tines. All these tines are only slightly upwardly extended.
In ventral view, the shedding scar is very clear. It is subcircular with an approximately anteroposteriorly oriented long-axis. The scar surface is concave, and is surrounded by a sharp-edged loop. On the scar surface (Fig. 6H), the bone tissue is spongy with plenty of small alveoli, indicating a strong absorption of the bone before it was casted. Conversely, the bone tissue from the internal (Fig. 6G) and surface (Fig. 6I) of the palm portion is compact, with regular arrangement that orients to the growth directions. Measurements: DapP, 35.29; HP, 14.62; DapS, 12.62; DmlS, 10.64; DmaxTa, 10.42; DminTa, 9.37; DmaxTl, 12.77; DminTl, 9.93; DmaxTp, 15.56; DminTp, 11.05.V 25231.2 (Fig. 6E, F) is a broken tine of an antler. It is slender and tapers distally with a pointed tip. Tines with similar morphology can be observed in the antler sample of Stephanocemas from the Halamagai Formation (i.e. V 8607).
Comparison and discussion The antlers of Lagomerycinae are not always easy to discriminate among various groups, especially among Ligeromeryx, Lagomeryx, and Stephanocemas. Taxonomic discrimination of the aforementioned genera becomes more difficult when considering ontogenetic variations. The genus name Lagomeryx should be confined to small lagomerycines with a long pedicle supporting a small antler (Azanza and Ginsburg, 1997), e.g. the European species Lagomeryx ruetimeyeri, La. parvulus, and La. pumilio (R?ssner, 2010). This antler palmation also occurs in Lagomeryx, in which the palm portion is relatively high with relatively large HP/DapP value (Fig. 5B). In this feature, the Chibaerwoyi East specimen is similar to Lagomeryx. However, the Chibaerwoyi East specimen is much larger than any Lagomeryx (Fig. 5B). The tines in the Chibaerwoyi East specimen are elongated, show mainly peripheral emission, and are not upwardly pointed. However, in Lagomeryx, the tines are tiny with subequal dimension and are upwardly pointed (R?ssner, 2010; Azanza and Ginsburg, 1997). The other two Asian species, “La.” complicidens from Lantian, China (Young, 1964), and “La.” mania from Mae Moh, Tailand (Young, 1964), have generally Lagomeryx-like palmete antlers. However, the palm portion is relatively large with numerous dorsally erected tines compared with their European relatives (Suraprasit et al., 2014). These features are also distinct from the Chibaerwoyi East specimen.
In Ligeromeryx, the palm portion is poorly developed. The tines are rather long, and are multi-branched within a small area. Beside the European species Ligeromeryx praestans, the Asian species “La.” triacuminatus from Tunggur, “La.” colberti, “La.”telhardi, and “Stephanocemas cf. S. thomsoni” from Shanwang, as well as “La.” tsaidamensis all seem attributable to Ligeromeryx, or at least to very closely allied genera pending further revision (Azanza and Ginsburg, 1997). The Chibaerwoyi East specimen possesses a relatively large palm portion and short tines, which is distinct from Ligeromeryx praestans and its Asian relatives. Another taxon from Shanwang is Heterocemas simpsoni (Young, 1937). The antler possesses two forks. The major one is placed proximately and the minor one is positioned at the distal part of the primary posterior branch. This morphology is substantially distinct from that of the Chibaerwoyi East specimen.
Stephanocemas can be principally summarized as an Asian genus with a large horizontal palm portion and horizontally emitted tines on its antlers. Till now, at least seven species have been established (Wang et al., 2009b; Deng et al., 2014). The European species Stephanocemas elegantulus were excluded from this genus due to the presence of dorsal ridges along the anterior and posterior tines and the upwardly pointed tines (Stehlin, 1937). It was first transferred to Stehlinoceros and then grouped with Paradicrocerus (Azanza and Menendez, 1989-1990; Wang et al., 2009b). Compared with the typical Stephanocemas species, e.g., S. thomsoni and S. palmatus, the palmation in the Chibaerwoyi East specimen is weak. The palm portion in the Chibaerwoyi East specimen is small (relative to the scar area), and the palm height is relatively large, showing a bowl-like shape in lateral view. These features are distinct with S. thomsoni and S. palmatus (relatively small HP/DapP value, see Fig. 5B). The morphology of the Chibaerwoyi East specimen might be close to primitive species of the genus such as S. chinghaiensis and S. aralensis with the close value of Hp/DapP (Fig. 5B); however, considerations of ontogenetic variation further obscures the morphological comparison at the species level. Nevertheless, the morphology of the Chibaerwoyi East specimen shows some similarity to Lagomeryx (see above). Azanza and Ginsburg (1997) argued that Lagomeryx and Stephanocemas constitute a monophyletic clade due to the acquired palmation of the antler, which branched after Ligeromeryx. Considering the early age of the locality (see below), the Chibaerwoyi East specimen is close to the dichotomy of the Lagomeryx-Stephanocemas clade, and not far away from Ligeromeryx. Due to the relatively developed palm portion and relatively strong, not upwardly pointed tines, Stephanocemas is a more preferable generic attribution than Lagomeryx and Ligeromeryx for the Chibaerwoyi East specimen.
In addition, Ye (1989) reported lagomerycine antlers from the same region. However, all reported specimens are from the overlying Middle Miocene Halamagai Formation. Ye (1989) attributed them respectively into Stephanocemas and Lagomeryx. Based on our preliminary observation, Ye’s (1989) interpretation of Stephanocemas includes two genera, Stephanocemas (here we refer to S. aff. S. thosoni, the same as Ye, 1989) and Paradicrocerus (with ridges through the anterior and posterior tines); whereas, the specimens of Ye’s (1989) Lagomeryx, as that in other Asian localities, can be aligned with Ligeromeryx (also see Fig. 5B). Therefore, the Halamagai Formation records a great diversity of the Lagomerycines, which should be traced back to the underlying Suosuoquan Formation. The Stephanocemas from the Chibaerwoyi East offers important evidence regarding evolution within the Cervidae.
Fig. 5
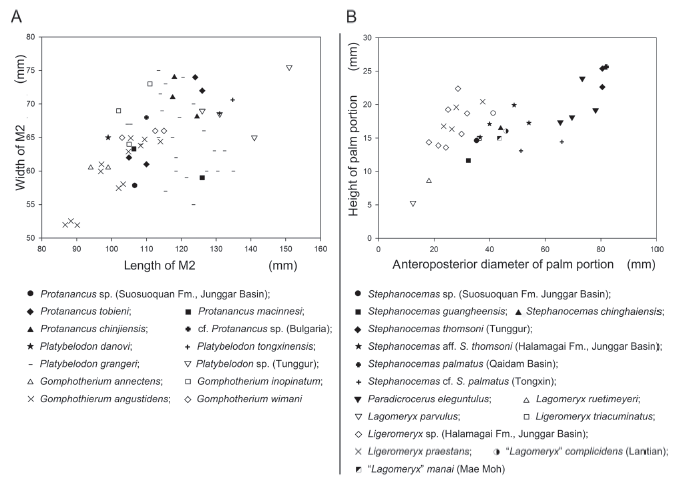 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 5Bivariate plots of measurements for some gomphotheriid M2 (A) and some lagomerycine antlers (B)
Data source: Protanancus tobieni, after Wang et al., 2015; P. macinnesi, after Tassy, 1986; P. chinjiensis, after Tassy, 1983; cf. Protanancus (Bulgaria), after Markov and Vergiev, 2010; Platybelodon danovi, after Borissiak, 1929; Pl. tongxinensis, after Ye and Jia, 1986; Pl. grangeri and Platybelodon sp. (Tunggur), after Wang et al., 2013; Gomphotherium annectens, after Tassy, 1994; G. inopinatum, after Borissiak and Belyaeva, 1928; G. angustidens, after Tassy, 2014; G. wimani, after Yang et al., 2017; Stephanocemas guangheensis, after Deng et al., 2014; S. chinghaiensis, after Bohlin, 1937; S. thomsoni (Tunggur), after Colbert, 1936; S. aff. S. thomsoni (Halamagai Formation, Junggar Basin), after Ye, 1989; S. palmatus (Qaidam Basin), after Wang et al., 2009; S. cf. S. palmatus (Tongxin), after Wang et al., 2016; Paradicrocerus eleguntulus, after Stehlin, 1937; Lagomeryx ruetimeyeri, a cast of the lectotype in NMB; La. parvulus, after R?ssner, 2010; Ligeromeryx triacuminatus, after Colbert, 1936; Li. praestans, after Stehlin, 1937; Azanza and Ginsburg, 1997; Ligeromeryx sp. (Halamagai Formation, Junggar Basin), after Ye, 1989; “La.” complicidens (Lantian), after Young, 1964; “La.” manai (Mae Moh), after Suraprasit et al., 2014
4 Significance for Chinese Shanwangian
Besides the evolutionary significance in proboscideans and cervids inferred by the new specimens from the Chibaerwoyi East locality, they also bear important stratigraphic significance for the Shanwangian Stage, both for the Ulungur River drainage area and for East Asia. Cenozoic sediments in the Ulungur River drainage area are nearly continuously deposited from the Upper Oligocene to the Upper Miocene, producing successive faunas, especially in the Upper Oligocene Tabenbulukian, lowest Miocene Xiejian, and Middle Miocene Tunggurian stages (Ye et al., 2001a, b, 2003, 2012; Meng et al., 2006, 2013, Wu et al., 2018; Jiangzuo et al., in press). However, fossils from the late Early Miocene Shanwangian were relatively sparse. Two previous Shanwangian localities from the Suosuoquan Formation, Nanganqu and Heishantou, lack continuous outcrops for complete stratigraphic correlation, and also lack large mammal fossils. Another locality, Duolebulejin, produces abundant fossil remains of small and large mammals. This fauna occurs in the top sandstones of the Suosuoquan Formation and represent the latest Shanwangian Age (Ye et al., 2012). Although lacking the key taxon, Platybelodon, the faunal assemblage appears similar to the Tunggurian Halamagai fauna that occurred only slightly later. However, fossils from the early Shanwangian Age are very rare, especially for large mammals.Fig. 6
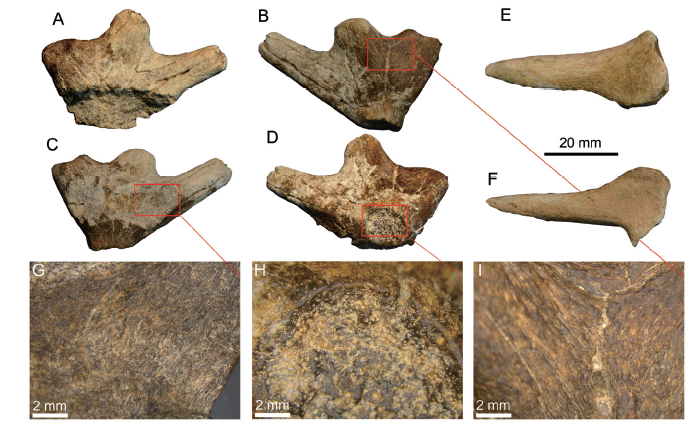 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 6Antlers of Stephanocemas sp. from the top of the Suosuoquan Formation, Chibaerwoyi East Section, Junggar Basin
A-D. left antler (IVPP V 25231.1), in dorsal (A), lateral (B), medial (C), and ventral (D) views; E, F. broken tine (V 25231.2), in dorsal (E) and ventral views (F); G-I. histology of the left antler (V 25231.1), showing the bone tissue of internal (G), the shedding scar (H), and the outer surface (I) of the palm The micrographs were taken by a Leica MC170 HD microscope, with the positions indicated by the red frame
The Suosuoquan Formation represents the earliest eolian records controlled by planetary winds system (i.e. westerly winds in the mid-latitude zone) (Sun et al., 2010). Generally, it is consisted of massive, reddish, sandy mudstones with poor development of beddings. The lithology of the Suosuoquan Formation is very stable over a large area. On the north of the Tieersihabahe Terrace, the Suosuoquan Formation comprises three members. The middle fluviolacustrine member that consists of thick-layered siltstones and mudstones intercalated by thin- to medium-layered, fine, muddy conglomeratic sandstones is sandwiched between the upper and lower eolian pourly-bedded, silty mudstones with rather stable thickness (~130 m totally). Meng et al. (2006) reported the palaeomagnetic data from the Tieersihabahe section, which is ~6 km east to the Chibaerwoyi East section (Fig. 7). The Tieersihabahe paleomagnetic column covers the Tieersihabahe, Suosuoquan, Halamagai, and Kekemaideng formations from the bottom up, spanning ~24.5-15 Ma (Meng et al., 2006). The column is almost temporarily continuous except a hiatus of ~2 Ma between the Suosuoquan Formation and the overlying Halamagai Formation. Three small mammal assemblages have been recognized from the lower part of the Suosuoquan Formation, the Tabenbulukian Suosuoquan I, the Xiejian Suosuoquan II and III biozones (Meng et al., 2006). All these fossil assemblages are from the lower 110 m of the Tieersihabahe section, and in the upper part of the Suosuoquan Formation (110-165 m) that corresponds the lower Shanwangian, no fossils were recorded (Meng et al., 2006).
Fig. 7
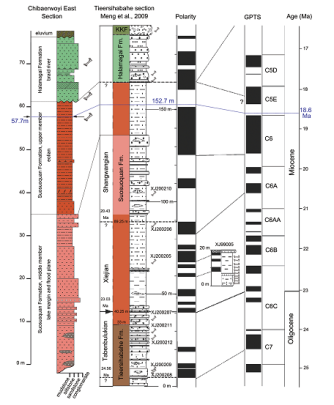 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 7Stratigraphic correlation of the Chibaerwoyi East section and the Tieersihabahe section with the dating of the fossil horizon of the upper Suosuoquan Formation The lithology, palaeomagnetic polarity, and its correlation to GPTS are redrawn after Meng et al. (2006)
Our Chibaerwoyi East section covers most of the Suosuoquan middle fluviolacustrine member, the entirety of the Suosuoquan upper eolian member, and the Halamagai Formation (Figs. 1B, 2). The Chibaerwoyi East fossil locality is located at 19.2 m to the eolian/fluviolacustrine boundary (Figs. 2, 7). The Suosuoquan upper eolian member is 22.7 m, very close to the thickness of the same strata (25.7 m) in the Tieersihabahe section. Assuming a uniform sedimentation rate in eolian deposits and considering the potentially inconsistent erosion of the overlying fluvial Halamagai Formation, we dated the Chibaerwoyi East fossil locality to 18.6 Ma, which is approximately equal to 19.2 m of the Suosuoquan upper eolian member (152.7 m in the whole section) in the Tieersihabahe palaeomagnetic column (Fig. 7). This age corresponds to the early Shanwangian, and is later than the Shanwang fauna (~17 Ma).
In China, fossils from the early Shanwangian Stage (i.e. ~20-18 Ma) are especially rare. In the recent Chinese Neogene mammalian biochronology chart (Deng, 2006:fig. 2), almost no mammalian fauna was found in this period. In the sequence of Chinese Neogene Faunal Units (NMUs), NMU 4 covers ~20-17 Ma, correlated to the European MN 3 and 4. However, all known NMU 4 faunas were correlated to MN 4, and no fauna can be correlated to MN 3 prior to Deng (2006) . Recently, several new findings filled this gap and uncovered the faunal appearance in the early Chinese Shanwangian, such as the upper Aoerban fauna from Nei Mongol (Wang et al., 2009a) and the Yangping fauna, Gansu (Sun et al., 2018).
Our Chibaerwoyi East fauna also contributes to our knowledge of the early Shanwangian, especially due to the co-occurrence of proboscideans and cervids. Because both groups may have occurred at the Xiejian/Shanwangian transition period (see Introduction), the first occurrence of either the earliest proboscidean or the earliest cervids is potentially the biochronological marker for the lower stratotype boundary of the Shanwangian, which has not been previously established. The Chibaerwoyi East fauna seems slightly young for establishing the stratotype of the Shanwangian lower boundary, not only because of the dating of the fossil locality (18.6 Ma), but also the morphology of the Stephanocemas sp.—the palmation of the antler, and the regulation of the shedding scar are derived features compared with the earliest Ligeromeryx. Therefore, the occurrence of the Cervidae should be earlier than the Chibaerwoyi East. Nevertheless, it is worth searching for the first occurrence of proboscideans and/or deer in the Suosuoquan Formation in the Ulungur River drainage area, which is fundamental for the early Shanwangian of China.
Acknowledgements
We are thankful to Deng Tao, Wang Xiao-Ming, Ni Xi-Jun, Bastien Mennecart, Manuela Aiglstorfer, and Lo?c Costeur for their useful discussions and help with the literature. We thank Zhang Han-Wen and Wang Li-Hua for providing much advice upon reviewing the manuscript. We thank Xu Si-Jian, Suo Chao, and Su Dan for fossil excavation and preparation. This work is supported by the Chinese Academy of Sciences (Grant Nos. XDA20070203, QYZDY-SSW-DQC022), the National Natural Science Foundation of China (Grant Nos. 41372001, 41430102, 41625005), the Special Research Program of Basic Science, the Technology of the Ministry of Science and Technology (Grant No. 2015FY310100-14), and all China Commission of Stratigraphy Project (Grant No. DD20160120-04).参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 6]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 4]
[本文引用: 5]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 10]
[本文引用: 5]
[本文引用: 1]
[本文引用: 4]
[本文引用: 1]
[本文引用: 3]
[本文引用: 4]
[本文引用: 2]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 3]
[本文引用: 7]
[本文引用: 3]
[本文引用: 1]
[本文引用: 4]
[本文引用: 1]
[本文引用: 2]
[本文引用: 3]
[本文引用: 6]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 5]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 7]
[本文引用: 2]
[本文引用: 7]
[本文引用: 3]
[本文引用: 1]
[本文引用: 5]
[本文引用: 3]
[本文引用: 4]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}