 ,1,2, 邹晶梅1,2, 周忠和1,2
,1,2, 邹晶梅1,2, 周忠和1,2A taxonomical revision of the Confuciusornithiformes (Aves: Pygostylia)
WANG Min,1,2, Jingmai O’CONNOR1,2, ZHOU Zhong-He1,2收稿日期:2018-02-5网络出版日期:2019-01-20
| 基金资助: |
Corresponding authors: wangmin@ivpp.ac.cn
Received:2018-02-5Online:2019-01-20

摘要
孔子鸟目是早白垩世鸟类中具有角质喙和尾综骨的最原始的鸟类类群之一。在过去近20年的研究历史中,千余件孔子鸟类标本被发现,其数目多过其他中生代鸟类和非鸟类恐龙。目前,共计4个属,10个种被先后归入到孔子鸟目中,然而这些属种的分类位置是否有效尚不清楚。通过对已发表的所有孔子鸟类属种进行详细的对比研究,证实了其中多数属种的鉴定特征有误,这些特征或受保存的影响,或代表了种内和个体发育的变化,亦或广泛分布在孔子鸟类或基干鸟类中,而并不能成为这些属种建属或建种的依据。认为此前命名的孙氏孔子鸟、费氏孔子鸟、义县锦州鸟、张吉营锦州鸟和建昌孔子鸟均属于圣贤孔子鸟的同物异名;而川州孔子鸟未保存确切的鉴定特征而难以和其他孔子鸟类相区别,暂将其归入孔子鸟科未定种。至此,对所有已报道的孔子鸟目的分类厘定认为,目前孔子鸟目共包括1科、3属、4种,分别是圣贤孔子鸟、杜氏孔子鸟、横道子长城鸟和郑氏始孔子鸟,并对这些属种各自的鉴定特征进行了修正。
关键词:
Abstract
The Confuciusornithiformes is a basal clade of Early Cretaceous birds that includes the oldest and most basal birds with a toothless beak and an abbreviated bony tail. Over the last two decades, thousands of specimens have been collected, more than for any other group of Mesozoic birds or non-avian dinosaurs. Ten species separated into four genera have been erected with limited taxonomic phylogenetic scrutiny. Here, we perform a comparative study of these ten species, and demonstrate that most of these taxa were originally diagnosed by characters that prove to be either preservational artifacts, intraspecific variations, subject to ontogenetic variation, or widely distributed among the Confuciusornithiformes or a more phylogenetically inclusive group. Our results suggest that ‘Confuciusornis suniae’, ‘C. feducciai’, ‘Jinzhouornis yixianensis’, ‘J. zhangjiyingia’, and ‘C. jianchangensis’ are all junior synonyms of C. sanctus. ‘C. chuonzhous’ lacks autapomorphies of C. sanctus and is referred to Confuciusornithiformes incertae sedis. Our taxonomic reappraisal of published materials indicates that the Confuciusornithiformes consists of one family, three genera, and four species: C. sanctus, C. dui, Changchengornis hengdaoziensis, and Eoconfuciusornis zhengi, for which we provide revised diagnoses.
Keywords:
PDF (10304KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
王敏, 邹晶梅, 周忠和. 孔子鸟目(鸟类:尾综骨类)的分类厘定. 古脊椎动物学报[J], 2019, 57(1): 1-37 DOI:10.19615/j.cnki.1000-3118.180530
WANG Min, Jingmai O’CONNOR, ZHOU Zhong-He.
1 Introduction
The Confuciusornithiformes arguably is the most specimen-rich clade of Mesozoic birds, with thousands of known specimens curated in public and private collections worldwide (Chiappe et al., 1999; Zhou and Zhang, 2006a; Chiappe and Meng, 2016). Most of these specimens are preserved as complete and articulated skeletons, often with additional soft tissue traces (e.g., feathers, skins, ovarian follicles, beta-keratin, etc.) (Zhou and Zhang, 2006a; Wang and Zhou, 2017a). Together these wealthy specimens provide unprecedented information regarding the skeletal morphology, ontogeny, and biology of these Early Cretaceous birds (Zhang et al., 2010; Pan et al., 2016; Zheng et al., 2017) and significantly advances our knowledge of early avian evolution. Confuciusornithiforms have an iconic role in tracing the evolution of key avian features such as the horny beak and pygostyle (O'Connor et al., 2011; Wang and Zhou, 2017a). Known specimens of confuciusornithiforms come mainly from the Early Cretaceous Jehol Biota in northeastern China (Zhou and Zhang, 2006a; Li, 2010), with a few specimens from contemporary deposits in North Korea (Gao et al., 2009). Confuciusornithiforms are present throughout the entirety of the Jehol, indicating that this clade persisted for at least 11 million years (130.7 Ma-120 Ma; Zhou, 2014). Three specimens of the Confuciusornithiformes have been collected from the Protopteryx-horizon of the Huajiying Formation (Zhang et al., 2008; Zheng et al., 2017; Navalón et al., 2018), the oldest fossil bearing unit of the Jehol Biota and the second-oldest known avian-bearing deposits in the world (Zhou, 2014), being surpassed in age only by the German Upper Jurassic Solnhofen Limestones that yield Archaeopteryx. Confuciusornithiforms are extremely abundant in the 125 Ma Yixian Formation, with only a few specimens collected from the younger Jiufotang Formation. Within a period of less than 15 years, four genera and ten species were assigned to the Confuciusornithiformes (Confuciusornis sanctus, 'C. suniae', C. dui, 'C. chuonzhous', 'C. feducciai', 'C. jianchangensis', Changchengornis hengdaoziensis, 'Jinzhouornis yixianensis', 'J. zhangjiyingia', and Eoconfuciusornis zhengi) (Hou et al., 1995, 2002; Hou, 1997; Chiappe et al., 1999; Zhang et al., 2008, 2009; Li et al., 2010). More than half of these taxa have only ever been described in Chinese (Hou, 1997; Hou et al., 2002; Li et al., 2010), which has hindered evaluation of their validity through direct study or phylogenetic analysis by the international research community. In part due to the rapid rate of discovery, morphologically similar new species were not adequately compared to other available materials and thus poorly diagnosed. As a result, nearly every species has had its validity questioned at least once in the literature. Previous studies regarded 'J. zhangjiyingia', 'J. yixianensis', 'C. chuonzhous', 'C. suniae', and 'C. feducciai' as junior synonyms of C. sanctus (Chiappe et al. 1999, 2008; Marugán-Lobón et al., 2011). However, this conclusion was neither reached through detailed comparisons between these putative taxa nor documented with photographs or other figures. In addition to Confuciusornis, two genera have been widely accepted: while Changchengornis can be readily distinguished by its hooked rostrum (Chiappe et al., 1999), it is notable that no specimens referable to this genus have surfaced; the basal Eoconfuciusornis is distinguishable by provenance and a few primitive features (Zhang et al., 2008), but with the discovery of new specimens (Zheng et al., 2017; Navalón et al., 2018) the taxonomy of this taxon is no longer straight forward. Currently, the Confuciusornithiformes includes several C. sanctus-like species of uncertain validity. Although no other Mesozoic avian group is represented by numbers of specimens comparable to the Confuciusornithiformes, most specimens have not been properly identified at the species or even generic level. The current state of confuciusornithiform taxonomy limits our ability to make taxonomic referrals and severely undermines our understanding of the diversity and degree of morphological disparity in this important clade. In order to rectify these long-standing taxonomic issues, we performed a comparative study of all published taxa referred to the Confuciusornithiformes. Here we provide a taxonomic revision of this clade, in which we address the validity of every character initially used to diagnose each taxon and offer revised diagnoses where appropriate.2 Materials and methods
Anatomical abbreviations ad-1,2, proximal and distal phalanges of alular digit; al, alular metacarpal; an, angular; ar, articular; cf, caudal mandibular fenestra; cv, cervical vertebrate; de, dentary; dm, dorsal process of maxilla; dI-IV, pedal digits 1-4; en, external naris; fd, fenestra of deltopectoral crest; fe, fenestra; fi, fibula; fm, femur; fp, frontal process of premaxilla; fr, frontal; fu, furcula; hu, humerus; if, infratemporal fenestra; is, ischium; ju, jugal; lc, lacrimal; lp, lateral process of sternum; lti, left tibiotarsus; ma, major metacarpal; mad-1-3, proximal, intermediate and distal phalanges of major digit; mb, mandible; md-1-4, proximal, second, penultimate, and distal phalanges of minor digit; mi, minor metacarpal; mtI-V, metatarsals I-V; mx, maxilla; na, nasal; nm, notch of mandibular symphysis; or, orbital; pa, parietal; pm, premaxilla; po, postorbital; pu, pubis; qj, quadratojugal; qu, quadrate; ra, radius; rc, right coracoid; rf, rostral mandible fenestra; sc, scapulocoraocid; scl, scleral ossicle; sp, splenial; sq, squamosal; st, sternum; su, surangular; ti, tibiotarsus; tm, tarsometatarsus; tu, tubercle; tv, thoracic vertebrate; ul, ulna; vp, ventral process of surangular; xp, xiphoid process (l, left; r, right).Institutional abbreviations BMNHC, Beijing Museum of Natural History, Beijing, China; DNHM, Dalian Natural History Museum, Dalian, China; GMV, National Geological Museum of China, Beijing, China; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China; MACN-PV, Vertebrate Paleontology Collection, Museo Argentino de Ciencias Naturales 'Bernardino Rivadavia', Buenos Aires, Argentina; PMOL, Liaoning Paleontological Museum, Shenyang, China.
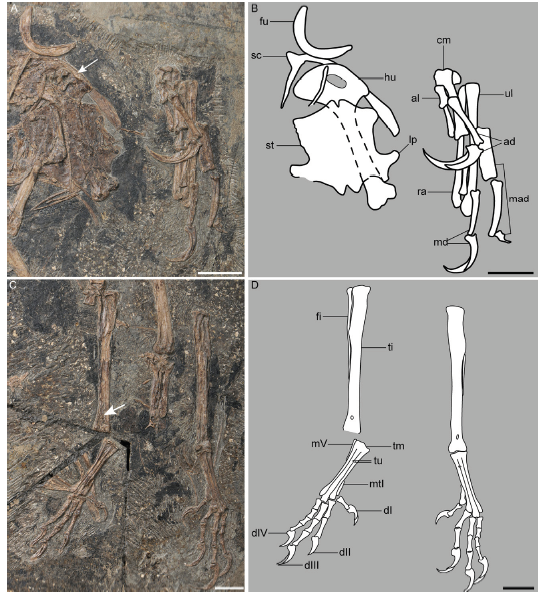
Specimens included We reexamined all the published material representing the Confuciusornithiformes at first hand, including holotypes and referred specimens of Confuciusornis sanctus Hou et al., 1995 (Figs. 1, 2), 'C. suniae' Hou, 1997, 'C. chuonzhous' Hou, 1997 (Fig. 3), Changchengornis hengdaoziensis Chiappe et al., 1999 (Figs. 4-6), 'Jinzhouornis yixianensis' Hou et al., 2002 (Fig. 7), 'J. zhangjiyingia' Hou et al., 2002 (Figs. 8, 9), Eoconfuciusornis zhengi Zhang et al., 2008 (Figs. 10, 11), 'C. feducciai' Zhang et al., 2009 (Figs. 12, 13), 'C. jianchangensis' Li et al., 2010 (Fig. 14). The holotype and the only known specimen of C. dui Hou et al., 1999, catalogued as IVPP V 11553, is not available for study, and thus we relied on published photographs for comparison.
Fig. 1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1Skull of Confuciusornis sanctus in comparison with those of C. dui and other Early Cretaceous birds
A, C. photograph (A) and line drawing (C) of Confuciusornis sanctus IVPP V 13168 with a separate drawing of the maxilla (right), showing the halberd-shaped dorsal process of the maxilla (considered an autapomorphy here); B. C. sanctus V 13171 with line drawing of the maxilla; D. drawing of C. dui with a separate drawing of the maxilla (modified from Hou et al., 1999); E. Jeholornis sp. V 14978 (Aves, non-Pygostylia); F. Pterygornis dapingfangensis V 16363 (Aves, Enantiornithes). The arrowheads in B denote the rostral notches of the premaxillary and mandibular symphyses. The arrow in B indicates the ventral process of the surangular (considered an autapomorphy here). Scale bars = 10 mm; C, D not to scale
Fig. 2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Detail anatomy of Confuciusornis sanctus showing some diagnostic characters
A. right forelimb of C. sanctus IVPP V 10928 (hand in ventral view, humerus in cranial view); B. hindlimbs of C. sanctus V 11552 in dorsal view; C, D. photograph (C) and reconstruction (D) of the sternum of C. sanctus in dorsal view; E. sternal reconstruction of C. dui (modified from Hou et al., 1999); F. sternal reconstruction of Eoconfuciusornis zhengi (modified from Zhang et al., 2008) The arrowhead in A denotes the ridge on the craniaodorsal margin of the deltopectoral crest, and the arrow indicates the unfused carpometacarpus. The arrowheads in B mark the extensor retinaculum tubercle on the dorsal surface of the distal tibiotarsus (diagnostic character); and the arrows indicate the dorsal tubercles on metatarsals II and III. The arrows in D denote the cranial and caudal projections of the lateral process of the sternum in C. sanctus (diagnostic character). The dash lines in E indicate the length (vertical) and width (horizontal) of the sternum that are measured in this study Scale bars equal 10 mm in A and C, 5 mm in B; D-F not to scale
3 Systematic paleontology
Aves Linnaeus, 1758Pygostylia Chiappe, 2002
Confuciusornithiformes Hou et al., 1995
Diagnosis As for the family.
Confuciusornithidae Hou et al., 1995
Type genusConfuciusornis Hou et al., 1995.
Type speciesConfuciusornis sanctusHou et al., 1995.
Diagnosis Upper and lower jaws edentulous and beaked; mandible with large rostral and small caudal fenestrae that are visible laterally; boomerang-shaped furcula without a hypocleidium; coracoid and scapula fused into a scapulocoracoid; humeral deltopectoral crest triangular with sharp caudodorsal corner; claw of major digit significantly reduced; and claw of hallux smaller than those of other pedal digits.
Phylogenetic definition The most-inclusive clade that contains Confuciusornis sanctusHou et al., 1995 and Eoconfuciusornis zhengi Zhang et al., 2008, but not Sapeornis chaoyangensis Zhou & Zhang, 2002, or Passer domesticus Linnaeus, 1758.
Included taxaC. dui Hou et al., 1999, Ch. hengdaoziensis Chiappe et al., 1999, E. zhengi Zhang et al., 2008.
Confuciusornis sanctusHou et al., 1995
Junior synonyms 'C. suniae' Hou, 1997; 'J. yixianensis' Hou et al., 2002; 'J. zhangjiyingia' Hou et al., 2002; 'C. feducciai' Zhang et al., 2009; 'C. jianchangensis' Li et al., 2010.
Holotype IVPP V 10918.
Referred specimens IVPP V 11308 (former holotype of 'C. suniae'); V 14412 (former holotype of 'J. yixianensis'); V 12352 (former holotype of 'J. zhangjiyingia'); DNHM D2454 (former holotype of 'C. feducciai'); PMOL-AB00114 (former holotype of 'C. jianchangensis'); IVPP V 11307, V 11370, V 11374, V 11375, V 12644, V 13156, V 13167, V 13168, V 13171, V 13172, V 13175, V 13178, V 14373, V 14385, V 16066; GMV 2130-2133.
Horizon and locality Villages of Shangyuan, Sihetun, and Zhangjiying near Beipiao City, and Wutun Village near Jinzhou City, Liaoning Province, northeastern China (Yixian Formation, 125 Ma; He et al., 2004); Toudaoyingzi Village, Jianchang County, Huludao City, Liaoning Province, northeastern China (Jiufotang Formation, 120 Ma; He et al., 2004).
Original diagnosis Following Hou et al. (1995), a basal bird, subequal to the size of the Eichs?tt specimen of Archaeopteryx, differs from other Mesozoic birds in the following features: upper and lower jaws toothless with a horny beak; large orbit; small antorbital fenestra; humerus expanded proximally with a deltopectoral crest perforated by a pneumatic foramen; carpals unfused with metacarpals; manual digits unreduced; alular digit ungual robust and large; ischium robust with a dorsal process proximally and a slightly expanded distal end; metatarsal V present; and pedal claws large and recurved.
Revised diagnosis A confuciusornithiform distinguished by the following aut-apomorphies: maxilla with an imperforate halberd-shaped dorsal process; and tibiotarsus with a tubercle on the distal part of the dorsal surface, possibly representing an attachment point for the extensor retinaculum. C. sanctus can be further differentiated from other confuciusornithiforms on basis of the following unique combination of features: rostral part of ventral margin of dentary convex (absent in C. dui); surangular with a triangular ventral process (absent in C. dui); deltopectoral crest perforated by a suboval fenestra (considered absent in E. zhengi); proximal phalanx of major digit shorter than intermediate phalanx (opposite condition in Ch. hengdaoziensis); and hallux less than half the length of pedal digit II (proportionally shorter than in Ch. hengdaoziensis).
Remarks This taxon was originally described from eight specimens collected from Yixian Formation deposits near Shangyuan Village in Liaoning: IVPP V 10895, V 10918-V 10925 (Hou et al., 1995). Since then numerous specimens have been assigned to this taxon (Hou, 1997; Chiappe et al., 1999; Hou et al., 2002; Li et al., 2010). Hou (1997) later assigned V 10919 as the holotype of C. chuonzhous, which is considered as junior synonym of C. sanctus (see text below). V 10920-V 10925 preserve only traces of feathers, and their referrals to C. sanctus are questionable given that no morphological differences are present between the feathers of C. sanctus and other Mesozoic birds. V 10895 only preserves right hindlimb and part of the ilium, but the poor preservation makes its assignment problematic. We cautiously focused on specimens identified in previous studies that can be confidently assigned to C. sanctus, as well as some that we assign to C. sanctus for the first time here. A total of 20 specimens of C. sanctus are included in this study: V 10918 (Hou et al., 1995); V 11307, V 11370, V 12644, V 13156, V 13167, V 13168, V 13171, V 13172, V 13175, V 14373, V 14385, V 16066 (Li, 2010); GMV 2130-2133 (Chiappe et al., 1999); V 11374, V 11375, V 13178 (recognized here). These specimens are well preserved, and some overlapping elements are exposed in different views, allowing us to observe almost the entire skeletal morphology of this species.
According to Hou et al. (1995), C. sanctus is as large as the Eichst?tt specimen of Archaeopteryx. Discoveries subsequent to this study revealed that C. sanctus has a wide range of body sizes, with adult individuals generally being larger than the Eichst?tt Archaeopteryx. The second feature used to diagnose C. sanctus is the presence of a toothless beak (Fig. 1). This trait is shared with C. dui, Ch. hengdaoziensis and E. zhengi (Figs. 1, 5, 10), and thus is optimized to diagnose the entire Confuciusornithiformes, which evolved independently from the toothless seen in some enantiornithines and ornithuromorphs (Zhou et al., 2010). Hou et al. (1995) indicated that C. sanctus has a large orbit and a small antorbital fenestra. However, little difference in the proportional size of the orbit can be found between C. sanctus and other Early Cretaceous birds. Although the antorbital fenestra does appear to be relatively smaller than in other Early Cretaceous birds, this diagnostic character is not especially useful because the size of the fenestra cannot be accurately assessed in most taxa due to poor preservation of the delicate cranial elements.
Three putative features originally used to diagnose this taxon pertain to the manus (Hou et al., 1995). The first character, lack of fusion between the semilunate carpals and the metacarpals, has been clearly demonstrated by subsequent discoveries to be subject to ontogenetic variations with the proximal carpometacarpus becoming fully fused in adults (Figs. 8, 12). The two other manual characters are not unique to C. sanctus. The presence of three unreduced manual digits (manual phalangeal formula is 2-3-4) is a plesiomorphic avian condition, present in Archaeopteryx and Jeholornis prima, and characterizes all confuciusornithiforms (Figs. 4, 7, 8, 10, 12-14). The presence of a large recurved alular ungual phalanx similarly is not restricted to C. sanctus, being present in other confuciusornithiforms as well as in Early Cretaceous birds such as Sapeornis chaoyangensis and some members of the Enantiornithes. However, the alular ungual phalanx is proportionally larger in confuciusornithiforms than in other Mesozoic birds. Hou et al. (1995) described C. sanctus as having a unique ischium with a proximal dorsal process and a second distal dorsal expansion distally (V 11374, V 13168). This ischium morphology is present in other confuciusornithiforms and distinguishes this clade from other non-ornithothoracine birds. J. prima and S. chaoyangensis both possess a proximal dorsal process but lack the distal dorsal expansion, the distal margin of the ischium being blunt and weakly expanded in these taxa (Zhou and Zhang, 2002, 2003).
Fig. 3
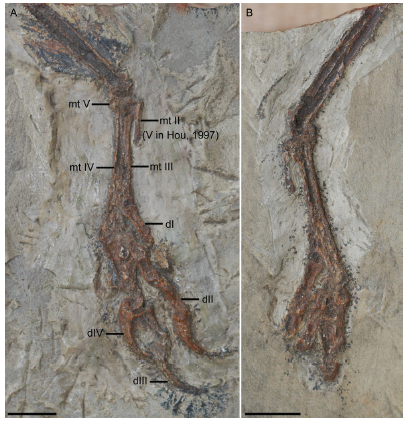 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3Photographs of Confuciusornis sanctus [= 'Confuciusornis chuonzhous'], IVPP V 10919
A. slab; B. counter slab. The broken piece of the proximal end of metatarsal II was misinterpreted as metatarsal V by Hou (1997). See text for further explanations. Scale bars = 10 mm
Due to the limited fossil record of Mesozoic birds known in 1995, most of the features initially used to diagnose C. sanctus have since proved to be invalid. The diagnosis has been revised in two subsequent studies (Chiappe et al., 1999; Li, 2010). However, some diagnostic characters were overlooked or misidentified in these works. Chiappe et al. (1999) described the presence of a single round foramen piercing the dorsal process of the maxilla, which they considered homologous to the maxillary fenestra in non-avian dinosaurs. C. sanctus was thus considered to document a transitional stage between the presence of both maxillary and promaxillary fenestrae in Archaeopteryx and the complete absence of fenestrae in the maxillae of derived birds. This feature was described from GMV 2130 (see fig.15 in Chiappe et al., 1999). However, Li (2010) suggested that the dorsal process described by Chiappe et al. (1999) is in fact the lacrimal. The skull in GMV 2130 is exposed in right lateral view and one of the elements forming the rostral margin of the orbit is perforated by a round foramen. Unfortunately, the overlapping cranial elements are heavily compressed, preventing clear identification of the perforated element (i.e., as the lacrimal or as the dorsal process of the maxilla). However, the dorsal process is clearly imperforate in the well-preserved maxillae visible in C. sanctus specimens V 13175, V 13156 and V 13172 (Fig. 1A-D). Meanwhile, the well-preserved lacrimal in V 13168 clearly demonstrates that this element is pierced by a round foramen (Fig. 1A, C). On the basis of these observations, we concur with Li (2010) that the dorsal process of the maxilla identified in Chiappe et al. (1999) is in fact the lacrimal. The dorsal process is imperforate and abruptly tapers dorsally with a broad base, giving this process a halberd-shaped appearance in lateral view (Fig. 1A-C), whereas this process is triangular in C. dui and rod-like in E. zhengi (Figs. 1D, 10B). In S. chaoyangensis the dorsal process is triangular and imperforate in lateral view (Wang Y et al., 2017), whereas in Jeholornis, most enantiornithines, and basal ornithuromorphs this process is longer, thinner and caudodorsally oriented (Zhou and Zhang, 2006b; O'Connor and Chiappe, 2011; Wang et al., 2017b; Fig. 1E, FAt this time, the halberd-shaped dorsal process of the maxilla is considered an autapomorphy of C. sanctus. In C. sanctus the surangular bears near its midpoint a triangular ventral process (Fig. 1B) that is absent in C. dui, and in other Mesozoic birds such as Archaeopteryx, S. chaoyangensis, Enantiornithes and basal Ornithuromorpha, and is here considered a diagnostic feature of C. sanctus.
Chiappe et al. (1999) considered the presence of a notch on the rostral margin of the premaxillary symphysis a diagnostic feature of C. sanctus, although in light of more recent discoveries this feature is most likely a shared feature of the Confuciusornithiformes. A rostrally notched premaxillary symphysis is present in the basal E. zhengi (Fig. 10B, C), but poor preservation unfortunately prevents observation of this feature in C. dui and Ch. hengdaoziensis (Figs. 1D, 5).
'Confuciusornis suniae' Hou, 1997
Remarks This species was described from the single specimen IVPP V 11308, a nearly complete individual preserved in a single slab, collected from the Yixian Formation at Sihetun Village, Beipiao City, Liaoning Province (He et al., 2004). Hou (1997) originally diagnosed 'C. suniae' as “distinguishable from C. sanctus by the following combination of characters: premaxillary symphysis notched rostrally; frontal process of the premaxilla elongated; large external naris; short frontal; large parietal; cervical vertebrae wide with small lateral excavation; neural spine of cervical vertebrae craniocaudally short and dorsoventrally low; thoracic vertebrae transversely short and craniocaudally long with deep lateral excavation; transverse processes of the caudalmost three thoracic vertebrae fused; transverse processes of the synscrals contacting the ilium; neural spines of the synscrals fused; and pygostyle fused”.
All the verifiable characters originally used to diagnose 'C. suniae' are shared with C. sanctus (e.g., elongated frontal process of the premaxilla) or widely distributed among Mesozoic birds (e.g., caudals fused into a pygostyle). Hou (1997) considered the premaxillary symphysis to be notched rostrally in 'C. suniae', but not C. sanctus. This is incorrect; the rostral notch is readily visible in C. sanctus specimens in which the premaxillary symphysis is exposed in dorsal or ventral view (Figs. 1B, 8B, 12B). According to Hou (1997), the frontal process of the premaxilla is longer in 'C. suniae' than in C. sanctus. In fact, the frontal processes are elongated and wedged between the rostral ends of the frontals in C. sanctus (Fig. 1A-C), a condition identical to that seen in 'C. suniae'. Hou (1997) additionally considered that the proportions of the external nares, frontals and parietals differ between 'C. suniae' and C. sanctus. Based on our observations, the subtle differences in the frontals and parietals that might be taken to separate V 11308 from C. sanctus specimens fall within the range of preservational variation seen in this species. The cranial elements in 'C. suniae' V 11308 are disarticulated so that the exact size of the nares cannot be determined, preventing reliable comparison of this feature with C. sanctus.
Proposed diagnostic differences in the cervical and thoracic vertebrae (Hou, 1997) are also unsupported. According to our observations, the cervicals and thoracics preserved in V 11308 are indistinguishable in all visible features (e.g., shared presence of lateral excavations on the thoracic centra) from those of C. sanctus. Hou (1997) described 'C. suniae' as having fused transverse processes in the caudalmost three thoracics and a synsacrum consisting of only four sacrals, whereas in C. sanctus the transverse processes are unfused and the synsacrum is formed by seven sacrals (Hou, 1997). These features are the result of misinterpretation of the proximal synsacral vertebrae as thoracics in 'C. suniae'. Reinterpretation of the caudal three thoracics as sacral vertebrae produces a synsacrum consisting of seven vertebrae, as in other confuciusornithiforms. The fusion of the neural spines of the sacral vertebrae into a spinous crest (Hou, 1997) is not a unique feature of 'C. suniae', being present in most Mesozoic birds including C. sanctus.
In our current review, none of the diagnostic features initially proposed for 'C. suniae' stand up to scrutiny. Given that 'C. suniae' exhibits many synapomorphies of C. sanctus, such as well-developed rostrally notched premaxillary and mandibular symphyses, deltopectoral crest triangular and perforated by an oval fenestra, proximal phalanx shorter than second phalanx in major digit, second phalanx of major digit bowed, and major digit claw strongly reduced, we conclude that 'C. suniae' is a junior synonym of C. sanctus.
'Confuciusornis chuonzhous' Hou, 1997
Remarks 'C. chuonzhous' was originally described by Hou (1997) as a “primitive bird diagnosed by the following characters: tibia robust without distal expansion; proximal tarsals unfused with tibia; metatarsal V unfused with other metatarsals; small hallux claw; and pedal claws less recurved than in other Mesozoic birds”. The holotype and the only known specimen of this taxon, IVPP V 10919, is incomplete, preserving only a partial hindlimb including the distal tibia, tarsals and the foot (Fig. 3), collected from the Yixian Formation deposits at the Huanbanjigou locality near Shangyuan Village, Beipiao City, in Liaoning Province (He et al., 2004).
Hou (1997) identified these elements as belonging to the right hindlimb, although our reexamination of V 10919 reveals that it is in fact the left hindlimb exposed in caudal view (Fig. 3). This misidentification was probably due to the severely crushed condition in which the bones are preserved. The proximal tarsals are not fused to the tibia in 'C. chuonzhous'. Initially, this was considered a valid diagnostic trait because the tibiotarsus was fully fused in specimens of C. sanctus known at the time (Hou et al., 1995). However, subsequent discoveries of numerous well preserved Early Cretaceous birds, including a plethora of confuciusornithiforms, coupled with osteohistological studies (Zhang et al., 1998; De Ricqlès et al., 2003), have since provided compelling evidence that the proximal tarsals and tibia became fully fused relatively late in basal bird ontogeny (Wang and Zhou, 2017b). Therefore, the unfused tibiotarsus in V 10919 only indicates the specimen is not an adult, rather than being a genuine diagnostic feature. According to Hou (1997) , a mediolaterally expanded distal tibia, a feature that characterizes C. sanctus, is absent in 'C. chuonzhous'. However, given that the distal expansion of the tibia results from fusion with the proximal tarsals, this feature is also subject to ontogenetic variation. Hou (1997) further argued that the tibia is more robust in 'C. chuonzhous' than in C. sanctus (4 mm vs. 2.5 mm, measured from the mediolateral width at the widest point of the tibia). However, this measurement is easily biased by preservation, given that the bones are crushed in 'C. chuonzhous'. Furthermore, the mediolateral width of the tibia of 'C. chuonzhous' V 10919 falls within the range of known specimens of C. sanctus: the mediolateral width measured at the level of the proximal margin of the medial condyle is 3.1 mm in V 10928, 4.5 mm in V 13171, and 5.4 mm in V 11552.
Hou (1997) diagnosed 'C. chuonzhous' as having a metatarsal V that is not fused to the rest of the tarsometatarsus (Fig. 3A), a feature that is now regarded as plesiomorphic for Aves. Furthermore, the element identified as metatarsal V by Hou (1997) is in fact the incomplete metatarsal II, as indicated by its medial position and robust size (Fig. 3A). We interpret a small fragment preserved lateral to the proximal tarsometatarsus as the proximal end of metatarsal V (Fig. 3A).
Hou (1997) describes the hallucal claw as being less than half the length of the other pedal claws in 'C. chuonzhous', and thus proportionally smaller than in C. sanctus. However, the hallucal claw is preserved in a severely deformed condition in 'C. chuonzhous', so that its length cannot be accurately measured. Furthermore, a short hallucal claw is also present in C. sanctus. According to Hou (1997), the pedal claws are less recurved in 'C. chuonzhous' than in C. sanctus. However, this apparent difference is due to the fact that the pedal claws in 'C. chuonzhous' V 10919 do not preserve their horny sheaths, features that are evident in many C. sanctus specimens and increase the overall curvature of the claws. We find little difference in the curvature of the ungual phalanges between 'C. chuonzhous' and other confuciusornithiforms (Figs. 2B, 3).
Collectively, the synapomorphies listed in Hou (1997) cannot distinguish 'C. chuonzhous' from other known confuciusornithiforms. Moreover, our examination failed to reveal any new diagnostic features, indicating that this taxon is invalid. Although incomplete and poorly preserved, V 10919 is morphologically more similar to confuciusornithiforms than to other contemporary birds. The pedal digits are more robust than in Archaeopteryx, enantiornithines and most Early Cretaceous ornithuromorphs, and the proportions are consistent with a confuciusornithiform identification. The hallux in V 10919 is proportionally shorter than in S. chaoyangensis and J. prima, but consistent with the length of this digit in other confuciusornithiforms. Given the incompleteness of the material and its poor preservation, we refer V 10919 to Confuciusornithidae incertae sedis.
Confuciusornis duiHou et al., 1999
Holotype IVPP V 11553.
Horizon and locality Libalanggou, Zhangjiying Village, Beipiao City, Liaoning Province; Yixian Formation (125 Ma; He et al., 2004).
Original diagnosis Following Hou et al. (1999), a confuciusornithiform that differs from C. sanctus in the following features: smaller body size; premaxilla more pointed rostrally than in C. sanctus; mandible slender without rostroventral expansion on the ventral margin of dentary; small alular digit claw; sternum more elongated than in C. sanctus with a rostral notch and a pair of short lateral processes; tarsometatarsus shorter than pygostyle, and proportionally shorter than in C. sanctus.
Revised diagnosis A small confuciusornithiform distinguishable through the following combination of characters: rostrum sharply pointed; dorsal process of maxilla triangular (halberd-shaped in C. sanctus); ventral margin of dentary straight (ventral convexity present in C. sanctus); surangular without a ventral process (present in C. sanctus and E. zhengi); deltopectoral crest pierced by suboval fenestra (absent in E. zhengi); and alular digit claw relatively small.
Remarks This taxon was described from two nearly complete specimens, V 11553 (holotype) and V 11521 (Hou et al., 1999). Unfortunately, neither of these specimens can be located at this time. However, given the importance of this species to confuciusornithiform diversity and the possibility that these specimens may yet resurface, we do not consider this taxon to be a nomen dubium. At this time our comparison is based on cast of V 11553 and published photographs (Hou et al., 1999, 2002). According to Hou et al. (1999) , C. dui is smaller than C. sanctus, the lengths of the humerus and femur being 42 and 35 mm, respectively (Supplementary Table 1). Although body size is not generally a good diagnostic character, in this case the small body size of C. dui is unlikely to be due to ontogenetic variation. The complete fusion of the carpometacarpus, tibiotarsus, and tarsometatarsus in V 11553 indicates that despite its small size it is subadult or adult. C. dui V 11553 is subequal in size to Ch. hengdaoziensis GMV 2129 (humerus and femur length approximately 33.5 mm) and E. zhengi V 11977 (in which compound bones are unfused; humerus length 41.3 mm, femur length 43.7 mm), but smaller than all the specimens of C. sanctus measured in this and previous studies (humerus length: 47.5-78.1 mm; femur length: 59.5-41.4 mm; Supplementary Table 1), including a subadult individual (V 13172) with an unfused carpus. Compared to other confuciusornithiforms, the skeleton of C. dui has a more gracile skeleton. At this time, we consider small body size to be a true diagnostic feature of C. dui.
Hou et al. (1999) described the rostrum in C. dui as more pointed than in C. sanctus (Fig. 1A-D). The frontal and maxillary processes of the premaxilla form an angle of 17° in C. dui, considerably smaller than in C. sanctus (e.g., 30° and 36.5° in the holotype V 10918 and V 13172, respectively). The angle cannot be measured in E. zhengi due to preservation (Fig. 10), or in Ch. hengdaoziensis because of its curved rostrum tip (Fig. 5). C. dui can be further distinguished from C. sanctus based on the morphology of the dentary. The ventral margin of the dentary is nearly straight in C. dui (Fig. 1D), without the characteristic ventral convexity present in C. sanctus (Chiappe et al., 1999; Fig. 1A) or in 'C. feducciai', 'J. yixianensis', and 'J. zhangjiyingia' (all considered here to be junior synonyms of C. sanctus, see discussion below; Figs. 7, 8,12). This feature cannot be assessed in 'C. jianchangensis' (also a junior synonym of C. sanctus, see discussion below), E. zhengi, or Ch. hengdaoziensis (Figs. 5, 10, 14).
Fig. 4
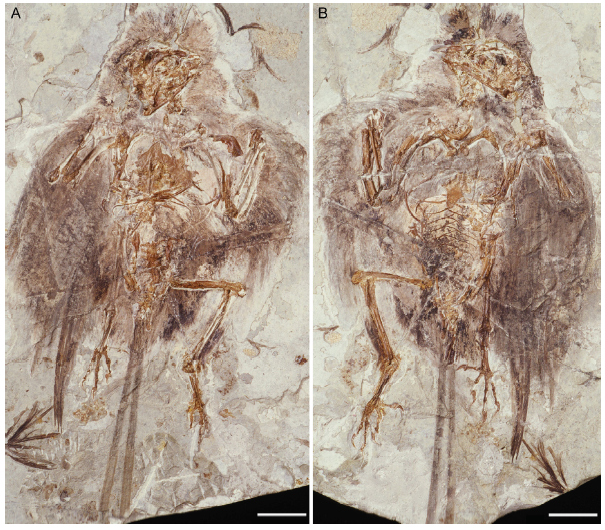 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 4Photograph of the holotype of Changchengornis hengdaoziensis, GMV 2129 Scale bars = 20 mm
The alular digit claw in C. dui is smaller than in C. sanctus (Hou et al., 1999). In C. dui, the alular digit claw is subequal in size to that of the minor digit, but it is much larger than the latter in C. sanctus, Ch. hengdaoziensis, and E. zhengi (Figs. 2A, 4, 10). Hou et al. (1999) considered the sternum of C. dui to be proportionally longer than in C. sanctus. In order to quantify this feature, we use the ratio of the width (the transverse distance between the proximal ends of the lateral processes) to the length of the sternal body. Based on the sternal reconstruction in Hou et al. (1999) , this ratio is 0.55 in C. dui V 11521, versus 0.59 in C. sanctus V 10928. This subtle difference fails to contribute substantively to the diagnosis of C. dui, given the potential for the calculated ratio to be affected by measurement error and individual variation. According to Hou et al. (1999) , the lateral processes of the sternum are triangular and project cranially in C. dui (Fig. 2E). Based on our observations of the cast of V 11553, the preserved morphology is comparable to C. sanctus: the craniolateral and caudolateral margins of the lateral processes project strongly craniolaterally and caudolaterally, respectively, and the lateral margin between them is concave (Fig. 2C, D), as opposed to semicircular without cranial and caudal projections in E. zhengi (Zhang et al., 2008; Fig. 2F). The last sternal trait originally used to distinguish C. dui from C. sanctus is the presence of a cranial notch on the midline of the sternal body. However, this trait is present in some specimens of C. sanctus (V 13313 and V 13156, and the holotypes of 'J. zhangjiyingia' and 'C. feducciai') (Figs. 8A, 12D), despite being absent in others (GMV 2130, see fig. 28 in Chiappe et al., 1999). Similar notches are present in subadult jeholornithiforms and some enantiornithines (e.g., Pengornithidae sp. STM 29-15, Eoalulavis hoyasi). O'Connor et al. (2015) argued that the cranial notch results from incomplete fusion between the sternal plates, and becomes closed later in ontogeny. Therefore, the presence of a cranial notch cannot be considered a synapomorphy of C. dui, but instead indicates that the holotype is a subadult. Hou et al. (1999) described the tarsometatarsus in C. dui as shorter than the pygostyle, and proportionally shorter than in C. sanctus. The length ratio between the pygostyle and tarsometatarsus is 1.18 in C. dui V 11553, which falls within the range observed in known specimens of C. sanctus (1.22 in GMV 2133, 1.02 in GMV 2131, and 1.08 in 'J. zhangjiyingia'), and thus fails to diagnose C. dui.
In addition to the characters originally proposed by Hou et al. (1999) , we identify two new cranial features diagnostic of C. dui: the dorsal process of the maxilla rapidly tapers dorsally, forming a triangular shape in lateral view (Fig. 1D), whereas the dorsal process is halberd-shaped in C. sanctus (Li, 2010; Fig. 1A-C) and rod-like in E. zhengi (Zhang et al., 2008); and the ventral margin of the surangular is straight in C. dui lacking the ventral process seen in C. sanctus and E. zhengi (Figs. 2B, 10C).
Changchengornis hengdaoziensisChiappe et al., 1999
Holotype GMV 2129.
Horizon and locality Jianshangou, Shangyuan Village, Beipiao City, Liaoning Province; Yixian Formation (125 Ma; He et al., 2004).
Original diagnosis Following Chiappe et al. (1999), a confuciusornithiform that differs from C. sanctus in the following features: rostrum curved ventrally; rostrum short relative to skull length, with a length ratio about 1/3 (compared with 1/2 in C. sanctus); mandible shorter than skull and increasing height caudally; clavicular rami having a horizontal 8-shaped cross section, compared to suboval in C. sanctus; furcular symphysis bearing a tubercle caudally; caudal margin of sternum more pointed than in C. sanctus; humeral deltopectoral crest imperforated; length ratio of alular and major metacarpals about 0.5; intermediate phalanx shorter than proximal phalanx in major digit; metatarsals III and IV fused distally; tarsometatarsus not excavated plantarly; and hallux proportionally longer than in C. sanctus.
Revised diagnosis A relatively small confuciusornithiform distinguishable from other confuciusornithiforms based on the following combination of features: tip of the rostrum curved ventrally; mandible shorter than skull and increasing in dorsoventral height caudally; clavicular symphysis with a caudal tubercle; alular metacarpal half the length of major metacarpal (1/3 in C. sanctus and E. zhengi); intermediate phalanx of major digit straight, shorter than proximal phalanx (opposite condition in C. sanctus and E. zhengi); tarsometatarsus without plantar excavation; and hallux more than half the length of pedal digit II, proportionally longer than in C. sanctus, C. dui, and E. zhengi.
RemarksCh. hengdaoziensis is known from a single nearly complete specimen preserved in a slab (GMV 2129a) and counter slab (GMV 2129b(Fig. 4). Chiappe et al. (1999) described the dorsal process of the maxilla as pierced by a round foramen in Ch. hengdaoziensis, resembling the purported condition in C. sanctus, and identified a splint-like fragment projecting cranioventrally from the dorsal margin of the orbit as the lacrimal. Our examination of Ch. hengdaoziensis suggests that these interpretations are incorrect. The so-called dorsal process is a fragment that lies ventral to the frontal process of the premaxilla, with no indication that it is associated with the body of the maxilla (Fig. 5). In GMV 2129b, in which the skull is preserved in left lateral view, there is a void in front of the “dorsal process” (labeled as “mx” in fig. 53 in Chiappe et al., 1999) that perfectly matches the shape of the nasal fragment preserved in GMV 2129a (labeled as “n” in fig. 52 in Chiappe et al., 1999), indicating that the void was left by the nasal (Fig. 5). It follows that the “lacrimal” (labeled as “l” in fig. 52 in Chiappe et al., 1999) situated caudal to the nasal must be the “dorsal process” labelled by Chiappe et al. (1999) as “mx” in their figure 53 preserved caudal to the void (left by the nasal) in the GMV 2129b (Fig. 5A, C). Therefore, if the original interpretation is correct a paradox would occur, with a single fragment representing the “dorsal process of the maxilla” in GMV 2129b but the “lacrimal” in GMV 2129a. Instead, we argue that the “lacrimal” identified by Chiappe et al. (1999) is only the caudal margin of the lacrimal, while the majority of the lacrimal is preserved in GMV 2129b and incorrectly interpreted as the “dorsal process” of the maxilla (Fig. 5). This reinterpretation is further supported by the fact that in C. sanctus the lacrimal is also perforated by a round foramen (see discussion above; Figs. 1A, C, 5).
Fig. 5
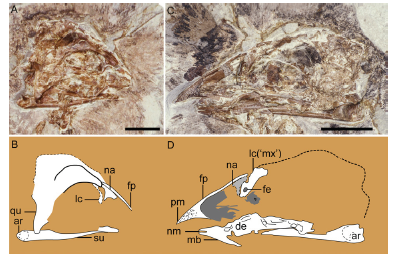 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 5Cranial morphology of Changchengornis hengdaoziensis, GMV 2129
A, B. skull in right lateral view preserved on GMV 2129a; C, D. skull in left lateral view preserved on GMV 2129b. A, C. photograph; B, D. line drawing. The dorsal process of the maxilla (labeled as 'mx') identified in Chiappe et al. (1999) is reinterpreted to be the lacrimal here (see text for explanation) Scale bars = 10 mm (A, C); line drawings (C, D) are not to scale
According to Chiappe et al. (1999), the dorsal surface of the furcula is excavated by a longitudinal furrow in Ch. hengdaoziensis, giving each ramus a C-shaped cross section (Fig. 6A) rather than an oval one as in C. sanctus. However, a comparable dorsal furrow is present in some specimens of C. sanctus, e.g., V 11370 and V 11521 (Fig. 6C), but absent in most indicating the caudal furrow may be the result of crushing. As such, this feature is no longer considered a diagnostic character of Ch. hengdaoziensis. Chiappe et al. (1999) described the caudal margin of the sternum of Ch. hengdaoziensis more pointed than in C. sanctus. However, we find only subtle differences between these two taxa (Figs. 2C, 6B). Another putative character used to diagnose Ch. hengdaoziensis is the absence of a fenestra in the deltopectoral crest of the humerus (Chiappe et al., 1999). However, in C. hengdaoziensis GMV 2129 the deltopectoral crest is incomplete in both humeri, including the area corresponding to the location of the fenestra (Fig. 6A). As such, unless new specimens with a complete deltopectoral crest are found, this character should not be used to diagnose Ch. hengdaoziensis.
'Jinzhouornis yixianensis' Hou et al., 2002
RemarksHou et al. (2002) erected the genus 'Jinzhouornis' and named the two species 'J. yixianensis' and 'J. zhangjiyingia', each represented by a single specimen. The holotype and only known specimen of 'J. yixianensis' is an incomplete specimen preserved in a single slab, IVPP V 14412 (Fig. 7), collected from Yixian Formation deposits near Wutun Village, Jinzhou City, Liaoning Province. According to the original diagnosis 'Jinzhouornis' is distinguishable from other Mesozoic birds in having the following features: skull low with antorbital region making up more than half the skull length; small orbit; cervical vertebrae shorter than in C. sanctus; more than 12 thoracic vertebrae; manual claws strongly recurved; major metacarpal and digit not expanded; scapula and humerus subequal in length; metatarsal II bearing a dorsal tubercle; and metatarsal III having a weak dorsal tubercle. The species 'J. yixianensis' is further diagnosed by the following features: elongated rostrum; small braincase; slender humerus; tarsometatarsus proximally fused; metatarsal V not fused to the other metatarsals; and proximal phalanx of digit III associated with a bony element proximally (Hou et al., 2002).
The cranial bones of 'J. yixianensis' V 14412 are heavily compressed and the bones forming the margins of the orbit and the antorbital fenestra have been lost (Fig. 7B). Thus, none of the cranial features used to diagnose this taxon can be verified. According to Hou et al. (2002), 'J. yixianensis' has more than 12 thoracic vertebrae, more than the 11 preserved in C. sanctus. However, the axial column is poorly preserved in 'J. yixianensis' V 14412 and the caudal thoracics are missing (Fig. 7A), making any count speculative. Hou et al. (2002) suggested that the humerus of 'J. yixianensis' is slenderer than that of C. sanctus. Interestingly, the bones are nearly three dimensionally preserved in 'J. yixianensis' V 14412, as revealed by the suboval cross sections exposed by breakages in some of the limb bones (Fig. 7). In contrast, the bones of most other confuciusornithiform specimens are compressed, increasing the apparent transverse widths of the limb bones. Despite these differences in preservation, we find only subtle differences in the measured relative robustness of the humerus between 'J. yixianensis' and C. sanctus.
Fig. 6
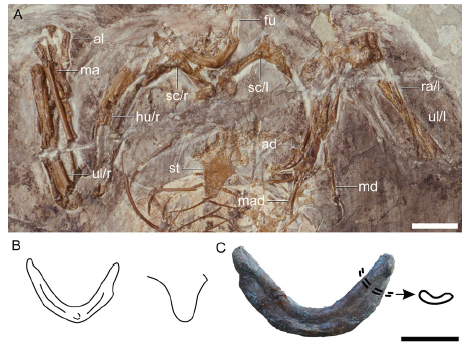 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 6Pectoral girdle of Changchengornis hengdaoziensis in comparison with Confuciusornis sanctus
A. pectoral girdle of Ch. hengdaoziensis in dorsal view; B. line drawing of the furcula (left) and caudal end of the sternum (right) of Ch. hengdaoziensis, showing that the dorsal surface is excavated by a dorsal furrow as in C. sanctus; C. furcula of C. sanctus IVPP V 11521 in dorsal view with drawing of the cross section Scale bars = 10 mm (A, C); line drawings (B, C) are not to scale
In our study we find no evidence to support the claim by Hou et al. (2002) that the manual claws of 'J. yixianensis' are more recurved than those of C. sanctus (Figs. 2A, 7). Similarly, purported differences in the width of the major metacarpal and digit between these species cannot be confirmed, especially in light of preservational biases. Hou et al. (2002) suggested that the scapula and humerus are subequal in length in 'J. yixianensis', whereas the former is shorter in C. sanctus. However, by our measurements the scapula (35.5 mm) is much shorter than the humerus (46.5 mm) in 'J. yixianensis' V 14412, as in C. sanctus. The proximal ends of metatarsals II and III each bear a dorsal tubercle in 'J. yixianensis' V 14412 (Fig. 7D), a feature which Hou et al. (2002) considered absent in C. sanctus. However, comparable tubercles are in fact present in C. sanctus (Fig. 2B), and considered to represent a diagnostic feature of this taxon (Chiappe et al., 1999). Hou et al. (2002) described 'J. yixianensis' as having metatarsals II-IV proximally fused and metatarsal V free, a condition shared with other confuciusornithiforms and widely distributed among Mesozoic birds (e.g., Jeholornithiformes, Sapeornithiformes, Enantiornithes, and the basalmost ornithuromorph Archaeorhynchus spathula; Chiappe and Walker, 2002; Zhou and Zhang, 2002, 2003; Wang and Zhou, 2017b). The last putative trait used to diagnose 'J. yixianensis' is the presence of a bony element proximal to the first phalanx of pedal digit III (Hou et al., 2002). This “element” is reinterpreted as a detached fragment of the proximal part of the first phalanx of digit III (Fig. 7D).
Fig. 7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 7Skeleton and anatomy of Confuciusornis sanctus [= 'Jinzhouornis yixianensis'], IVPP V 14412
A. overview; B. skull in oblique right lateral view; C. proximal left humerus showing the ridge on the cranial surface (arrow), as in C. sanctus; D. lower legs in dorsal view, exhibiting the presences of an extensor retinaculum on distal tibiotarsus (arrowhead), and the proximal ends of metatarsals II and III (arrows), as in C. sanctus. Scale bars = 10 mm
The original features used to diagnose 'J. yixianensis' are all found in the present study to be either preservational artifacts or widely distributed among confuciusornithiforms and/or other Mesozoic birds. Several features support referral of V 14412 to C. sanctus, such as a humerus with a triangular deltopectoral crest perforated by a suboval foramen (Fig. 7B); distal tibiotarsus with a tubercle for attachment of the extensor retinaculum; and short hallux measuring less than half the length of the second toe (Fig. 7C). In addition, the presence of a ridge along the proximodorsal margin of the cranial surface of the deltopectoral crest (Fig. 7B), and metatarsals II and III each bearing a proximally located dorsal tubercle (Fig. 7C), indirectly supports its referral to C. sanctus, although whether these features are present in other confuciusornithiforms cannot be determined due to preservation (a ridge on the proximal part of the humerus is present in C. hengdaoziensis). Therefore, we conclude that 'J. yixianensis' is a junior synonym of C. sanctus.
'Jinzhouornis zhangjiyingia' Hou et al., 2002
Remarks The holotype and only known specimen of 'J. zhangjiyingia', IVPP V 12352, is a nearly complete individual, preserved in a single slab (Fig. 8), collected from Yixian Formation deposits at Heitizigou in Zhangjiying Village, Beipiao City, Liaoning Province. Hou et al. (2002) stated that “in additional to the generic features, 'J. zhangjiyingia' is further characterized by the following features: frontal processes of premaxilla elongated and reaching rostral ends of frontals; large infratemporal fenestra; quadratojugal contributing to caudal boundary of orbit; small orbit; furcula slender than in C. sanctus; and humerus robust than in 'J. yixianensis'”.
As discussed above, the diagnostic features of the genus 'Jinzhouornis' are all either preservational artifacts or shared with C. sanctus. With regard to the specific characters initially proposed for 'J. zhangjiyingia', the first synapomorphy, the presence of a long frontal process of the premaxilla (Fig. 8B, C), is a feature widespread in the Confuciusornithiformes. In 'J. zhangjiyingia' V 12352 this process is identical in all aspects to that of C. sanctus: the frontal processes are unfused with each other medially, and extend caudally beyond the rostral margins of the frontals (Fig. 2A-C). According to Hou et al. (2002), the orbit and the infratemporal fenestra are both smaller in 'J. zhangjiyingia' than in C. sanctus. This putative size discrepancy is partly a result of preservation. The lacrimal, which forms the rostral margin of the orbit, is displaced caudally in 'J. zhangjiyingia' V 12352, consequently reducing the apparent size of the orbit. The postorbital region is exposed in oblique dorsal view, and the postorbital and the squamosal have been pushed ventrally so that they overlie the jugal and quadratojugal, precluding size assessment of the infratemporal fenestra (Fig. 8B, C). Hou et al. (2002) described the quadratojugal of 'J. zhangjiyingia' as extending rostrally well beyond the caudal end of the jugal, forming the caudoventral margin of the orbit (Fig. 8B); in contrast, the quadratojugal terminates well caudal to the caudal margin of the orbit in C. sanctus. This proposed cranial reconstruction for 'J. zhangjiyingia' is bizarre in that it contradicts the configuration of the postorbital region in all known Mesozoic birds and closely related non-avian theropod dinosaurs, in which the quadratojugal articulates with the distal end of the jugal and terminates caudal to the caudal boundary of the orbit (Sullivan and Xu, 2017; Wang and Hu, 2017). Reexamination of this specimen reveals that the postorbital elements, especially the jugal, quadratojugal, postorbital, and squamosal, are heavily compressed, making it difficult to identify the sutures between them. The ventral process of the postorbital can be readily discerned, separating the orbit from the infratemporal fenestra. This process is ventrally continuous with the putative “quadratojugal”, indicating that the “quadratojugal” is in fact part of the postorbital (Fig. 8C). A short element is preserved caudal to the distal end of the ventral process of the postorbital. The bone appears to be L-shaped, although its rostral and caudal ends are partially obscured by the postorbital rostrally and the squamosal caudally (Fig. 8C). We interpret this bone as the actual quadratojugal given its similarity in shape to the quadratojugals of C. sanctus and C. dui. Following this reinterpretation, the quadratojugal does not contribute to the caudal margin of the orbit, contra Hou et al. (2002), and the orbit of 'J. zhangjiyingia' is comparable to those of other confuciusornithiforms and Early Cretaceous birds.
Fig. 8
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 8Skeleton and cranial anatomy of Confuciusornis sanctus [='Jinzhouornis zhangjiyingia'], IVPP V 12352
A. overview; B, C. photograph (B) and line drawing (C) of the skull in left lateral viewThe arrowhead in B denotes the ventral end of the ventral process of the postorbital, which was misinterpreted as the quadratojugal by Hou (1997). Scale bars = 10 mm
Hou et al. (2002) further distinguished 'J. zhangjiyingia' from 'J. yixianensis' and C. sanctus based on the relative robustness of the furcula and humerus. However, only very subtle differences are observed in these features when 'J. zhangjiyingia' is compared to similarly sized specimens of C. sanctus such as GMV 2133. To quantify the robustness of the humerus, we calculated the ratio between the mediolateral width of the midshaft and the proximodsital length (8.7% in 'J. yixianensis' V 14412 and 11% in 'J. zhangjiyingia' V 12352). Given that the bones are three-dimensionally preserved in 'J. yixianensis' V 14412 and heavily compressed in 'J. zhangjiyingia' V 12352, this subtle difference is considered a preservational artifact.
None of the initially proposed synapomorphies of 'J. zhangjiyingia' are currently valid although there is no doubt that 'J. zhangjiyingia' is a confuciusornithiform. Our study of V 12352 reveals several morphological features that support referral to C. sanctus: the rostrum is robust with straight occlusal margins and the premaxillary symphysis is notched (Figs. 2A-C, 8B, C), in contrast to the gracile morphology in C. dui and the ventrally curved condition in Ch. hengdaoziensis (Figs. 2D, 5); the dorsal process of the maxilla is halberd-shaped, rather than triangular as in C. dui or rod-like as in E. zhengi (Figs. 2D, 10A, B); the ventral margin of the dentary is convex, rather than straight as in C. dui; the second phalanx of the major digit is longer than the proximal phalanx (Fig. 9A, B), opposite to the condition in Ch. hengdaoziensis; and the alular digit claw is the largest in the manus, the major digit claw being extremely reduced (Figs. 2A, 9A, B). The hindlimb morphology of 'J. zhangjiyingia' V 12352 is also comparable to that of C. sanctus: the distal tibiotarsus bears a tubercle, possibly providing attachment for an extensor retinaculum, on its dorsal surface (Fig. 9C, D); and the hallux is short, being less than half the length of pedal digit II (Chiappe et al., 1999). The presence of tubercles on both dorsal surfaces of metatarsals II and III also recalls the condition in C. sanctus. These observations indicate that 'J. zhangjiyingia' is a junior synonym of C. sanctus. Because both species of 'Jinzhouornis' are here found to be junior synonyms of C. sanctus, this genus can be considered a junior synonym of Confuciusornis.
Fig. 9
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 9Detail anatomy of Confuciusornis sanctus [= 'Jinzhouornis zhangjiyingia'], IVPP V 12352
A, B. pectoral girdle, sternum, and left forelimb; C, D. hindlimb showing the presence of an extensor retinaculum on distal tibiotarsus (arrow); A, C. photographs; B, D. line drawing. The arrow in A denotes the ridge on the cranial surface of the deltopectoral crest, as in C. sanctus. Scale bars = 10 mm
Eoconfuciusornis zhengiZhang et al., 2008
Holotype IVPP V 11977.
Horizon and locality Sichakou, Fengning County, Hebei Province, northeastern China; Huajiying Formation (130.7 Ma; He et al., 2006).
Original diagnosis Paraphrased from Zhang et al. (2008), a confuciusornithiform that is distinguishable from other confuciusornithiforms based on the combination of following features: thoracic vertebrae without lateral excavation; scapula without a prominent acromion and glenoid facet; coracoid short with relatively short articulation facet for sternum; deltopectoral crest small and imperforate; proximal end of humerus no more than twice as wide as its distal end; astragalus pierced by a foramina; and tarsometatarsus more than half the length of the tibia.
Revised diagnosis A small confuciusornithiform distinguished by the following unique combination of characters: rostrum sharply pointed as in C. dui (rather than stout as in C. sanctus or ventrally curved as in Ch. hengdaoziensis); dorsal process of maxilla craniocaudally compressed (as opposed to triangular in C. dui, and halberd-shaped in C. sanctus); surangular bearing a ventral process (absent in C. dui); thoracic vertebrae without lateral excavations; humeral deltopectoral crest imperforate (fenestrated in C. sanctus and C. dui; however, this feature is possibly subject to ontogenetic variation, as discussed below); and proximal phalanx shorter than intermediate phalanx in major digit (intermediate phalanx shorter in Ch. hengdaoziensis).
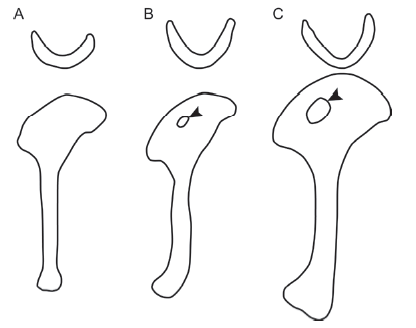
RemarksE. zhengi is the oldest known confuciusornithiform, initially described from a nearly complete specimen preserved in a slab and counterslab from the Protopteryx-horizon of the Huajiying Formation (Fig. 10). The holotype V 11977 is considered a subadult or possibly even younger individual, due to the readily visible incomplete ossification of the periosteal surfaces of the postcranial elements and the presence of free carpal and tarsal elements (Fig. 10). Zhang et al. (2008) originally described V 11977 as “nearly an adult” based on its size, considered to exceed that of other known adult confuciusornithiforms, and the presence of a pair of extremely long central tail feathers. Similar tail feathers in C. sanctus are considered a sexually dimorphic feature only present in males of the species. In extant birds, sexual traits typically do not express themselves until reproductive maturity, which follows skeletal maturity (Gill, 2007). Therefore, Zhang et al. (2008) considered V 11977 to be a reproductively mature adult or very near adult individual. However, according to our measurements V 11977 falls within the lower end of the size range of C. sanctus specimens measured in this study. The discovery of ovarian follicles in skeletally immature specimens of Jeholornis sp. and Enantiornithes indet. (Zheng et al., 2013), combined with histological evidence (O'Connor et al., 2014), and the presence of tail feathers in juvenile enantiornithines (Zheng et al., 2012), indicate that, unlike modern birds (Gill, 2007), basal taxa reached sexual maturity before skeletal maturity. As such, the presence of long tail feathers does not justify identification of the holotype of E. zhengi as a skeletally mature individual but does suggest that it is a male.
Reexamination of V 11977 suggests that some characters included in the original diagnosis in fact represent ontogenetic variation, such as the small size of the humeral deltopectoral crest. E. zhengi supposedly lacks a ridge on the proximal part of the cranial surface of the deltopectoral crest (Zhang et al., 2008), a feature present in C. sanctus and Ch. hengdaoziensis (Chiappe et al., 1999; Fig. 10D). Where present in confuciusornithiforms, this ridge runs along the proximal part of the cranial surface of the deltopectoral crest (Figs. 2A, 7C, 8A). However, in V 17911, the bone surface corresponding to this margin is not fully ossified, as indicated by the coarse and heavily pitted periosteum (Fig. 10D), making it possible that this ridge was simply unossified at the time of death. According to Zhang et al. (2008), the scapula of E. zhengi lacks both an acromion process and a glenoid facet. We are skeptical about this description given the widespread presence of these features in confuciusornithiforms and other Mesozoic birds (e.g., J. prima and S. chaoyangensis). The periosteum of the proximal part of the scapula is coarse and extensively pitted. In modern birds, the proximal end of the scapula ossifies after the central portion of the scapular body (Maxwell, 2008). Therefore, it is possible that the acromion process had not yet ossified in V 17911. The coracoid and scapula are fused into a scapulocoracoid in E. zhengi and other confuciusornithiforms (Figs. 2A, 6A, 7, 9) and the glenoid facet is located on the lateroventral surface of the proximal scapula. In V 17911 the scapulocoraocid is laterally compressed, which may be obscuring the glenoid.
Zhang et al. (2008) described the tarsometatarsus of E. zhengi as proportionally longer than that of C. sanctus. The ratio of the length of the tarsometatarsus to that of the tibia is 0.51 in V 17911, falling within the range seen in C. sanctus specimens sampled here (0.46-0.52; Supplementary Table 1). Regardless, in light of the fact that V 17911 is skeletally immature, limb proportions cannot be considered valid diagnostic features. Zhang et al. (2008) considered the sternal articulation of the coracoid to be relatively short in E. zhengi. However, only very subtle differences, which we do not consider to be diagnostic, exist between E. zhengi and other confuciusornithiforms.
In E. zhengi the astragalus is pierced by a foramen, and this feature is described as comparable to the opening of the extensor sulcus of the tibiotarsus in modern birds (Zhang et al., 2008). Given that the astragalus and calcaneum are fused neither to each other nor to the tibia in E. zhengi V 17911, this homology is probably incorrect. The periosteal surface of the proximal tarsals is poorly ossified. Without additional specimens we cannot determine whether or not the presence of a foramen in the astragalus is subject to ontogenetic variation.
Despite the skeletal immaturity of the specimen, we identify a few features that we consider less subject to ontogenetic variation and therefore diagnostic of E. zhengi. The rostrum is more pointed than in C. sanctus (but not as sharply pointed as in C. dui, Figs. 2A-D, 10A, B) and the premaxillary corpus is proportionately smaller (Zheng et al., 2017). The dorsal process of the maxilla is craniocaudally thin and rod-like (Fig. 10C), rather than triangular as in C. dui or halberd-shaped as in C. sanctus (Fig. 2C, D). As in C. sanctus, the surangular bears a ventral process (Fig. 10C), a structure absent in C. dui. The manual proportions are also taxonomically significant. As in C. sanctus the proximal phalanx of the major digit is shorter than the intermediate phalanx, but the opposite is true in Ch. hengdaoziensis. The size disparity of the manual unguals is weaker compared to younger confuciusornithiforms; the major ungual is 70% the size of the minor ungual compared to 50% in C. sanctus. One additional feature may be more controversial, namely the purported absence of a fenestra in the deltopectoral crest of the humerus (Fig. 11A). It has been previously suggested that this fenestra is a size related feature that appears fairly late in ontogeny. Although even the smallest known subadult specimens of C. sanctus (e.g., V 13172) preserve this feature. Still, we cannot rule out the possibility that the fenestra may have developed in E. zhengi V 11977 at a relatively later ontogenetic stage than in C. sanctus. Until new material is discovered, we suggest that E. zhengi can be diagnosed as having an imperforate deltopectoral crest at least in subadults, thus still differentiating it from C. sanctus (in which fenestra present is present in subadults).
Eoconfuciusornis sp.
Referred specimens STM7-144, BMNHC-PH870.
Remarks Recently, two new specimens (STM7-144, BMNHC-PH870) from the Protopteryx-horizon have been referred to Eoconfuciusornis (Zheng et al., 2017; Navalón et al., 2018). STM7-144 differs from other known confuciusornithiforms but resembles E. zhengi IVPP V 119177 in having a proportionately small premaxillary corpus and a humerus with an imperforate deltopectoral crest. Based on hindlimb elements, STM7-144 is 83%-87% the size (here referring to linear dimensions of long bones) of the holotype of E. zhengi. Despite its smaller size, the relatively greater degree of sternal ossification in STM7-144 suggests this specimen is more skeletally mature than V 11977 is less skeletally mature (Zheng et al., 2017). Preserved remains of the ovaries indicate STM7-144 is a female, consistent with the absence of elongate tail feathers, and the difference in body size between this specimen and V 11977 may be related to sexual dimorphism. Despite the fact that the plumage is preserved with great fidelity, the bones are poorly preserved in STM7-144. Still, some morphologies clearly differ from those seen in the holotype of E. zhengi, such as the proportionately smaller forelimb (roughly 75%-80% as long as that of V 11977) and subequal minor and major digit unguals (minor digit ungual is larger in V 11977, although not enlarged to the extent observed in C. sanctus) in STM7-144 (Zheng et al., 2017). Currently, observed differences between the two specimens could be interpreted to reflect ontogenetic variation, sexual dimorphism, and/or taxonomic diversity; given this uncertainty we concur with Zheng et al. (2017) and refer STM7-144 to Eoconfuciusornis indet.
Using humeral length as a proxy for body size, BMNHC-PH870 is approximately 10% larger than V 17911 in terms of linear dimensions. Navalón et al. (2018) concluded that BMNHC-PH870 is an adult individual given the presence of fully fused compound elements including the carpometacarpus, tibiotarsus and tarsometatarsus. BMNHC-PH870 was considered morphologically more similar to E. zhengi than to other confuciusornithiforms in having a slender humerus with a less dorsally projecting deltopectoral crest (Navalón et al., 2018). However, the authors also noted that BMNHC-PH870 differs from E. zhengi V 11977 in having a smaller interclavicular angle (64° compared with 78° in V 11977) and a perforated humeral deltopectoral crest. Based on our observations (superimposing photographs of both furculae) the furculae are nearly identical in these two specimens. Thus, the only major difference between V 17911 and BMNHC-PH870 lies in the presence of a small suboval fenestra in the humeral deltopectoral crest of the latter (Fig. 11A, B). The fenestra is approximately one half to one third the diameter of the similar feature in C. sanctus (Fig. 11B, C). The presence and size of this fenestra may be affected by ontogenetic variation. Although we do not have any direct information regarding the development of the deltopectoral crest in E. zhengi, information from C. sanctus may shed some light on this issue. In C. sanctus, the fenestra is consistently developed even in the most skeletally immature individuals such as V 13172, in which the carpometacarpus is unfused as in E. zhengi V 17911 (Fig. 2A). Similarly, this foramen is present in subadult S. chaoyangensis (Gao et al., 2012). These observations suggest that this feature occurs relatively early in ontogeny in at least some taxa, and its absence in E. zhengi could be genuine. However, no small juvenile specimens are known for either C. sanctus or S. chaoyangensis. Therefore, a viable alternative is that the absence of this foramen in E. zhengi V 17911 is in fact ontogenetic, and the deltopectoral crest is perforated relatively late in ontogeny. In that case, BMNHC-PH870 would most likely represent an adult individual of E. zhengi. This would suggest that the humeral fenestra appears later in ontogeny in E. zhengi than in derived confuciusornithiforms. Obviously, more specimens that record different developmental stages, particularly juveniles or hatchlings, will be needed to clarify this issue.
Fig. 10
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 10Skeletal anatomy of the holotype of Eoconfuciusornis zhengi, IVPP V 11977
A. overview; B, C. photograph (B) and line drawing (C) of the skull; D. furcula (dorsal view) and proximal part of right humerus (cranial view). The arrowhead in C indicates that the surangular bears a ventral process as in C. sanctus. The arrow in C shows that the dorsal process of the maxilla is craniocaudally compressed Scale bars = 10 mm (A, B, D); line drawing (C) is not to scale
Fig. 11
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 11Comparison of furcula and humerus anatomy between Eoconfuciusornis zhengi and Confuciusornis sanctus
A. holotype of Eoconfuciusornis zhengi, IVPP V 11977; B. BMNHC-PH870, referred to E. zhengi in Navalón et al. (2018); C. C. sanctus. The arrowheads in B and C indicate that the deltopectoral crest is perforated by a suboval fenestra in BMNHC-PH870 and C. sanctus, but not in E. zhengi. Line drawings are not to scale
The skeletal immaturity of the holotype of E. zhengi and the poor preservation of BMNHC-PH870 make comparisons between these specimens difficult; nevertheless, several small morphological differences are noted, in addition to the discrepancy in humeral morphology. In BMNHC-PH870 metatarsal III is much wider than metatarsals II and IV as in STM7-144, whereas all these metatarsals are subequal in in width C. sanctus and E. zhengi; metatarsal II terminates more proximally relative to metatarsal IV in BMNHC-PH870 and E. zhengi than in C. sanctus; the major digit ungual is approximately half the size of that of the minor digit in BMNHC-PH870, whereas the corresponding ratio is 0.7 in STM7-144. However, without more material or better understanding of ontogenetic variation and potentially sexual dimorphism in early confuciusornithiforms it is inappropriate to erect a new taxon based on this specimen at this point. We suggest referring BMNHC-PH870 to Eoconfuciusornis sp., or Confuciusornithiformes incertae sedis.
'Confuciusornis feducciai' Zhang et al., 2009
Remarks The holotype and the only known specimen of 'C. feducciai', DNHM D2454, is nearly complete and preserved in a single slab (Fig. 12A), collected from Yixian Formation deposits at Sihetun, Shangyuan Village, Beipiao City, Liaoning Province (Zhang et al., 2009). This taxon was originally diagnosed as “a confuciusornithiform that is distinguishable from C. sanctus in the following features: large body size; forelimb elongated with a length ratio (humerus+ulna+carpometacarpus)/(femur+tibiotarsus+tarsometatarsus) approximately 1.15; proximal end of humerus flat and thin; deltopectoral crest rectangular and imperforated; proximal phalanx of alular digit slender; furcula V-shaped; sternum wider than long; and length ratio of ischium/pubis about 2/3” (Zhang et al., 2009).
Zhang et al. (2009) described the humeral deltopectoral crest of 'C. feducciai' as rectangular and imperforate, in contrast to the triangular and perforated condition in C. sanctus. However, reexamination of the holotype of 'C. feducciai' reveals that the deltopectoral crests of both humeri are clearly perforated (Fig. 13A, B), as in C. sanctus (Figs. 2A, 7, 9) and we find negligible differences in the shape of this crest between these two taxa. According to Zhang et al. (2009), the proximal phalanx of the alular digit is more slender than in C. sanctus, and the claw of the alular digit is as large as that of the minor digit, rather than being the largest in the hand as in C. sanctus. This description is incorrect; contra Zhang et al. (2009) we find the manual morphology to be identical in 'C. feducciai' and C. sanctus (Figs. 2A, 9A, B, 13C). One putative difference between 'C. feducciai' and C. sanctus is the presence of a V-shaped furcula in 'C. feducciai', in contrast to the U-shaped condition in C. sanctus. This difference is a preservational artifact resulting from overlap with the cervical vertebrae, which causes the clavicular symphysis to appear elevated relative to the clavicular rami (Fig. 12C). Zhang et al. (2009) considered the sternum to be wider than long in 'C. feducciai', opposite to the condition in C. sanctus. However, their sternal reconstruction of 'C. feducciai' is problematic; the caudal margin of the sternum is in fact missing in DNHM D2454, making it impossible to estimate the proportions of this element (Fig. 12D).
The ischium is described as proportionally longer than in 'C. feducciai', with the ischium to pubis length ratio reported as 2/3 (Zhang et al., 2009) compared with 0.44 in C. sanctus. However, the length of the ischium cannot be measured accurately in 'C. feducciai'; the proximal and distal ends of the bone are obscured by the pubis on both sides of the skeleton (12E). Zhang et al. (2009) diagnosed 'C. feducciai' as larger than C. sanctus; with a humeral length of 80.1 mm. It is true that the holotype of 'C. feducciai' is one of the largest known confuciusornithiforms, being considerably larger than the specimens of C. sanctus measured in this study (humeral length 69.5-47.5 mm). However, given the considerable size range observed in C. sanctus, 'C. feducciai' could conceivably fall within the growth trajectory of C. sanctus, as suggested in a recent morphometric study (Marugán-Lobón et al., 2011). Zhang et al. (2009) describe the forelimb as proportionally longer in 'C. feducciai' than in C. sanctus. The intermembral index (humerus+ulna+carpometacarpus)/(femur+tibiotarsus+tarso-metatarsus) is 1.15 in 'C. feducciai' (Zhang et al., 2009), compared to 0.87-1.09 in C. sanctus specimens studied here (Supplementary Table 1). However, advanced ontogenetic status may explain the proportionally longer forelimb in 'C. feducciai'.
Interestingly, although 'C. feducciai' is the largest known confuciusornithiform, the two dentaries are unfused rostrally this species (Fig. 12B). Li (2010) noticed that the degree of fusion of the dentaries varies in C. sanctus, apparently in a size-independent manner. For instance, the dentaries are completely fused in IVPP V 13156 (humeral length 61.2 mm) and V 13171 (humeral length 62.7 mm), but not in V 13178 (humerus length 66.4 mm), suggesting intraspecific variation in this feature. Our examination of DNHM D2454 fails to identify clear morphologies capable of supporting 'C. feducciai' as a valid taxon, although the specimen exhibits numerous synapomorphies of C. sanctus: robust mandible with rostral part of ventral margin convex (Fig. 12B); mandibular symphysis notched rostrally (Fig. 12B); humerus expanded proximally with large, triangular deltopectoral crest; deltopectoral crest perforated by a suboval fenestra (Fig. 13A, B); penultimate phalanx of the major digit bowed and longer than the preceding phalanx (Fig. 13C); and hallux short, less than half the length of the second toe. Consistent with its referral to C. sanctus, 'C. feducciai' preserves a ridge on the proximal part of the cranial surface of the deltopectoral crest, and tubercles on the dorsal facet of both metatarsals II and III (Fig. 13D). At this time, we consider the difference in body size between 'C. feducciai' and undoubted specimens of C. sanctus, and 0.06 difference in limb proportions, to be within the scope of ontogenetic variation. Therefore, we conclude that 'C. feducciai' is a junior synonym of C. sanctus and that DNHM D2454 may be one of the most mature known specimens of this species.
Fig. 12
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 12Skeleton and anatomy of Confuciusornis sanctus [= 'Confuciusornis feducciai'], DNHM D2454
A. overview; B. skull in ventral view, showing that the mandibular symphysis is notched rostrally (arrow); C. furcula in ventral view; D. sternum in ventral view, showing the incomplete caudal end (dashed line) and a cleft at the midpoint of the rostral margin (arrow); E. pelvis in ventral viewScale bars equal 20 mm in A, 10 mm in B-E
Fig. 13
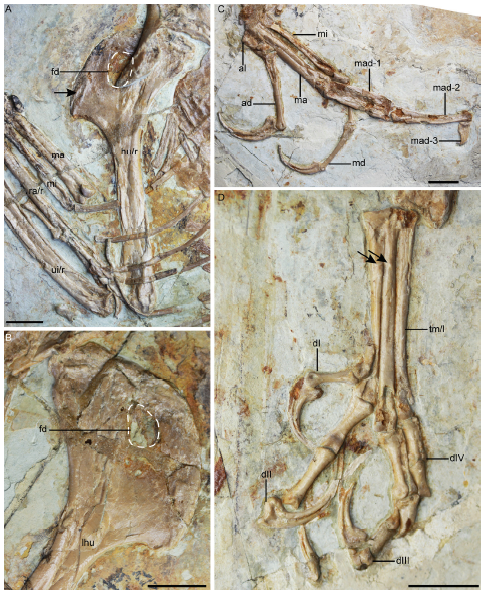 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 13Detail anatomy of Confuciusornis sanctus [= 'Confuciusornis feducciai'], DNHM D2454, showing that it is morphologically identical to C. sanctus
A. right humerus in cranial view showing the ridge on the proximal part of the cranial surface of the deltopectoral crest (arrow); B. proximal left humerus in cranial view showing the perforated deltopectoral crest; C. dorsal view of metacarpals; D. dorsal view of metatarsals The arrows in D denote the dorsal tubercles on metatarsals II and III. Scale bars = 10 mm
'Confuciusornis jianchangensis' Li et al., 2010
Remarks This taxon was described based on PMOL-AB00114, an incomplete skeleton missing the forelimb and pectoral girdle (Fig. 14A) collected from the Jiufotang Formation near Toudaoyingzi Village, Jianchang County, Huludao City, Liaoning Province. It is one of the few confuciusornithiform specimens to have been collected from the Jiufotang Formation. The original diagnosis states 'C. jianchangensis' is “a small confuciusornithiform that is distinguishable in having the follow features: teeth reduced; triangular squamosal; large quadrate; dentary slender and elongated with convex dorsal margin; femur/tibiotarsus length ratio about 0.89; metatarsals II-IV fused proximally; metatarsal V absent; and pygostyle large” (Li et al., 2010).
Most of these features used to diagnose 'C. jianchangensis' are present in all confucius-ornithiforms (e.g., toothless upper and lower jaws) and/or widely distributed in Mesozoic birds (e.g., proximally fused metatarsals II-IV present in other confuciusornithiforms, J. prima, S. chaoyangensis, and the Enantiornithes). The squamosal is described as triangular and the quadrate as broad in 'C. jianchangensis' (Li et al., 2010), both features consistent with referral to C. sanctus. Li et al. (2010) described the dentary as slender with a convex dorsal margin; this interpretation is incorrect. The skull is preserved obliquely in right dorsolateral view, giving the dentary a slightly more delicate appearance than it would have in strict lateral view, and only the right dentary is partially exposed (Fig. 14B). The presence of a convex dorsal margin in the dentary is not supported here, because this element has been severely deformed. The length ratio between the femur and tibiotarsus is 0.89 in 'C. jianchangensis' (Li et al., 2010). This falls within the range of C. sanctus specimens included in this study (0.81-0.91). Li et al. (2010) considered the absence of metatarsal V (which is present in C. sanctus) to be a diagnostic feature of 'C. jianchangensis'. However, this small splint-like element is not fused to the other metatarsals and is often not preserved even in articulated specimens, e.g., C. sanctus IVPP V 11552 (Fig. 2B), 'C. feducciai' (Fig. 13D), GMV 2141 and GMV 2130 (see figs. 44, 45 in Chiappe et al., 1999), indicating that its absence in 'C. jianchangensis' could be a taphonomic artifact. The last putative trait used to diagnose 'C. jianchangensis' is the presence of a large pygostyle. However, the pygostyle in 'C. jianchangensis' appears identical to that of C. sanctus (for instance, the pygostyle/tarsometatarsus length ratio is 1.20 in 'C. jianchangensis', compared with 1.22 in GMV 2133 and 1.02 in GMV 2131; Fig. 14A).
Fig. 14
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 14Skeleton and anatomy of Confuciusornis sanctus [='Confuciusornis jianchangensis'], PMOL-AB00114
A. overview; B. skull; C. thoracic vertebrae showing that the centra are excavated by lateral suboval fossae (arrow), as in C. sanctus; D. hindlimb. The arrows in D denote the dorsal tubercles on metatarsals II and III Scale bars = 20 mm
The original diagnosis fails to distinguish 'C. jianchangensis' from C. sanctus, and our reexamination indicates that the former putative taxon is very morphologically similar to C. sanctus. Several diagnostic characters of C. sanctus can be clearly observed in PMOL-AB00114, including a tibiotarsus with a distal tubercle for the extensor retinaculum, and a short hallux (Fig. 14B-D). In addition, the presence tubercles on the dorsal surfaces of both metatarsals II and III inderectly supports referral to C. sanctus. Therefore, we argue that 'C. jianchangensis' is a junior synonym of C. sanctus.
4 Phylogenetic analysis
In order to explore interrelationships within the Confuciusornithiformes and the syste-matic position of this clade with respect to other Mesozoic birds, a comprehensive phylo-genetic analysis was performed. We formulated a new character matrix based on the most recent phylogenetic studies (Wang et al., 2014, 2017a , b; Hu and O'Connor, 2017), with extensive modifications including revisions of character scorings and the addition of new taxa and characters. We added 18 new characters (numbered from 263 to 280; Supplementary Information) to the data matrix in Wang et al. (2017a), which originally contained 262 characters and 59 taxa. We added nine recently described Mesozoic birds, including Chongmingia zhengi, Elsornis keni, Dunhuangia cuii, Piscivorenantiornis inusitatus, Linyiornis amoena, Pterygornis dapingfangensis, Monoenantiornis sihedangia, Bellulornis rectusunguis, and Iteravis huchzermeyeri. Character scorings for these taxa follow published studies (Hu and O'Connor, 2017; Wang et al., 2017a). In addition, many previously missing data have been filled in on the basis of new referred specimens discovered subsequent to the initial descriptions of the taxa in question, including Archaeorhynchus spathula (IVPP V 17091, V 17075, and V 20312; Zhou et al., 2013; Wang and Zhou, 2017b), Pterygornis dapingfangensis (V 16363; Wang et al., 2017b), and Vegavis iaai (MACN-PV 19.748; Agnolín et al., 2017).The dataset was analyzed using the TNT software package (Goloboff and Catalano, 2016), with the following settings: all characters were weighted equally, and 35 characters were ordered; an unconstrained heuristic search was used; 1000 replicates of random stepwise addition (branch swapping trees: tree-bisection-reconnection, TBR), with ten trees kept at each step, were carried out in the first round, and the second round of searching was performed using the trees from memory. We calculated Bootstrap and Bremer values as indices as support. Bootstrap values were calculated by conducting 1000 replicates in TNT using the default settings. Bremer values were calculated using the Bremer script embedded in the TNT software.
The phylogenetic analysis produced 144 most parsimonious trees, and in the strict consensus tree all the sampled confuciusornithiforms were resolved together in a clade (Fig. 15), the Confuciusornithiformes, which constituted the direct outgroup to the Sapeornis + Ornithothoraces clade. C. dui, C. hengdaoziensis, and E. zhengi formed the successive outgroup to C. sanctus + 'J. zhangjiyingia' clade. All characters that can be scored for C. sanctus and 'J. zhangjiyingia' are identical, confirming that 'J. zhangjiyingia' is a junior synonym of the C. sanctus. The resolved topology of the Confuciusornithiformes supports the monophyly of the genus Confuciusornis by grouping C. dui and C. sanctus together. Based on this analysis, the Confuciusornithiformes may be phylogenetically defined as the most-inclusive clade that contains E. zhengi Zhang et al., 2008, but not Sapeornis chaoyangensis Zhou & Zhang, 2002, or Passer domesticus Linnaeus, 1758. Although the interrelationships within confuciusornithiforms were completely resolved in the new topology, all the clades received low support values. Specifically, all the nodes had frequency values less than 50% in the bootstrapping analysis, and the Bremer values were 1 or 2 for the internal clades. The low levels of support can be ascribed to multiple factors or a combination of them: except C. sanctus, other taxa are represented by one or two specimens that are poorly preserved, severely limiting the amount of characters that can be identified; moreover, because most of the confuciusornithiform specimens are preserved in two-dimensions, some characters (e.g., cranial elements) are obscured, causing abundant missing entries in the phylogenetic analysis; thirdly, these low support values may due to homoplasy which has been argued to be widely present in early avian evolution (Zheng et al., 2012; O'Connor et al., 2015; Wang et al., 2017a); and finally, the number of characters used in present and recent phylogenetic analyses of Mesozoic birds are relatively small, and thus are not able to adequately resolve the entire range of morphology of Mesozoic birds, given the fact that the increase of new taxon appears to outpace that of morphological characters. As such, further data, particularly with applications of CT scanning that can obtain more morphologies, is needed in order to develop a more robust hypothesis of confuciusornithiform interrelationships.
Fig. 15
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 15Cladogram of Mesozoic birds showing interrelationships within the Confuciusornithiformes
The strict consensus tree is built from 144 most parsimonious trees (Length = 1200; Consistency index = 0.324; Retention index = 0.673) that were found in the present phylogenetic analysis Bootstrap and Bremer values are indicated in bold and italic fonts near the corresponding nodes, respectively
5 Conclusion
Reexamination of all currently recognized species of the Confuciusornithiformes supports widespread conjecture in the literature that most taxa are invalid due to inadequate diagnoses, based on characters that are either widely distributed among confuciusornithiforms, taphonomic artifacts, points of intraspecific variation, or simply misinterpreted. 'C. feducciai', 'C. suniae', 'J. yixianensis', 'J. zhangjiyingia', and 'C. jianchangensis' should be regarded as junior synonyms of C. sanctus. 'C. chuonzhous' is a nomen dubium and should be referred to Confuciusornithiformes incertae sedis. The recently described BMNHC-PH870 is certainly a confuciusornithiform, but cannot be adequately diagnosed at this time without the discovery of more material to clarify potential ontogenetic differences between this specimen and other confuciusornithiforms from the Huajiying Formation. Therefore, BMNHC-PH870 is regarded as Confuciusornithiformes incertae sedis. Under the present taxonomic revision, the Confuciusornithiformes consists of one family, three genera, and four species, C. sanctus, C. dui, Ch. hengdaoziensis, and E. zhengi. The actual diversity of the Confuciusornithiformes is in all probability higher. However, the identification of new species needs to be well supported by valid diagnostic features that clearly distinguish the putative new taxa from existing confuciusornithiforms. This taxonomic revision clarifies the current diversity and provides a launching point for future studies attempting to tackle the thousands of unstudied confuciusornithiforms in collections throughout China.Acknowledgments
We thank Corwin Sullivan and Li Zhiheng for their constructive comments on this manuscript. We thank Gao Wei for photography; Luis M. Chiappe and Hu Dongyu for sharing specimen photographs; and Gao Chunling for facilitating access to the holotype specimen of ‘Confuciusornis feducciai’. This work was supported by National Natural Science Foundation of China (41722202, 41502002, 41688103).参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 29]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 2]
DOIURL [本文引用: 1]
[本文引用: 7]
[本文引用: 2]
[本文引用: 20]
[本文引用: 12]
DOIURL [本文引用: 13]
[本文引用: 19]
DOIURL [本文引用: 2]
[本文引用: 15]
DOIURL [本文引用: 2]

Confuciusornis sanctus stands out among the remarkable diversity of birds of the Jehol biota (Lower Cretaceous, Liaoning Province, China). Its basal position in the phylogenetic tree of birds, combined with the exceptional number of well-preserved, largely complete and articulated specimens, makes it a perfect model system for studying the variation, development and life history of early Mesozoic birds. A comprehensive morphometric study (measurements of humerus, ulna, radius, femur and tibia) previously identified two distinct size classes of C. sanctus, while demonstrating the lack of statistical support of the association between this size dimorphism and the characteristic pair of long tail feathers present in some specimens. Four plausible explanations were discussed to account for the resultant size classes: the existence of more than one species in the sample; sexual size dimorphism; two size classes corresponding to attritional death assemblages; and/or a particular growth pattern similar to that inferred for non-avian dinosaurs. Here we present an expanded statistical analysis based on a larger sample of C. sanctus that substantiates previous interpretations, but also addresses the statistical association between the presence/absence of tail feathers and fore- and hind-limb allometry. We discuss the implication of the resulting quantitative patterns of morphological variation to understand better confuciusornithid taxonomy and the life history of C. sanctus.
DOIPMID [本文引用: 1]
Ossification sequences are poorly known for birds in general, even for common domestic and experimental species. Such sequences constitute a rich source of data on character evolution, and may even provide phylogenetic information. It is not clear, however, what factors influence ossification sequences and what the relative importance of phylogeny is to the sequences. Galliformes constitute a good group to examine these variables. These birds are osteologically conservative, have precocial young, but have a broad spectrum of body sizes and incubation periods. Here, I describe the embryonic ossification of the skeleton in the domestic turkey (Meleagris gallopavo), and compare it to the domestic chicken (Gallus gallus) and the Japanese quail (Coturnix coturnix). Ossification sequences in this group are not affected by egg size or incubation period. They also appear to be independent of both the spatial location and the embryonic tissue from which the osteogenic cells originated. Accumulation of a wider sample of ossification sequences from more morphologically variable avian taxa will be necessary in order to test functional and phylogenetic effects.
DOIURL [本文引用: 5]
DOIURL [本文引用: 2]
Enantiornithines are the most speciose avian clade in the Mesozoic, with a fossil record that nearly spans the Cretaceous; however, with less than half of known taxa preserving skull material, our understanding of their cranial morphology remains incomplete. Here we present a comprehensive overview of the current knowledge of enantiornithine skull anatomy and discuss the range of morphologies known for each of the main cranial elements. The typical enantiornithine skull retains numerous ancestral features such as the absence of fusion among bones, the presence of a postorbital bone, a primitive quadrate with a single headed otic process, an unforked dentary, and teeth. The postorbital in at least one taxon is unreduced, suggesting the existence of a complete infratemporal fenestra and thus an unmodified diapsid skull as in confuciusornithids. The rostrum is well known and shows considerable variation, typical of theropods; however, in terms of rostral proportions, enantiornithines are extremely limited within the modern avian spectrum. Although Late Cretaceous skull material is extremely fragmentary, when compared to Early Cretaceous material it reveals a trend towards more specialized morphologies in younger taxa. The foramen magnum in all taxa points caudally, indicating that the 'flexed' type skull morphology may not have evolved in this group. Enantiornithine teeth show considerable diversity in numbers, size, morphology and placement, ranging from taxa with large teeth found throughout the jaws to taxa with small, rostrally restricted teeth, to the fully edentulous. Despite limited preservation of skull material, a number of trophic specializations can be deduced from the range of preserved morphologies, further hinting at the morphological and ecological diversity of the Cretaceous Enantiornithes.
[本文引用: 1]
DOIURL [本文引用: 2]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 4]
DOIURL [本文引用: 5]
[本文引用: 1]
DOIPMID [本文引用: 2]
Enantiornithes are the most successful clade of Mesozoic birds. Here, we describe a new enantiornithine bird, Cruralispennia multidonta gen. et sp. nov., from the Protopteryx-horizon of the Early Cretaceous Huajiying Formation of China. Despite being among the oldest known enantiornithines, Cruralispennia displays derived morphologies that are unexpected at such an early stage in the evolution of this clade. A plough-shaped pygostyle, like that of the Ornithuromorpha, evolved convergently in the Cruralispennia lineage, highlighting the homoplastic nature of early avian evolution. The extremely slender coracoid morphology was previously unknown among Early Cretaceous enantiornithines but is common in Late Cretaceous taxa, indicating that by 131 million years ago this clade had already experienced considerable morphological differentiation. Cruralispennia preserves unusual crural feathers that are proximally wire-like with filamentous distal tips, a new morphotype previously unknown among fossil or modern feathers, further increasing the known diversity of primitive feather morphologies.
DOIURL [本文引用: 2]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 14]
DOIURL [本文引用: 1]
DOIURL [本文引用: 9]
DOIURL [本文引用: 2]
DOIURL [本文引用: 1]
The two groups of archosaurs, crocodilians and birds, form an extant phylogenetic bracket for understanding the reproductive behaviour of dinosaurs. This behaviour is inferred from preserved nests and eggs, and even gravid individuals(1). Data indicate that many 'avian' traits were already present in Paraves-the clade that includes birds and their close relatives-and that the early evolution of the modern avian form of reproduction was already well on its way(2,3). Like living neornithine birds, non-avian maniraptorans had daily oviposition and asymmetrical eggs with complex shell microstructure, and were known to protect their clutches(4-6). However, like crocodilians, non-avian maniraptorans had two active oviducts (one present in living birds), relatively smaller eggs, and may not have turned their eggs in the way that living birds do(1,6). Here we report on the first discovery of fossilized mature or nearly mature ovarian follicles, revealing a previously undocumented stage in dinosaur reproduction: reproductively active females near ovulation. Preserved in a specimen of the long bony-tailed Jeholornis and two enantiornithine birds from the Early Cretaceous period lacustrine Jehol Biota in northeastern China, these discoveries indicate that basal birds only had one functional ovary, but retained primitive morphologies as a result of their lower metabolic rate relative to living birds. They also indicate that basal birds reached sexual maturity before skeletal maturity, as in crocodiles and paravian dinosaurs. Differences in follicular morphology between Jeholornis and the enantiornithines are interpreted as forming an evolutionary gradient from the reproductive condition in paravian dinosaurs towards neornithine birds. Furthermore, differences between the two enantiornithines indicate that this lineage might also have evolved advanced reproductive traits in parallel to the neornithine lineage.
DOIURL [本文引用: 8]
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
DOIURL [本文引用: 2]
DOIURL [本文引用: 2]
[本文引用: 3]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}