 ,1,2, 张百平,1,2, 王晶1,2, 余付勤1,2, 赵超3, 姚永慧1
,1,2, 张百平,1,2, 王晶1,2, 余付勤1,2, 赵超3, 姚永慧1North-south transect series in vegetation of eastern China's north-south transitional zone and their significance for determining climate dividing line
ZHANG Xinghang,1,2, ZHANG Baiping,1,2, WANG Jing1,2, YU Fuqin1,2, ZHAO Chao3, YAO Yonghui1通讯作者:
收稿日期:2019-12-9修回日期:2020-12-4网络出版日期:2021-01-25
| 基金资助: |
Received:2019-12-9Revised:2020-12-4Online:2021-01-25
| Fund supported: |
作者简介 About authors
张兴航(1992-), 女, 河北沙河人, 博士生, 主要从事山地GIS、山地生态环境研究。E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (4493KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
张兴航, 张百平, 王晶, 余付勤, 赵超, 姚永慧. 中国南北过渡带东段样带植被序列与气候分界问题. 地理学报[J], 2021, 76(1): 30-43 doi:10.11821/dlxb202101003
ZHANG Xinghang, ZHANG Baiping, WANG Jing, YU Fuqin, ZHAO Chao, YAO Yonghui.
1 引言
以秦岭—大巴山(以下简称“秦巴山地”)为主体的中国南北过渡带,具有多维地带性结构,是中国南北地理和气候的天然界限,表现出过渡性、复杂性、多样性[1]。植被作为反映自然地理环境的“镜子”,在衡量山地环境结构中发挥重要作用[2,3]。探究秦巴山地植物种群、群落优势成分和物种多样性,能够深化对中国南北过渡带植被变化序列的认识。同时对识别气候南北分界线位置、认识生物多样性和生态安全具有重要意义[2, 4-5]。秦巴山地由秦岭与大巴山组成,秦岭通过高大山体,阻碍南部暖湿气流,也阻隔向南侵入的干冷气流,造成秦岭两侧具有不同气候类型。此处植物群落复杂多样,在南北向上交叉错杂呈现过渡性[1, 6-7]。目前认为秦岭是南北方气候的分界,是常绿阔叶树生存的北端,公认的中国南北方分界线。对于界线具体位置,大体有3种观点:秦岭主脊、秦岭北麓、秦岭南麓,也有****认为大巴山更适合作为气候南北方分界线[8,9,10]。从植被垂直带谱结构上看[9],常绿阔叶林基带的北端在大巴山南坡,表明从秦岭北坡到大巴山南坡完成暖温带到亚热带完整过渡。作为中国南北过渡带主体,秦巴山地是一个完整的地理、地貌单元,将视野从秦岭扩展到整个秦巴山地,会更清晰认识过渡带内变化序列和南北分界位置。因而在东大巴山(eastern Daba Mountains, EBM)、东秦岭(eastern Qinling Mountains, EQM)进行自南向北连续调查和研究,对完整和全面认识东秦巴山地(eastern Qinling-Daba Mountains, EQBM)南北分界线非常必要。
不同****通过研究EQBM自然地理现象得到以下规律:在地形方面,EQBM地形表现出一定程度垂直分带;在气候方面,受山体高度影响,气温和降水呈现垂直地带性和过渡性;在植物方面,植物种类在EQBM南北向有混合现象;在土壤方面,表现明显山地土壤特征和南北向过渡特征[11,12,13,14,15,16]。上述规律均反映出EQBM在南北方向上的过渡特性,表明EQBM是研究中国南北过渡带植被序列与气候南北分界问题的天然实验室。
植物种群和物种组成的空间渐变特征揭示植物变化序列和群落内在性质[4, 17-18]。植物物种多度及其性质在地理空间上的变化,能定量反映该区域植物沿纬度过渡模式,即在南北方向的空间分异[18,19,20,21]。除了种群数量递变特征,植物物种组成的过渡特征也是研究植被在南北向变化的重要内容[22,23]。植物物种组成主要包括群落优势成分与物种多样性[24]。群落优势成分一般用物种重要值表示,即通过相对多度、相对显著度以及相对频度三者之和来衡量[4, 25-26],物种多样性一般用丰富度表示。通过种群数量、物种丰富度等可以定量反映植物南北向变化序列[27,28,29]。
目前已有很多****对EQBM植被过渡特征进行研究,大多针对某个小区域开展。叶永忠等[30]对豫南鸡公山植物区系分布进行分析,认为鸡公山是南北植物分布的天然界线之一,但仅与其临近的黄山、神农架地区进行对比,缺乏沿南北向更多地区序列对比;杨春锋等[31]通过调查和分析九宫山自然保护区植被类型和群系,发现该保护区植物组成类型和森林植被类型具有明显的地带性过渡特征,但是没有进一步讨论过渡特征和南北方分界线问题;秦育峰[32]采用样地记录法对伏牛山区内植物区系与物种多样性进行调查分析,发现该区南北植物交汇明显,是秦岭以东植物多样性最高地区,但文章没有描述具体调查方法,仅是对调查获取数据简单统计;叶永忠等[33]、胡新玉等[34]根据多年实地调查以及文献资料收集,发现河南连康山各种植物成分兼容并存,植物区系种类组成丰富,具有南暖温带向北亚热带过渡的性质,但是没有研究其内部过渡序列,缺乏南北过渡细节的变化研究。而开展大区域研究,主要利用遥感数据,对秦巴山地植被类型进行分类[35]、定量化分析秦巴山地山体效应[36]、挖掘秦巴山地多维地带性背景下其高度复杂的物候现象[37]。以往研究主要通过对区域内植物种类、植物区系、物种多样性等进行分析,认为区内植物种类丰富,存在南北植物交汇,具有明显过渡特征[38,39,40,41,42,43,44]。虽然在一定程度上揭示了EQBM植物群落分布情况,但研究范围局限于小尺度局部区域,在南北方向上缺乏连续采样,忽略了山脉南北延绵本身特点对植物群落带来的巨大影响。
基于此,本文以EQBM南北样带为研究对象,利用数量生态学方法分析群落物种组成,从植物种群、群落结构、物种多样性空间分布3个方面,探讨EQBM植被在南北向的变化序列,间接反映气候差异导致植物由北(暖温带)到南(亚热带)性质的过渡变化,尤其是常绿树种,从而揭示气候南北方界线。本文对于促进我国地理格局、自然资源分布的认识有极为重要的意义,并对农业生产、文化旅游发展、生态环境保护等具有指导作用。
2 数据来源与研究方法
2.1 数据源
本文植物群落数据是通过南北样带调查获取。该样带以人员具有可达性、植物代表性强和人为干扰性低为基本原则进行设计:① 可达性通过距离道路远近判断;② 代表性通过遥感影像纹理、距离自然保护区远近判定;③ 干扰性通过地形判断。结合南北连续采样要求,得到初步样带设计方案,该样带能够代表EQBM相对完整的森林地带性植被;按照初步样带设计方案进行预勘查(2017年5月),根据野外预调查的可达性、代表性和干扰性,进行了调整;在实地植物群落采样调查中主要参照以下原则:“群落内部的物种组成、群落结构和生境相对均匀,群落面积足够,使样方四周能够有10~20 m以上的缓冲区,除依赖于特定生境的群落外,一般选择平(台)地或缓坡上相对均一的坡面,避免坡顶、沟谷或复杂地形”等。自南向北沿着巴东县—兴山县—神农架林区—房县—十堰市—勋县—淅川县—西峡县—卢氏县—灵宝市样带方案进行采样。2017年8月到实地调查采集数据。调查样点数据分布在东秦巴山地(31.0°N~34.5°N, 110.3°E~111.3°E)、受人类活动影响相对较小(海拔范围210~1840 m)的典型森林群落,采样间隔设置为南北5~8 km,大小为20 m×20 m,共获得71个植物群落样点数据,构成EQBM南北方向上一条样带(图1)。数据记录森林样方内胸径大于2 cm的所有乔、灌木物种,调查指标包括:乔木层及灌木层的物种种类、个体数量、树高、冠幅、胸径(基径)、盖度、样地经纬度、海拔高度、坡度、坡向、坡位。共包含315个物种类型,其中乔木物种有172种。地形数据来自美国地质调查局1∶25万的数字高程模型(Digital Elevation Model, DEM)。
图1
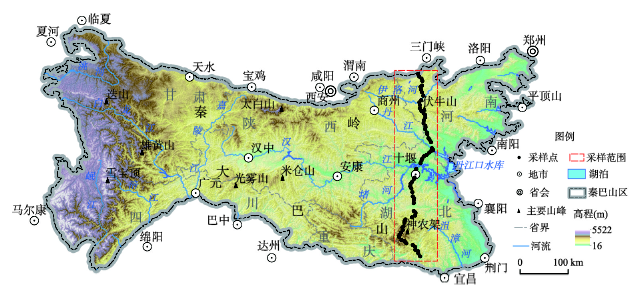 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1中国南北过渡带主体区域及采样点分布
Fig. 1The main body of the north-south transitional zone of China and distribution of sampling sites
2.2 分析方法
采用主成分法分析东秦巴山地(EQBM)南北样带植物种群在南北向的变化规律。通过降维技术把一组存在相关性的变量转换为少数几个不相关的变量,转换后取几个主要的变量为主成分。主成分分析是对多维变量之间相关性进行分离的一种统计方法,利用少数几个主要成分来揭示多维变量对目标影响关系,在尽可能多地保留原始变量信息情况下,压缩数据维度,减少数据量。运用相异系数法探讨东秦巴山地南北样点之间物种的空间变异性,进行群落结构划分。在物种数据分析方面,常用系数有Bray-Curtis相异系数、弦距离和Hellinger距离。这些系数既要体现每个样本里物种存在与否,还要体现每个物种相对丰度。其中弦距离实际是指先将样方向量的范数标准化后(平方和为1)再计算样方对的欧氏距离;Hellinger距离是多度值先除以样方多度总和再取平方根后计算的欧氏距离;Bray-Curtis相异矩阵(式(1))可以直接由原始数据计算,依据多度绝对数量差,在实际中运用较多。因此本次研究选择Bray-Curtis相异矩阵方法计算相异系数。
式中:d(i, j)是样点i和样点j物种的相异系数;k是物种数(样地—物种矩阵中物种数);y表示物种多度;i和j表示待比较的2个样地编号。
Ward最小方差聚类法主要包括:① 选定相异性指标,计算不同样方之间相异性,本次采用Bray-Curtis相异系数作为评价样方之间相异性指标;② 将差异最小的样方合并成新样方组;③ 计算新样方组相异性;④ 重复②、③步骤,直至全部样方聚为一类停止。
重要值主要是衡量物种在群落中重要程度,通过测度指标推算获取,不是简单测量得到。计算公式是(式(5))[4]用相对多度(式(2))、相对频度(式(3))和相对优势度(式(4))三者之和来表示。但在森林植被群落里,通常采用乔木层的相对优势度(相对胸高断面积)表示重要值(式(6))[45]。因此采用乔木物种相对胸高断面积表示重要值。
本文按照植物种群、群落结构、物种多样性3个方面系统展示EQBM植被在南北方向变化序列,揭示EQBM的过渡性和丰富性,并为气候南北方分界线的判断提供新思路。
3 结果分析
3.1 典型乔木种群在南北样带的变化序列
图2
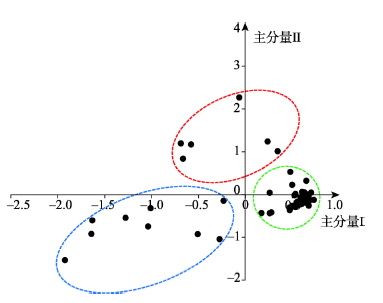 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2东秦巴山地南北样带上植物种类排序
Fig. 2Sequence of plant species in the north-south transect of eastern Qinling-Daba Mountains
本文利用调查的71个样方数据,筛选各样方乔木物种,根据乔木物种在样方中的多度指标(出现的样方数≥ 8),基于主成分法对典型乔木植物排序,计算结果中前两个主分量信息量最大,分别占总信息量的35.66%和23.84%,能够代表植物种群的区域分异特点,因此由第一、二主分量构成排序平面图(图2)。结果表明,EQBM乔木物种在排序图中形成相对集中3组。
为了更清晰显示排序图2中3组植物物种的空间分布规律,对组1、组2的乔木物种以及组3中典型的乔木物种多度沿南北向变化做气泡地图(图3)。
图3
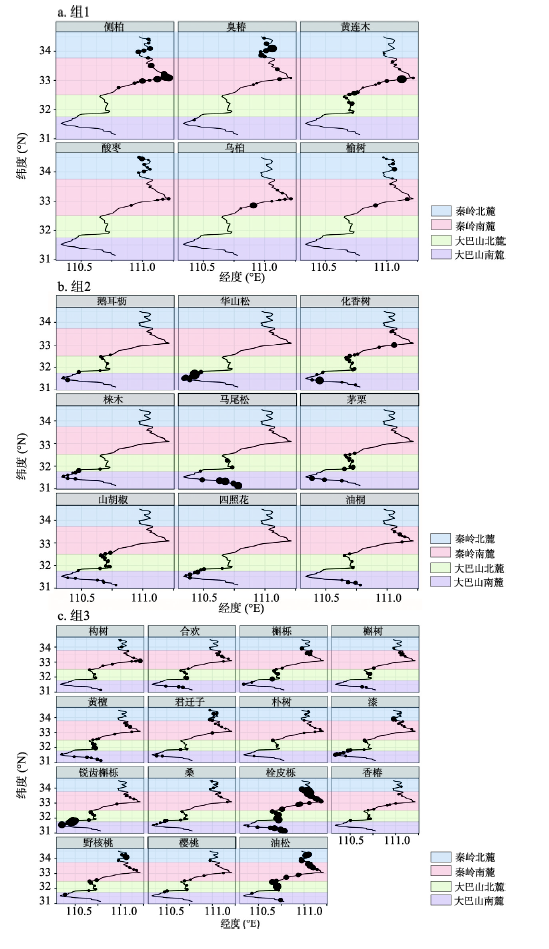 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3物种多度沿南北方向分布的气泡地图
注:图中的气泡大小与当前物种多度成正比。
Fig. 3Bubble map of species abundance along the north-south direction
这些排序图中不同组植物,大致以东秦岭南麓(south flank of eastern Qinling Mountains, EQMS)为界表现出不同空间分布规律(图3),并对各组植物性质进行统计分析(表1),结果表明:① 组1中植物主要分布在EQMS以北(32.5°N~33.0°N以北)(图3a),为北方旱性较强的一组植物(表1),以下均称为北方组植物,如侧柏。② 组2中植物主要分布在EQBM南段(32.5°N~33.0°N以南)(图3b)。植物性质属于南方喜湿喜热的植物成分(表1),以下均称为南方组植物,如马尾松等。③ 组3中植物在东秦巴山地南北均有分布(图3c),以下均称为广布种,说明该组植物对本地区生态环境具有较强的适应能力。
Tab. 1
表1
表1排序结果中各组乔木植物性质构成
Tab. 1
| 组别 | 过渡种占比(%) | 广布种占比(%) | 集中分布地段 | |
|---|---|---|---|---|
| 北方类型植物 | 南方类型植物 | |||
| 1 | 83 | 17 | 0 | 北段 |
| 2 | 0 | 78 | 22 | 南段 |
| 3 | 13 | 0 | 87 | 北、南段 |
新窗口打开|下载CSV
根据图2中3组乔木物种在南北分布的物种数统计(表2),发现见于EQBM北部的乔木物种有25种,其中仅在北部出现的有4种,另外21种均在南部零星出现。而见于EQBM南部的乔木物种有28种,其中仅在南部出现的有10种,另外18种均在北部零星出现。经过百分比计算(表2),得出见于EQBM北部的植物,84%都见于南部。而见于南部的植物,只有64%见于北部。产生这种现象的原因可能是植物从偏冷地区(北部)到偏潮湿温润地区(南部)更加有利。而喜热喜湿的南方植物(如山胡椒、化香树、马尾松等等)则不能越过北部山坡。
Tab. 2
表2
表2植物物种在样线南北部的分布比例情况
Tab. 2
| 物种 | 北部出现 | 南部出现 | |||
|---|---|---|---|---|---|
| 仅在北部出现 | 南部也零星出现或分布 | 仅在南部出现 | 北部也零星出现或分布 | ||
| 物种数量(种) | 4 | 21 | 10 | 18 | |
| 占比(%) | 16 | 84 | 26 | 64 | |
新窗口打开|下载CSV
3.2 乔木群落类型在南北样带的变化序列
3.2.1 南北样带乔木群落类型 根据中国植被分类法,植物群落分类单位一般采用群丛、群系和植被型[46]。层片结构特点和优势物种均一致的群落合并为群丛。优势物种一致的植物群落合并为群系[47]。依照这一分类系统,运用Ward最小方差聚类法,对EQBM南北样带上乔木植物群落进行聚类分析。在R语言中编程[48]得到聚类结果(图4),当欧氏距离d = 1.5时,EQBM南北样带乔木植物群落划分为7个群系;当欧式距离d = 1时,又可分为16个群丛。图4
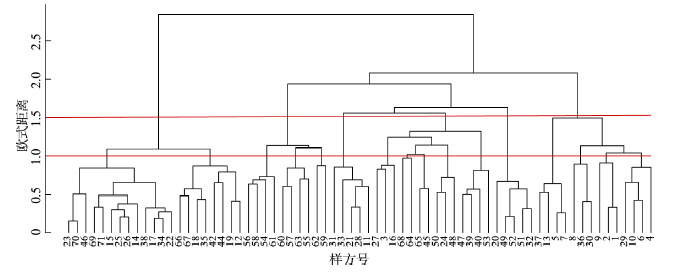 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4东秦巴山地南北样带乔木植物群系Ward聚类图
Fig. 4Ward cluster map of tree flora in north-south transect in the eastern Qinling-Daba Mountains
重要值作为一种综合性指标,可以衡量植物物种在群落中相对重要性。通过计算每组物种重要值来获取该组优势种,按照优势种对这7个群系进行命名,分别为:栓皮栎+马尾松群系、锐齿槲栎+华山松群系、栓皮栎+侧柏群系、油松+化香树群系、栓皮栎+毛黄栌群系、油松+刺槐群系、刺槐+野核桃群系。16个群丛分别为:栓皮栎+马尾松+油松群丛、栓皮栎+马尾松群丛、锐齿槲栎+华山松群丛、锐齿槲栎+华山松+四照花群丛、米心水青冈+湖北海棠群丛、栓皮栎+侧柏+黄连木群丛、侧柏+毛黄栌群丛、化香树+青冈群丛、短柄枹栎+杉木群丛、油松+黄檀群丛、槲栎+山胡椒群丛、栓皮栎+毛黄栌群丛、油松+刺槐群丛、栾树+刺槐群丛、野核桃+臭椿群丛、刺槐+侧柏+构树群丛。
3.2.2 南北样带乔木群落特征 聚类结果中7个群系在EQBM南北样带空间分布结果如图5所示。调查样地主要群系为栓皮栎+马尾松群系、锐齿槲栎+华山松群系和刺槐+野核桃群系。栓皮栎+马尾松群系最北分布至伏牛山,伏牛山以北主要为刺槐+野核桃群系,锐齿槲栎+华山松群系仅分布于南北样带南部(EBMS)。各群系具体分布特征为:① 栓皮栎+马尾松群系:最北分布至伏牛山,物种丰富(74种乔木物种),优势种为栓皮栎和马尾松。另外,在东秦岭段(eastern Qinling Mountains, EQM)是以栓皮栎为优势的群系,而东大巴山段(eastern Daba Mountains, EBM)则是以马尾松为优势的群系,下文,该群系在EQM中表示为栓皮栎+马尾松,EBM中表示为马尾松+栓皮栎;② 锐齿槲栎+华山松群系:该群系仅分布于南北样带南段(31.3°N~31.5°N范围),即东大巴山南麓(south flank of eastern Daba Mountains, EBMS),分布集中,物种丰富(92种乔木物种);③ 栓皮栎+侧柏群系:该群系分布零星,仅见于样带的北段,即EQM一带;④ 油松+化香树群系:分布于东秦岭南麓(south flank of eastern Qinling Mountains, EQMS)和EBM;⑤ 栓皮栎+毛黄栌群系:该群系分布在锐齿槲栎+华山松群系的北部,与之相邻,主要在EQMS和东大巴山北麓(north flank of eastern Daba Mountains, EBMN);⑥ 油松+刺槐群系:主要分布于EQM,乔木物种简单;⑦ 刺槐+野核桃群系:主要分布在东秦岭北麓(north flank of eastern Qinling Mountains, EQMN),相对集中,群落物种丰富(34种乔木物种)。
图5
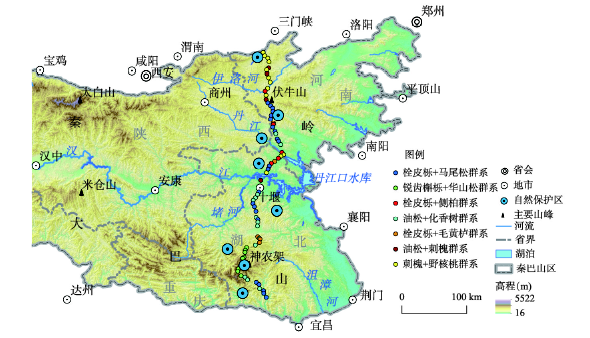 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5东秦巴山地南北样带上群系空间分布
Fig. 5Spatial distribution of formation on the north-south transect of the eastern Qinling-Daba Mountains
3.3 乔木物种多样性在南北样带的变化序列
本文选取物种丰富度量化植物物种多样性,分析其在南北向的变化特征。丰富度即每个样方中物种数,样方物种数越多,丰富度越大。对样方中乔木植物科、属、种在南北向的变化进行分析,结果表明(图6),科、属、种3个分类群表现近似变化特征:兴山县(31.5°N以南)地段热量充足,但其物种多样性明显偏低,由于海拔低,可能受人为干扰影响。向北至神农架林区,海拔升高,气候转湿,多样性显著增长,神农架北部(海拔约1800 m,31.5°N~32.0°N间)迅速达到峰值;然后随海拔下降,进入房县(32.0°N附近),多样性急剧下降达到谷底。然而,从房县北部(32.3°N附近)向北又出现一个物种多样性上升—下降变化模式直至EQBM分界线(32.8°N附近);再向北,即EQM,多样性呈平缓降低,在伏牛山(34.0°N附近)出现一个峰值,为EQM物种多样性最高值。图6中可以看出EQM科、属、种多样性相对EBM低,且保持相对平稳变化趋势。图6
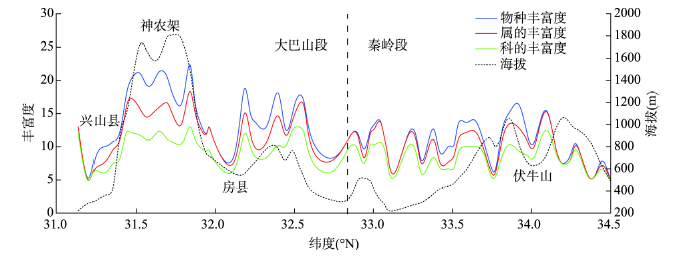 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6东秦巴山地南北样带物种丰富度的纬度梯度变化
Fig. 6Latitudinal gradient changes of species richness in the north-south transect of the eastern Qinling-Daba Mountains
4 讨论与结论
4.1 讨论
4.1.1 典型乔木种群和物种多样性的区域分异 EQBM南北样带上典型乔木种群和物种多样性具有南北分异特点,反映该地区生态环境条件复杂。从典型乔木种群南北变化序列特征看,EQBM北部地段植物北方旱生性强,主要受干冷气流影响大。向南受干冷气流影响小,生态环境相对温暖湿润,植物种群比北段复杂多样。最南段干冷空气基本消失,亚热带气候显著,气候适宜,植物种群最丰富[49]。
从乔木物种多样性南北变化序列特征看,物种丰富度随海拔增高和地理位置北移而不同程度地降低,物种多样性最高处在EBM段1800 m左右,物种多样性最低处在EQM段200~400 m。
4.1.2 乔木群落的空间分布特征 EQBM南北样带上乔木群落分为7个群系,其中栓皮栎+马尾松群系、油松+化香树群系、油松+刺槐群系和栓皮栎+侧柏群系为4个主要群落类型;其次,栓皮栎+毛黄栌群系、刺槐+野核桃群系和锐齿槲栎+华山松群系只分布在局部范围内。聚类结果主要反映群落生存的生态环境相似性,生境相似性最大的群落最先合并。油松+化香树群系分布于南北样带中部和南部,该群系属于南方植物,喜热喜湿。而栓皮栎+侧柏群系在南北样带零星分布,主要见于样带北部,群落属于北方旱性植物。栓皮栎+马尾松群系分布范围较广,群落物种复杂多样,聚类中最后合并,表明与其他群系生态环境有显著差异。
从该地区南北样带乔木群落结构可知,由于人类活动影响,该地区乔木植物群落受到较大破坏,如栓皮栎+侧柏群系等零星分布的小群落,可能与人类活动导致群落生境破碎有关。
4.1.3 东秦巴山地南北方分界线 本次研究结果说明,东秦岭南麓(EQMS)更适宜作为东秦巴山地(EQBM)南北分界线。首先,从典型乔木种群在南北样带变化序列得出,EQMN和EQMS以北方旱性植物为主,但EQMS出现了南方性质的常绿树种,也是南方和北方植物混合最明显地带,EBMN和EBMS以南方湿润性植物为主,基本没有出现北方旱性植物。这表明EQMS比其他地区植物种类混合强烈,更适合作为EQBM气候南北分界线地区。
其次,从乔木群落类型在南北样带的变化序列得出,EQMN群系4个(北方性质群系3个,南方性质群系0个,广布性质群系1个):栓皮栎+马尾松群系、刺槐+野核桃群系、栓皮栎+侧柏群系、油松+刺槐群系,主要为北方植物;EQMS群系6个(北方群系3个,南方群系1个,广布群系2个):栓皮栎+侧柏群系、刺槐+野核桃群系、栓皮栎+毛黄栌群系、油松+刺槐群系、栓皮栎+马尾松群系、油松+化香树群系,一部分群系和EQMN一致,另一部分群系偏南方植物性质,即开始发生南北过渡现象; EBMN群系4个(北方性质群系0个,南方性质群系2个,广布性质群系2个):栓皮栎+毛黄栌群系、油松+刺槐群系、马尾松+栓皮栎群系、油松+化香树群系,与EQMS南方植物性质的群系一致;EBMS群系3个(北方性质群系0个,南方性质群系3个,广布性质群系0个):油松+化香树群系、马尾松+栓皮栎群系、锐齿槲栎+华山松群系,前两个群系与EBMN群系中一致,而锐齿槲栎+华山松群系只在EBMS存在。只有EQMS群系出现南北性质的混合。因此群落结构层面,东秦岭南麓(EQMS)作为东秦巴山地(EQBM)气候南北方分界线值得考虑。
再之,科、属、种3个分类群在EQM上多样性的变化趋势相对平稳,EBM整体高于EQM。进一步印证EBM气候条件比EQM适合更多植物生存(与3.1小节中结果可以交叉验证),使得在确定EQBM气候南北分界线时采用南方植物初次出现的位置(即开始出现常绿树种[50]),而不是北方植物消失的位置。为了更直接反映东秦巴山地(EQBM)气候南北分界线的位置,本文将样方中植物分为南方和北方物种两类,分别计算每个样方中南方物种数比例和北方物种数比例(图7)。图7中可以看出,北方物种比例在EQMN最高,向南到EQMS逐渐减少,到EBM一带逐渐消失;而南方物种比例在EBMS最高,向北到EBMN略有减少,再向北到EQMS基本减少至零,在EQMN基本消失。由此可见,在东秦岭南麓(EQMS)南北方植物混杂现象最明显,东秦岭南麓(EQMS)更适合作为东秦巴山地(EQBM)气候南北方分界线。
图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7东秦巴山地南北样带不同类型物种丰富度的纬度梯度变化
Fig. 7Latitudinal gradient changes of species richness of different types in the north-south transect of eastern Qinling-Daba Mountains
最后,根据样线中物种丰富度最高的两个区域(宝天曼和神农架)野外实地调查数据,以及收集到EQMN小秦岭、EQMS新开岭、化龙山自然保护区、EBMN五道峡、十八里长峡自然保护区、EBMS五里坡共8个自然保护区(图5所示)的数据,EQMN到EQMS年均降雨量800 mm左右,与北方性质植物需求的水分条件一致。EQMS至EBMS,年均降水量均达到1100 mm以上,满足南方性质植物存在的水分需求。因此从干湿状况看东秦岭南麓(EQMS)作为东秦巴山地(EQBM)气候南北方分界线是合理的。气候类型和山地植被基带类型的分界在EQMS,如表3所示,从EQMN到EQMS,气候类型由暖温带季风变为亚热带暖温带季风气候,山地基带植被类型由落叶阔叶林变为常绿落叶阔叶混交林。这也表明东秦岭南麓(EQMS)更适宜作为东秦巴山地(EQBM)气候南北方分界线。
Tab. 3
表3
表3东秦巴山地南北样线上部分自然保护区的基础数据
Tab. 3
| 自然保护区 | 位置 | 纬度 (N) | 经度 (E) | 年均降水(mm) | 气候类型 | 山地植被基带类型 |
|---|---|---|---|---|---|---|
| 小秦岭 | 秦岭东段北坡 | 34°27′ | 110°33′ | 719.2 | 暖温带季风气候 | 低山灌丛草甸农作物带、落叶栎林带 |
| 宝天曼 | 33°30′ | 111°48′ | 885.6 | 暖温带季风气候 | 落叶栎林带 | |
| 新开岭 | 秦岭东段南坡 | 33°16′ | 110°38′ | 803 | 亚热带与暖温带季风气候 | 常绿落叶阔叶混交林 |
| 化龙山 | 33°07′ | 109°20′ | 1015 | 亚热带暖温带过湿山地气候 | 常绿阔叶林 | |
| 五道峡 | 大巴山东段北坡 | 32°5′ | 111°07′ | 1100 | 北亚热带季风气候 | 常绿阔叶林和常绿落叶阔叶混交林 |
| 十八里长峡 | 31°31′ | 109°50′ | 1250 | 北亚热带湿润季风气候 | 常绿落叶阔叶混交林 | |
| 神农架 | 大巴山东段南坡 | 31°36′ | 110°27′ | 1200 | 北亚热带季风气候 | 常绿阔叶林 |
| 五里坡 | 31°22′ | 110°03′ | 1400 | 中亚热带温润季风气候 | 常绿阔叶林和常绿针叶林带 |
新窗口打开|下载CSV
目前南北分界线位置主要是从植被宏观尺度(如植被类型、NDVI等)进行研究,而本文基于东秦巴山地自南向北连续植物群落样带调查数据,以定性和定量分析相结合的方式,从植被小尺度(植物物种、群落)上认识过渡带内变化序列,为气候南北方分界线的科学研究提供新思路。实地采样使数据更精准,定性是将植物分为南方和北方性质植物,利用南北植物混合现象来说明EQBM气候南北分界线,定量是将采样的植物群落数据进行量化,如典型植物种群多度、群系个数、物种丰富度。
本文认为东秦岭南麓(EQMS)更适合作为东秦巴山地(EQBM)南北方分界线的结论与张学忠等[51]、康慕谊等[16]看法相同。而与赵芳等[9]研究结论存在差异,其主要原因包括:研究尺度不同,本文是针对植物群落和种群尺度(常绿树种),而其是植被类型尺度(常绿阔叶林);研究角度不同,本文侧重水平向序列变化研究,而其侧重山地垂直向植被变化规律;采用的数据分布方式不同,本文是连续的水平样带调查数据,而其是离散的山地垂直带谱数据。目前,笔者正在收集秦巴山地自然保护区植物科、属、种数据,未来研究秦巴山地南北植物区系(特别是气候变化背景下)垂直向的响应及其量化。
4.2 结论
本文基于东秦巴山地南北样带植物群落实地调查数据,分析植物种群、群落结构、物种多样性在南北向的变化序列,得到以下结论:(1)植物种群具有南北分异特点,分布在EQMN的植物均属旱性特点的北方植物,EQMS开始出现常绿树种,分布在EBM的植物主要为喜湿喜热南方植物。
(2)运用Ward最小方差聚类法对东秦巴山地南北样带乔木群落聚类为7个群系和16个群丛。EQMN群系4个(北方群系3个,南方群系0个,广布群系1个)、EQMS群系6个(北方群系3个,南方群系1个,广布群系2个)、EBMN群系4个(北方群系0个,南方群系2个,广布群系2个)、EBMS群系3个(北方群系0个,南方群系3个,广布群系0个),只有东秦岭南麓(EQMS)群系出现南北性质的混合。
(3)科、属、种3个分类群的丰富度变化特征近似:由南向北,物种多样性呈减小趋势,EQM科、属、种多样性相对EBM低,变化趋势平稳,但南北方植物混合现象开始在东秦岭南麓(EQMS)出现。
本文关于过渡带内植被变化序列的结论增加了对东秦巴山地气候南北分界线判断的科学性,一致表明东秦岭南麓(EQMS)更适合作为东秦巴山地(EQBM)气候南北分界线。研究结果可为东秦巴山地植被保护、规划和管理提供基础参考,对国家保护地体系建立、国家生态环境保护和区域可持续发展具有重大意义,同时为中国南北过渡带研究的进步或突破提供坚实的科学数据。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 4]
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1073/pnas.1308932111URLPMID:24847062 [本文引用: 1]

Plant diversity, like that of most other taxonomic groups, peaks in the tropics, where climatic conditions are warm and wet, and it declines toward the temperate and polar zones as conditions become colder and drier, with more seasonally variable temperatures. Climate and evolutionary history are often considered competing explanations for the latitudinal gradient, but they are linked by the evolutionarily conserved environmental adaptations of species and the history of Earth's climate system. The tropical conservatism hypothesis (TCH) invokes niche conservatism, climatic limitations on establishment and survival, and paleoclimatic history to explain the latitudinal diversity gradient. Here, we use latitudinal distributions for over 12,500 woody angiosperm species, a fossil-calibrated supertree, and null modeling to test predictions of the TCH. Regional assemblages in the northern and southern temperate zones are less phylogenetically diverse than expected based on their species richness, because temperate taxa are clustered into relatively few clades. Moreover, lineages with temperate affinities are generally younger and nested within older, more tropical lineages. As predicted by the TCH, the vast majority of temperate lineages have arisen since global cooling began at the Eocene-Oligocene boundary (34 Mya). By linking physiological tolerances of species to evolutionary and biogeographic processes, phylogenetic niche conservatism may provide a theoretical framework for a generalized explanation for Earth's predominant pattern of biodiversity.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s11442-020-1727-6URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}