 , 高乃征2, 张贵驰1, 王建1
, 高乃征2, 张贵驰1, 王建1 , 凌婉婷1
, 凌婉婷11. 南京农业大学土壤有机污染物控制与修复研究所, 南京 210095;
2. 山东省沂南县第三中学, 临沂 276300
收稿日期: 2020-06-16; 修回日期: 2020-08-06; 录用日期: 2020-08-06
基金项目: 国家自然科学基金(No.41771523);中央高校基本科研业务费专项资金(No.KJQN202120)
作者简介: 段振宇(1996—), 女, E-mail: 2018103034@njau.edu.cn
通讯作者(责任作者): 王建, E-mail: wj308119@njau.edu.cn
摘要:蔬菜多环芳烃(PAHs)污染威胁人群健康.通过水培试验,研究了接种具有菲降解功能的植物内生细菌Diaphorobacter sp. Phe15对空心菜及其亚细胞组分中菲积累的影响,揭示了空心菜体内多酚氧化酶(PPO)和过氧化物酶(POD)的响应及其与PAHs积累的相关关系.Phe15可定殖在空心菜体内,不同部位菌株Phe15细胞数量表现为根表 > 根内 > 茎叶.接种菌株Phe15有效地降低空心菜及其亚细胞组分中菲含量,根部最高降低93.69%,叶部最高降50.26%,亚细胞水平上细胞液中降低最多.接种菌株Phe15提高了空心菜不同部位及其亚细胞组分中POD和PPO活性,且酶活性与菲含量之间呈负相关关系,表明这两种酶活性的提高可降低空心菜体内菲积累.研究结果从亚细胞层面揭示了功能内生细菌削减植物体内PAHs积累的作用规律及机制,为利用功能内生细菌削减作物PAHs污染风险、保障污染区农产品安全等提供了理论依据.
关键词:植物内生细菌多环芳烃空心菜亚细胞酶
Inoculation of endophytic bacteria for the abatement of the phenanthrene accumulation in vegetable subcellular fraction: Efficiency and mechanism
DUAN Zhenyu1
, GAO Naizheng2, ZHANG Guichi1, WANG Jian1, LING Wanting11. Institute of Organic Contaminant Control and Soil Remediation, Nanjing 210095;
2. The Third Middle School of Yinan, Linyi 276300
Received 16 June 2020; received in revised from 6 August 2020; accepted 6 August 2020
Abstract: In this study, the changes in phenanthrene contents and enzymatic activities (such as polyphenol oxidase (PPO) and peroxidase (POD)) in water spinach and its subcellular fractions were comprehensively investigated by inoculating a phenanthrene-degrading bacterium Diaphorobacter sp. Phe15, using hydroculture pot experiments. Results showed that strain Phe15 could effectively be colonized into water spinach through root immersion, and the cell counts of Phe15 in each part of water spinach were: root surfaces > inner roots > shoots. Inoculation led to a phenanthrene content reduction of 93.69% and 50.26% in roots and shoots, respectively, resulting in a decreased concentration in the subcellular fractions of water spinach. The phenanthrene content decreased the most in cell soluble fractions. Also, the enzymatic activities of POD and PPO in roots, shoots, and subcellular fractions of water spinach increased with the inoculation of Phe15. Notably, the enzymatic activities of POD and PPO negatively correlated with the contents of phenanthrene in water spinach, indicating that the increased activities of these enzymes would lead to a decrease in phenanthrene content in plants. The findings in this study reveals the underlying mechanisms of endophytic PAH degradation in plants, from the perspective of subcellular fractions and enzyme activity. This study will provide the necessary theoretical basis for using bacterial endophytes in reducing the risks while ensuring agricultural crop safety in PAH-contaminated fields.
Keywords: endophytic bacteriaPAHswater spinachsubcellular fractionPAH-degrading enzymes
1 引言(Introduction)PAHs是一类具有“三致”效应的有毒有机污染物, 易在土壤中累积.2014年环境保护部和国土资源部联合发布的《全国土壤污染状况调查公报》中指出, “我国土壤中PAHs点位超标率高达1.4%”.PAHs已成为我国土壤环境中常见的一类有机污染物(Ma et al., 2014; Wang et al., 2015; Yu et al., 2019).进入土壤中的PAHs可在土壤-作物系统中迁移并在作物体内积累, 导致作物PAHs污染, 最终通过食物链进入人体进而威胁人类健康.Tao等(2006)针对津、鲁地区的调查结果表明, 谷物中苯并[a]芘含量高达13.2 μg·kg-1, 超出国家标准2.6倍.Wang等(2017)对长三角典型PAHs污染区调查结果表明, 蔬菜和水稻中PAHs对当地居民存在潜在的致癌风险.显然, 作物PAHs污染问题已严重影响农产品安全和人群健康.因此, 如何有效地削减作物中PAHs积累, 保障农产品生产安全和人群健康已成为国内外环境领域研究的热点之一.
功能内生细菌是指从污染区健康植物体分离筛选获得的、具有有毒有机物降解能力、并能够重新定殖在植物体内的一类细菌.重新定殖回植物体内后, 功能内生细菌能在增强宿主植物抗性、减低植物体内有毒有机物积累等方面起着重要作用(Germaine et al., 2004; Afzal et al., 2014; Brader et al., 2014).近年来, 利用功能内生细菌削减植物体内PAHs污染风险的研究引起国内外****广泛关注.Liu等(2017)研究了接种Massilia sp. Pn2对小麦积累菲的影响, 结果发现, 菌株Pn2能够良好定殖在小麦体内, 并促进小麦体内菲的降解.Sun等(2014)从看麦娘中分离到一株具有芘降解能力的内生细菌BJ06, 发现接种菌株BJ06能够重新定殖在黑麦草体内并降低植物体内菲的积累.Zhu等(2017)研究发现, 定殖功能内生细菌Serratia sp. PW7显著降低小麦体内芘的积累.然而, 以往研究大部分是关注功能内生细菌对植物体内PAHs积累的影响, 关于功能内生细菌对不同亚细胞组分中PAHs积累的影响却鲜有报道.
众多研究表明, 一系列酶系在植物体内PAHs代谢过程中起着关键作用.过氧化物酶(POD)和多酚氧化酶(PPO)是有机污染物代谢过程中备受关注的两种酶.POD可以清除生物体内的活性氧自由基, 保护机体免受毒害(Yan et al., 1997).PPO能催化PAHs开环, 生成易降解的中间产物, 在PAHs降解过程中起关键作用(Iwabuchi et al., 1998).研究表明, 接种功能内生细菌可调控植物体内PAHs代谢相关酶系活性, 进而影响植物体内PAHs代谢.相妍冰(2017)通过定殖功能内生细菌Pn2使小麦POD、PPO的活性增强, 促进小麦体内菲的降解, 进而缓解菲对小麦生长的胁迫作用.研究指出, 植物亚细胞组分是有机污染物解毒的关键部位(Sandermann, 1994).然而, 功能内生细菌作用下植物亚细胞组分中PAHs代谢相关酶系活性的响应及其与PAHs积累的相关关系仍不明确, 有待进一步探讨.
基于以上, 本研究选择根系发达、容易培养、生长迅速、江苏省日常食用的空心菜作为代表性蔬菜, 以环境中分布较为广泛的菲为代表性PAHs, 利用实验室前期分离的一株具有菲降解能力的功能内生细菌Diaphorobacter sp. Phe15, 采用水培试验, 研究接种功能内生细菌对蔬菜亚细胞水平菲积累的削减作用, 分析功能内生细菌作用下亚细胞组分中PAHs代谢相关酶系活性的响应及其与PAHs积累的关系.研究结果有望从亚细胞层面阐释接种功能内生细菌影响作物体内PAHs积累的潜在机制, 以期为利用功能内生细菌减低污染区植物体内PAHs积累, 削减作物PAHs污染的人群健康风险, 保障污染区农产品生产安全和人群健康等提供理论依据.
2 材料与方法(Materials and methods)2.1 供试试剂所需试剂:菲购自Aldrich公司, 纯度均大于等于98%.聚乙烯吡咯烷酮(PVP)、邻苯二酚、愈创木酚、过氧化氢、磷酸盐缓冲液(PBS缓冲液)、醋酸缓冲液、亚细胞提取液、霍格兰营养液(HNS)、LB培养基和无机盐培养基(MSM).二氯甲烷、正己烷为分析纯, 甲醇为色谱纯.
2.2 供试植物与菌株供试植物:空心菜(Ipomoea aquatica Forsk.).
供试菌株:使用本实验室前期分离筛选的一株具有菲降解能力的Diaphorobacter sp. Phe15(顾玉骏, 2015)作为供试菌株.Phe15为革兰氏阴性菌, 菌体形状为杆状, 单端生鞭毛, 好氧(图 1a).将菌株Phe15的16S rRNA基因序列在NCBI上进行比对分析, 发现与Diaphorobacter sp.有很高的同源性(99.6%), 结合菌株的生理生化特性, 鉴定菌株Phe15属于Diaphorobacter sp.(图 1b).
图 1(Fig. 1)
 |
| 图 1 菌株Phe15的透射电镜图(a)及基于16S rRNA基因序列同源性的菌株Phe15系统发育树(b) Fig. 1Transmission electron micrographs of strains Phe15 (a) and the Phylogenetic tree of strains Phe15 based on the 16S rRNA gene homology (b) |
2.3 水培试验采用水培试验方法, 研究接种功能菌株对植物体内PAHs含量以及PPO和POD活性的影响.将空心菜种子用75%乙醇浸泡5 min后, 用无菌水清洗3次, 随后置于育苗盘中育苗.待空心菜幼苗长至约10 cm时备用.将菌株Phe15接种于含有氨苄青霉素(Amp)的液体LB培养基中, 于30 ℃, 150 r·min-1振荡培养, 待生长到对数生长期时, 6000 r·min-1离心10 min, 用MSM洗涤2次, 随后将它在600 nm处的吸光度调整为1.0, 备用.然后将备好的空心菜幼苗根部置于菌液中, 浸根8 h.对照处理浸根于等体积无菌水中.随后将空心菜苗移栽入含2 mg·L-1菲的HNS中.植物生长过程中根据具体情况适当补充HNS, 保持液面高度, 并定期随机交换培养瓶位置.于第2、5、8、12 d整瓶采样.实验共设两个处理:HNS+空心菜(CP), HNS+空心菜+功能菌(CPR), 每个处理有3个平行.
2.4 植物亚细胞组分提取采用Kang等(2010)方法分离空心菜亚细胞组分, 略有改动.将0.5 g新鲜空心菜样品置于5 mL亚细胞提取液中, 充分混合后研磨, 过100 μm尼龙细胞筛去除纤维, 收集过滤液于离心管中, 500 g离心5 min获得细胞壁组分(Cell wall, CW);将上清液转移至新的离心管内, 10000 g离心30 min获得细胞器组分(Cell organelle, CO);再次重复上一步, 将上清液置于高速离心机中10000 g离心30 min获得细胞膜组分(Cell membrane, CM), 上清液即为细胞液组分(Cell soluble, CS).
2.5 植物、水、亚细胞PAHs含量分析① 植物样PAHs提取:采集的植物样品冷冻干燥4 d后, 充分剪碎.称取一定量植物样品于25 mL玻璃瓶中, 加入10 mL 1∶1的二氯甲烷和正己烷溶液, 超声萃取30 min, 重复3次, 将萃取液过2 g无水硫酸钠和2 g硅胶的层析柱, 再用10 mL体积比为1∶1的二氯甲烷和正己烷溶液洗脱(高彦征等, 2005).将滤液全部转移至旋转蒸发瓶中, 并于40 ℃蒸发浓缩至干, 用2 mL色谱纯甲醇定容, 过0.22 μm有机相滤膜至液相小瓶中, 利用高效液相色谱(HPLC)对PAHs进行定量分析.
② 水样中PAHs提取:取10 mL水样, 加入20 mL色谱纯甲醇, 超声萃取1 h, 随后过0.22 μm有机相滤膜, 利用HPLC检测其中的PAHs含量.
③ 亚细胞组分PAHs提取:将分离得到的亚细胞组分冷冻干燥4 d后, 称取重量.加入5 mL 1∶1的二氯甲烷和正己烷溶液, 超声萃取30 min, 重复3次, 之后参考植物样操作进行提取.
HPLC的分析条件具体参考Chen等(2019).
2.6 植物及亚细胞POD、PPO活性测定参考Shah等(2001)研究方法, 测定空心菜体内及其亚细胞组织中PPO和POD活性, 略有改进.称取植物样品0.1 g, 加入0.05 g聚乙烯吡咯烷酮(PVP)和3 mL pH 7.8磷酸缓冲溶液(PBS), 在冰浴中充分研磨, 于10000 r·min-1离心10 min, 上清液即为植物体粗酶液.参考李腾武等(1999)研究方法, 在分离亚细胞组分的过程中, 每个层次离心得到的沉淀加入3 mL PBS, 于10000 r·min-1离心15 min, 上清液即为细胞壁、细胞器及细胞膜的酶提取液, 细胞液可直接测定酶活.
POD的测定参考郝再彬等(2004)的方法, 略有改动:向玻璃试管中依次加入2 mL 0.1 mol·L-1的醋酸缓冲液(pH=5.0), 1 mL愈创木酚(W/V=0.25%), 0.1 mL H2O2(V/V=3%)和20 μL粗酶液.于37 ℃水浴15 min, 于470 nm波长下测定吸光度.所有操作重复3次.
PPO的测定参考杨秀清等(2001)的方法, 略有改动:向玻璃试管中依次加入2 mL 0.01 mol·L-1的PBS(pH=6.0), 1 mL 0.1 mol·L-1邻苯二酚和40 μL粗酶液.30 ℃水浴10 min, 于420 nm波长下测定吸光度.所有操作重复3次.
酶活力(U·g-1·min-1)单位定义为每分钟吸光度(D值)增加0.01.具体计算公式如下:
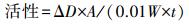 | (1) |
3 结果与分析(Results and analysis)3.1 空心菜体内的Phe15数量变化采用浸根方式将功能菌Phe15定殖到空心菜体内, 借助稀释涂布平板计数法和抗生素抗性标记筛选, 研究了功能内生细菌在蔬菜体内的定殖情况.结果如表 1所示, 菌株Phe15可在空心菜根表、根内和茎叶中良好定殖.在12 d的培养时间内, 空心菜各部位Phe15的数量均呈现出先增加后降低的趋势.根表和叶内Phe15数量在第2 d时达到最大, 分别为5.84 log CFU·g-1 (以鲜重计, f.w)和4.63 log CFU·g-1;而根内Phe15数量则在第5 d时达到最高值, 为5.73 log CFU·g-1.Phe15在蔬菜不同部位数量不同, 呈现如下趋势:根表>根内>茎叶.此外, 成功定殖于蔬菜根表的功能内生细菌可扩散至HNS中, 但数量明显少于植物体内, 且随着时间的延长其数量迅速减少.HNS中Phe15数量在定殖第2 d达到最大, 与根表和茎叶中细菌数量变化趋势一致.
表 1(Table 1)
| 表 1 处理期间培养液及空心菜不同部位Phe15细菌数量变化 Table 1 The cell counts of strain Phe15 in HNS and Water spinach | ||||||||||||||||||||||||||||||||||||||
表 1 处理期间培养液及空心菜不同部位Phe15细菌数量变化 Table 1 The cell counts of strain Phe15 in HNS and Water spinach
| ||||||||||||||||||||||||||||||||||||||
上述结果表明, 采用浸根的方式可以将功能内生细菌Phe15定殖于空心菜根部, 随着培养时间的延长, 根系中的内生细菌可逐渐扩散传输至空心菜地上部分.
3.2 接种Phe15对蔬菜体内及其亚细胞组分中菲含量的影响如图 2所示, 功能菌的定殖可以显著降低空心菜体内及其亚细胞组分PAHs含量.接种功能菌后, 空心菜体内菲含量均低于未接菌处理组.接种功能内生细菌后, 茎叶部PAHs含量在第2 d和第5 d显著低于CP(p < 0.05), 根部菲含量在整体处理期间均显著低于CP(p < 0.05或p < 0.01).处理结束时, 空心菜茎叶中菲含量减少了26.24%, 根部菲含量减少了86.34%.菌株Phe15在空心菜根部的定殖效能优于茎叶部.
图 2(Fig. 2)
 |
| 图 2 空心菜茎叶与根及各亚细胞组分中菲含量(CP表示未接菌对照, CPR表示接菌处理, CW表示细胞壁, CO表示细胞器, CM表示细胞膜, CS表示细胞液;*表示存在显著差异(p < 0.05), **表示存在极显著差异(p < 0.01)) Fig. 2Concentrations of phenanthrene in shoots and roots and their subcellular components of Water spinach with and without mixed bacteria inoculation (mg·kg-1)(CP means no bacteria control; CPR means inoculated with mixed bacteria. CW means cell wall; CO means cell organelle; CM means cell membrane; CS means cell soluble. * and ** indicate significant differences (p < 0.05) and (p < 0.01) between CP and CPR) |
在亚细胞水平, 与对照相比, 接菌处理可有效降低不同亚细胞组分中菲的含量.空心菜茎叶部各亚细胞组分CPR处理组的菲含量均低于CP处理组, 且CM中菲的浓度最高.其中CS中的菲含量在第2 d和第5 d显著低于未接菌处理组, CM中的菲含量在第8 d显著低于未接菌处理组(p < 0.05).根部各亚细胞组分中菲浓度在培养时间内均显著低于未接菌处理组(p < 0.05或p < 0.01), 且CS中菲的浓度最高.培养12 d后, 空心菜茎叶部亚细胞水平的菲含量变化情况为CW减少了10.89%, CO减少了10.05%, CM减少了37.32%, CS减少了29.61%, 而根部菲含量变化情况为CW减少了84.01%, CO减少了83.31%, CM减少了66.44%, CS减少了86.55%.亚细胞水平定殖功能菌对CS中PAHs有较好的去除效能.以上结果表明, 定殖菌株Phe15可在一定程度上降低空心菜不同部位及其亚细胞组分菲含量;与其他部位相比, 菌株Phe15在空心菜根部具有更好的定殖效能.
3.3 接种功能菌对空心菜及其亚细胞组分中POD与PPO活性的影响通过测定空心菜体内两种PAHs降解相关酶POD和PPO的活性, 尝试分析功能菌减低空心菜体内PAHs的可能机制.由图 3可以看出, 茎叶部和根部POD活性随时间的增长呈现先增加后降低的趋势, 且变化趋势不明显.接种功能菌之后, 茎叶部POD活性均得到一定程度的增长, 在处理后第8 d POD活性达到最大值(15504.42 U·g-1·min-1);而根部POD活性则在处理后第5 d达到最大值(59682.55 U·g-1·min-1), 此后酶活性开始下降.PPO活性的变化规律与POD略有不同, 接种功能菌之后, 茎叶部PPO活性前期均有所提高, 在第8 d时活性显著增强(p < 0.01), 增长率达到79.86%, 但处理12 d后接种功能内生细菌处理PPO酶活性略低于未加菌的处理;而根部PPO活性在第2 d时略低于未加菌处理, 但之后均得到了提高.定殖功能菌可以提高空心菜体内POD和PPO的活性.
图 3(Fig. 3)
 |
| 图 3 空心菜茎叶与根中POD和PPO活性(×1000 U·g-1·min-1)(CP表示未接菌对照, CPR表示接菌处理, **和*表示存在显著差异(p < 0.01和0.05)) Fig. 3Activity of POD and PPO in shoots and roots of Water spinach with and without mixed bacteria inoculation (×1000 U·g-1·min-1)(CP means no bacteria control; CPR means inoculated with mixed bacteria. * and ** indicate significant differences (p < 0.05) and (p < 0.01) between CP and CPR) |
为了进一步探究POD、PPO活性与PAHs残留含量之间的关系, 对PPO和POD活性与PAHs残留量之间进行了相关性分析, 结果见表 2.从中可以看出, 空心菜茎叶部和根部POD、PPO活性与PAHs残留含量之间呈负相关.其中, 茎叶部PPO活性与菲含量在第8 d呈现显著负相关(p < 0.05).空心菜体内POD、PPO活性会随PAHs含量的下降而提高.
表 2(Table 2)
| 表 2 植物PAHs残留量与酶活性相关性 Table 2 Pearson′s correlation analysis between Activity of POD and PPO and concentrations of phenanthrene in shoots and roots of Water spinach | ||||||||||||||||||||||||||||||||||||
表 2 植物PAHs残留量与酶活性相关性 Table 2 Pearson′s correlation analysis between Activity of POD and PPO and concentrations of phenanthrene in shoots and roots of Water spinach
| ||||||||||||||||||||||||||||||||||||
如图 4所示, CPR处理组各亚细胞组分中两种酶的活性整体高于未加菌处理组.空心菜茎叶部各亚细胞组分POD活性接菌后整体得到提高, CO中接菌后POD活性前期得到显著提高, 到第12 d活性有所下降, CM中活性普遍较低, 但在第2 d得到显著提高, CW和CS中无显著提高.根部各亚细胞组分中POD活性均在接菌后得到提高, 且CW、CO、CM中均在后期达到显著(p < 0.05或p < 0.01), 但CS中无显著变化.接种功能菌可以提高空心菜亚细胞水平POD活性.空心菜各亚细胞组分中PPO活性的变化与POD相似.茎叶部CW中PPO活性在第5 d和第8 d显著提高, CM中第5 d和第12 d显著提高, CO和CS中增加并不显著.根部各亚细胞组分PPO活性前期均有所提高, 但未达到显著水平, 但是第12 d CO中PPO活性有所下降, 根部CM加菌组PPO活性显著低于未加菌组(p < 0.05或p < 0.01).接种功能菌可以提高空心菜亚细胞水平PPO活性.
图 4(Fig. 4)
 |
| 图 4 空心菜茎叶与根各亚细胞组分中POD和PPO活性(×1000 U·g-1·min-1)(CP表示未接菌对照, CPR表示接菌处理, CW表示细胞壁, CO表示细胞器, CM表示细胞膜, CS表示细胞液;**和*表示存在显著差异(p < 0.01和0.05)) Fig. 4Activity of POD and PPO in the subcellular components of shoots and roots of Water spinach with and without mixed bacteria inoculation (×1000 U·g-1·min-1)(CP means no bacteria control; CPR means inoculated with mixed bacteria. CW means cell wall; CO means cell organelle; CM means cell membrane; CS means cell soluble. * and ** indicate significant differences (p < 0.05) and (p < 0.01) between CP and CPR) |
为了进一步探究亚细胞组分中POD、PPO活性与PAHs残留含量之间的关系, 对两者进行了相关性分析, 结果如表 3所示.空心菜各亚细胞组分POD、PPO活性与PAHs含量之间也呈现出与植物中相似的负相关规律.但是植物茎叶部和根部CM中POD活性与PAHs含量之间呈正相关, 茎叶部CO和CM中PPO活性与PAHs含量在第8 d和第12 d呈现正相关, 根部各亚细胞组分PPO活性与PAHs残留含量在第12 d均呈现正相关.整体来看, 空心菜不同部位亚细胞中PAHs浓度含量与POD、PPO的活性呈负相关.
表 3(Table 3)
| 表 3 空心菜不同部位亚细胞组分PAHs含量与酶活性相关性 Table 3 Pearson′s correlation analysis between Activity of POD and PPO and concentrations of phenanthrene in the subcellular components of shoots and roots of Water spinach | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 3 空心菜不同部位亚细胞组分PAHs含量与酶活性相关性 Table 3 Pearson′s correlation analysis between Activity of POD and PPO and concentrations of phenanthrene in the subcellular components of shoots and roots of Water spinach
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
4 讨论(Discussion)功能内生细菌可与宿主植物形成良好的共生体系, 共生体系中宿主植物能够提供功能内生细菌生长、繁殖所需的碳源和稳定的生态区位, 确保功能内生细菌在植物体内良好定殖.本研究中, 通过浸根的方式可将功能内生细菌Phe15定殖到空心菜根表, 随着培养时间的延长该功能菌株可以由根表转移至根内, 随后扩散到空心菜地上部分.培养期间, 菌株在空心菜各部位的数量均呈现出先增加后降低的趋势.Chen等(2019)研究发现, 采用浸根方式可将具有菲降解能力菌株RS1-gfp定殖到包括菠菜、白菜和大白菜在内的蔬菜根表, 且该菌株可从根表进入到植物体内, 培养期间蔬菜体内菌株数量呈现先增加后降低的趋势, 与本研究结果相似.这可能是由于在培养初期, 定殖在植物根表的功能细菌利用其根表吸附的PAHs作为碳源进行快速的生长、繁殖, 数量增多, 减少空心菜对于菲的吸收和积累.到培养中期, 功能菌转移至根内并向空心菜茎叶部扩散, 从而参与降解已经进入植物体内的菲, 进而缓解了菲对空心菜生长的胁迫.随着培养时间的延长, 空心菜体内的菲含量降低, 胁迫作用逐渐缓解, 功能内生细菌的数量也随之减少.同时, 扩散至HNS中的功能菌也可以降解溶液中的菲, 导致溶液中菲的浓度下降, 植物对溶液中菲的吸收也因此下降.
进入植物体内的功能内生细菌可以利用植物体内富集的PAHs作为碳源进行生长、繁殖, 进而削减植物体内PAHs积累(Khan et al., 2013).本研究中, 接种菌株Phe15有效地降低空心菜及其亚细胞组分中菲含量, 根部最高降低93.69%, 叶部最高降低50.26%.Liu等(2014)发现功能菌株Pn2可以定殖到黑麦草体内, 减低黑麦草体内的菲含量, 同未接菌对照相比, 接菌后黑麦草根与茎叶中菲浓度分别降低54%和57%.Zhu等(2017)研究了接种具有菲降解能力的菌株PW7对小麦体内芘含量的影响, 结果表明, 小麦接种PW7培养8 d后, 其茎叶内芘含量降低了35.7%~86.3%, 根内芘含量降低了26.8%~60.1%.以上结果说明, 接种功能内生细菌可有效地减低空心菜体内PAHs积累.
吸收后的PAHs可进一步在植物亚细胞组分进行分配(Gao et al., 2011).本研究表明, 吸收后的PAHs在空心菜亚细胞组分中分配存在差异, 茎叶部CM中菲的浓度最高, 根部CS中菲的浓度最高.定殖功能内生细菌后, 不同亚细胞组分中菲的含量均有所下降, CS中菲的去除效果最佳.Gao等(2013)研究了蒽及其代谢产物在高羊茅亚细胞组分中的分配规律, 发现茎叶部CS中蒽的含量最高, 根部CO中蒽的含量最高.可能是由于本实验分离出了CM, 进一步细化了菲在CM、CS中的不同分配;此外, 本实验培养时间短于Gao等, 菲主要残留在CS中, 并未大量迁移至CO内, 因此本研究中空心菜茎叶CM, 根部CS内菲含量最高, 与Gao等结果有差异.本研究中, 接种功能内生细菌减低了空心菜亚细胞组分PAHs的积累, 可能是由于菌株主要定殖在空心菜细胞间隙中, 其中含有大量营养物质, 如营养盐、糖类和氨基酸等, 能够促进菌株Phe15的生长繁殖、增强空心菜体内菲的降解代谢, 进而阻控菲向CW、CO、CM、CS的转移作用, 改变空心菜体内菲在亚细胞组分中的分配(Afzal et al., 2014; Brader et al., 2014).
植物代谢有机污染物的过程主要包括氧化反应、还原反应、水解作用等, 而这些过程均需要氧化还原酶等相关酶系的参与, POD和PPO是有机污染物代谢研究中受到关注较多的酶(Jeger et al., 2001; Kvesitadze et al., 2009; Sinsabaugh et al., 2010).本课题组前期的研究结果表明, 通过调节植物酶系调控植物PAHs代谢过程可有效地规避植物PAHs污染风险.Gao等(2012)研究表明, 添加抑制剂可降低植物体内POD和PPO的活性, 抑制了植物体内菲的代谢, 增加了菲在高羊茅体内的积累.Gong等(2011)采用水培试验研究了植物酶系活性与PAHs代谢的关系, 结果表明, POD和PPO活性与植物体内PAHs代谢效率呈显著正相关.本研究中, 接种功能内生细菌培养12 d后空心菜茎叶与根部POD和PPO活性均高于未接菌处理组, 且空心菜茎叶部和根部POD、PPO活性与PAHs残留含量之间呈负相关.Kucerová等(1998)研究指出植物体内PCBs降解与POD的含量呈正相关关系.何丽烂等(2004)发现蔬菜种子经耐氨固氮菌浸种处理后, 甘蓝中POD、PPO活性显著增加.孙凯等(2019)研究发现, 接种混合菌Pn2和033显著提高了小麦体内POD和PPO活性(p < 0.05), 同时降低了植物体内PAHs的含量.结果均与本研究类似.本研究中, 接菌和不接菌处理空心菜根部菲的含量差异很大, 但PPO和POD酶的活性差异不显著, 推测是由于植物体内PPO和POD酶活性同时受到污染条件和外源功能菌的双重影响, 接种功能菌和污染胁迫在一定程度上诱导PPO和POD酶活性, 与不接种功能菌相比, 接种功能菌提高了植物体内PPO和POD酶活性;此外, 定殖于植物体内的功能内生细菌能够以植物体内菲作为碳源生长繁殖, 进一步降低根部菲的积累, 在一定程度上缓解菲的胁迫作用, 导致在相同的污染条件下, 接菌和不接菌蔬菜根部PPO和POD酶的活性差异不显著.但关于接种功能菌株对植物体内PAHs分解代谢及相关酶活性变化的内在关系还有待进一步研究.
吸收的PAHs进入植物体内后经过一系列氧化、还原及水解过程形成一些结合产物, 所形成的结合产物最终在液泡、细胞壁等亚细胞组分中代谢, 进而减低PAHs毒性(林庆祺等, 2013).本研究中, 定殖功能菌之后, 接种功能内生细菌提高了植物各亚细胞组分酶活性, 随着培养时间的延长, 空心菜亚细胞PPO和POD酶活性有所下降.相关性分析结果表明, 亚细胞组分中POD、PPO的活性与PAHs含量呈负相关, 表明POD和PPO酶活性的提高降低了亚细胞中PAHs含量, 与上述植物体内的研究结果相似.但是在PAHs浓度较低的CM及降解后期的各亚细胞组分中呈正相关, 可能是由于PAHs浓度低时, 酶活性便会随PAHs含量的降低而下降.空心菜亚细胞组分中, 茎叶部和根部均为CS内的酶活性最高.Ling等(2012)研究了PAHs污染胁迫下高羊茅各亚细胞组分中PPO的活性差异, 发现酶活力主要集中在CS部分, 与本研究结果相似.这可能是由于水溶性较强的结合产物主要集中在CS中, 诱导PAHs代谢相关酶系活性, 导致CS内PPO和POD活性较高.但是关于定殖功能内生细菌促进植物亚细胞PAHs代谢的具体过程和涉及的酶促反应及关键蛋白等信息还有待进一步研究.
5 结论(Conclusions)1) 菌株Phe15可在空心菜体内良好定殖, 菌株Phe15细胞数量在空心菜不同部位分布规律为根表 > 根内 > 茎叶.
2) 接种菌株Phe15有效降低了空心菜不同部位及其亚细胞组分中菲的积累.
3) 接种菌株Phe15可以提高空心菜不同部位及其亚细胞组分POD和PPO活性, 且酶系活性与空心菜体内PAHs积累呈负相关关系.
参考文献
| Afzal M, Khan Q M, Sessitsch A. 2014. Endophytic bacteria: Prospects and applications for the phytoremediation of organic pollutants[J]. Chemosphere, 117: 232-242. DOI:10.1016/j.chemosphere.2014.06.078 |
| Brader G, Compant S, Mitter B, et al. 2014. Metabolic potential of endophytic bacteria[J]. Current Opinionin Biotechnology, 27: 30-37. DOI:10.1016/j.copbio.2013.09.012 |
| Chen S, Ma Z, Li S, et al. 2019. Colonization of polycyclic aromatic hydrocarbon-degrading bacteria on roots reduces the risk of PAH contamination in vegetables[J]. Environment International, 132: 105081. DOI:10.1016/j.envint.2019.105081 |
| Gao Y Z, Cao X Z, Kang F X, et al. 2011. PAHs pass through the cell wall and partition into organelles of arbuscular mycorrhizal roots of ryegrass[J]. Journal of Environmental Quality, 40(2): 653-656. DOI:10.2134/jeq2010.0320 |
| Gao Y Z, Zhang Y, Liu J, et al. 2013. Metabolism and subcellular distribution of anthracene in tall fescue (Festuca arundinacea Schreb.)[J]. Plant and Soil, 365(1/2): 171-182. DOI:10.1007/s11104-012-1386-1 |
| Gao Y Z, Li H, Gong S S. 2012. Ascorbic acid enhances the accumulation of Polycyclic Aromatic Hydrocarbons (PAHs) in Roots of TallFescue (Festuca arundinacea Schreb.)[[J]. Plos One, 7(11): e50467. DOI:10.1371/journal.pone.0050467 |
| 高彦征, 朱利中, 凌婉婷, 等. 2005. 土壤和植物样品的多环芳烃分析方法研究[J]. 农业环境科学学报, 24(5): 1003-1006. DOI:10.3321/j.issn:1672-2043.2005.05.035 |
| Germaine K, Keogh E, Garcia-Cabellos G, et al. 2004. Colonisation of poplar trees by gfp expressing bacterial endophytes[J]. FEMS Microbiology Letters, 48: 109-118. DOI:10.1016/j.femsec.2003.12.009 |
| 龚帅帅, 韩进, 高彦征, 等. 2011. 抑制剂和安全剂对高羊茅根中酶活性和菲代谢的影响[J]. 生态学报, 3(14): 4027-4033. |
| 顾玉骏. 2015. 根表多环芳烃降解细菌的分离筛选及其在植物根表的定殖和效能[D]. 南京: 南京农业大学 |
| 郝再彬, 苍晶, 徐仲. 2004. 植物生理实验[M]. 哈尔滨: 哈尔滨工业大学出版社. |
| 何丽烂, 区炳庆, 温海祥. 2004. 耐氨固氮菌浸种对几种蔬菜SOD, POD等活性的影响初探[J]. 佛山科学技术学院学报(自然科学版), 22(2): 71-74. |
| Iwabuchi T, Harayama S. 1998. Biochemical and molecular characterization of 1-hydroxy-2-naphthote dioxygenase from Nocardioides sp. KP7[J]. The Journal of Biological Chemistry, 273: 833-838. |
| Jeger M J, Spence N J. 2001. Biotic Interactions in Plant-Pathogen Associations[J]. Biotic Interactions in Plant-pathogen Associations: 87-119. |
| Kang F, Chen D, Gao Y, et al. 2010. Distribution of polycyclic aromatic hydrocarbons in subcellular root tissues of ryegrass (Lolium multiflorum Lam.)[J]. BMC Plant Biology, 10(1): 1. DOI:10.1186/1471-2229-10-1 |
| Khan S, Afzal M, Iqbal S, et al. 2013. Plant-bacteria partnerships for the remediation of hydrocarbon contaminated soils[J]. Chemosphere, 90(4): 1317-1332. DOI:10.1016/j.chemosphere.2012.09.045 |
| Kucerová P, Polachova L, Macek T, et al. 1998. Transformation of PCBs by plant cell cultures and relation to the production of plant peroxidases[J]. Internat Biodeterior Biodegrad, 42: 245-250. |
| Kvesitadze E, Sadunishvili T, Kvesitadze G. 2009. Mechanisms of organic contaminants uptake and degradation in plants[J]. World Academy of Science, Engineering and Technology, 55: 458-468. |
| 林庆祺, 蔡信德, 王诗忠, 等. 2013. 植物吸收, 迁移和代谢有机污染物的机理及影响因素[J]. 农业环境科学学报, 32(4): 661-667. |
| Ling W T, Lu X D, Gao Y Z, et al. 2012. Polyphenol oxidase activity in subcellular fractions of tall fescue contaminated by polycyclic aromatic hydrocarbons[J]. Journal of Environmental Quality, 41(3): 807-813. DOI:10.2134/jeq2011.0461 |
| Liu J, Liu S, Sun K, et al. 2014. Colonization on root surface by a Phenanthrene-degrading endophytic bacterium and its application for reducing plant phenanthrene contamination[J]. Plos one, 9(9): e108249. DOI:10.1371/journal.pone.0108249 |
| Liu J, Xiang Y B, Zhang Z M, et al. 2017. Inoculation of a phenanthrene-degrading endophytic bacterium reduces the phenanthrene level and alters the bacterial community structure in wheat[J]. Applied Microbiology and Biotechnology, 101(12): 5199-5212. DOI:10.1007/s00253-017-8247-z |
| 李腾武, 高希武, 郑炳宗, 等. 1999. 小菜蛾不同亚细胞层梭酸醋酶的性质研究[J]. 农药学学报, 1(2): 47-53. |
| Ma C L, Ye S Y, Lin T, et al. 2014. Source apportionment of polycyclic aromatic hydrocarbons in soils of wetlands in the Liao River Delta, Northeast China[J]. Marine Pollution Bulletin, 80(1/2): 160-167. |
| Sandermann Jr H. 1994. Higher-plant metabolism of xenobiotics-the green liver concept[J]. Pharmacogenetics, 4(5): 225-241. DOI:10.1097/00008571-199410000-00001 |
| Shah K, Kumar R G, Verma S, et al. 2001. Effect of cadmium on lipid peroxidation, superoxide anion generation and activities of antioxidant enzymes in growing rice seedlings[J]. Plant Science, 161(6): 1135-1144. DOI:10.1016/S0168-9452(01)00517-9 |
| Sinsabaugh R L. 2010. Phenol oxidase, peroxidase and organic matter dynamics of soil[J]. Soil Biology and Biochemisty, 42(3): 391-404. DOI:10.1016/j.soilbio.2009.10.014 |
| Sun K, Liu J, Jin L, et al. 2014. Utilizing pyrene-degrading endophytic bacteria to reduce the risk of plant pyrene contamination[J]. Plant and Soil, 374(1/2): 251-262. |
| 孙凯, 李舜尧, 陈明雨, 等. 2019. 接种混合功能细菌降低黑麦草体内菲和芘污染的机理初探[J]. 环境科学学报, 39(4): 1303-1311. |
| Tao S, Jiao X C, Chen S H, et al. 2006. Accumulation and distribution of polycyclic aromatic hydrocarbons in rice (Oryza sativa)[J]. Environmental Pollution, 140(3): 406-415. DOI:10.1016/j.envpol.2005.08.004 |
| Wang C H, Wu S H, Zhou S L, et al. 2015. Polycyclic aromatic hydrocarbons in soils from urban to rural areas in Nanjing: Concentration, source, spatial distribution, and potential human health risk[J]. Science of the Total Environment, 527: 375-383. |
| Wang J, Zhang X F, Ling W T, et al. 2017. Contamination and health risk assessment of PAHs in soils and crops in industrial areas of the Yangtze River Delta region, China[J]. Chemosphere, 168: 976-987. DOI:10.1016/j.chemosphere.2016.10.113 |
| 相妍冰. 2017. 功能内生细菌Pn2定殖对小麦体内菲污染和土著细菌群落结构的影响[D]. 南京: 南京农业大学 |
| Yan C L, Hong Y T, Fu S Z, et al. 1997. Effect of Cd, Pb stress on scarenging system of activated oxygen in leaves of tobacco[J]. Acta ecologica Sinica, 17(5): 488-492. |
| 杨秀清, 原海云, 焦晓光. 2001. 铜、锌元素对华北落叶松苗期酶活性影响[J]. 山西农业大学学报, 21(3): 277-280. DOI:10.3969/j.issn.1671-8151.2001.03.023 |
| Yu H Y, Li T J, Liu Y, et al. 2019. Spatial distribution of polycyclic aromatic hydrocarbon contamination in urban soil of China[J]. Chemosphere, 230: 498-509. DOI:10.1016/j.chemosphere.2019.05.006 |
| Zhu X Z, Wang W Q, Crowley D E, et al. 2017. The endophytic bacterium Serratia sp. PW7 degrades pyrene in wheat[J]. Environmental Science and Pollution Research, 24(7): 6648-6656. DOI:10.1007/s11356-016-8345-y |
| Zhu X Z, Wang W Q, Sun K, et al. 2017. Inoculating wheat (Triticum aestivum L.) with the endophytic bacterium Serratia sp. PW7 to reduce pyrene contamination[J]. International Journal of Phytoremediation, 19(8): 718-824. DOI:10.1080/15226514.2017.1284747 |
