
 , 董世魁2, 彭明春3
, 董世魁2, 彭明春31. 交通运输部水运科学研究所, 北京 100088;
2. 北京师范大学环境学院, 北京 100088;
3. 云南大学生态学与地植物学研究所, 昆明 650091
收稿日期: 2018-01-08; 修回日期: 2018-02-09; 录用日期: 2018-02-09
基金项目: 国家自然科学基金(No.51509116);国家重点研发计划项目(No.2016YFC0503601)
作者简介: 李晋鹏(1984-), 男, E-mail:lijp_cool@126.com, lijinpeng@wti.ac.cn
通讯作者(责任作者): LI Jinpeng, E-mail:lijp_cool@126.com, lijinpeng@wti.ac.cn
摘要: 梯级水电开发虽然可以最大程度地利用河流的水能资源,但同时也是对河流生态系统的大规模扰动.大型底栖动物被认为是开展梯级水坝水生态系统变化和演替研究的指示生物类群.本文选取澜沧江中下游水电基地为研究区,于2016年开展漫湾和大朝山梯级水坝库区大型底栖动物群落采样调查,分析梯级库区大型底栖动物的群落结构和分布格局,基于生物指数(BI)开展水质生物学评价.结果表明:大型底栖动物物种组成以寡毛纲、软体动物门及昆虫纲的摇蚊类为主,优势种则以软体动物门的河蚬占绝对优势;群落密度和生物量分布格局表现为静水区显著高于过渡区;静水区功能摄食群密度组成以收集者(GC)和滤食者(FC)占优势,生物量组成则以滤食者(FC)占优势;ABC曲线和生物指数分析表明,上游漫湾库区群落稳定性及水质优于下游大朝山库区,反映了梯级水坝沿着河流纵向梯度水环境逐渐变差的趋势,澜沧江梯级水坝库区大型底栖动物群落结构、分布格局和库区水质状况受梯级水坝运行影响表现为明显的累积效应和富营养化效应.
关键词:大型底栖动物梯级水坝水质生物学评价澜沧江
The structure of benthic macrofauna and bioassessment of water quality in cascade dammed reservoirs of Lancang River
LI Jinpeng1
, DONG Shikui2, PENG Mingchun3 1. China Waterborne Transport Research Institute, Beijing 100088;
2. School of Environment, Beijing Normal University, Beijing 100088;
3. Institute of Ecology and Geobotany, Yunnan University, Kunming 650091
Received 8 January 2018; received in revised from 9 February 2018; accepted 9 February 2018
Supported by the National Natural Science Foundation of China (No.51509116) and the National Key Research & Development Plan(No.2016YFC0503601)
Biography: LI Jinpeng(1984—), male, E-mail:lijp_cool@126.com, lijinpeng@wti.ac.cn
Abstract: Although the cascading hydropower exploitation can maximize utilization the water power resources, it can result in a large-scale disturbance on the river ecosystem. Benthic macrofauna are generally considered as an indicator of biological group for the aquatic ecosystem change and succession in cascade dammed reservoirs. In this study, the middle and lower reaches of Lancang River were selected as study areas. The benthic macrofaunas were sampled in cascade dammed reservoirs including Manwan and Dachaoshan in 2016. The structure, distribution pattern of benthic macrofauna, and the bioassessment of water quality based on the biotic index (BI) were analyzed and performed. The results showed that the species composition of benthic macrofauna were mainly Oligochaeta, Mollusca, and chironomids, which belonged to Insecta. The dominant species of benthic macrofauna was Corbicula fluminea, which belonged to Mullusca. The density and biomass of benthic macrofaunas in the lacustrine zone were significantly higher than those in transition zone. As for the functional feeding group, the density composition of benthic macrofaunas was dominated by the gatherer-collector (GC) and filter-collector (FC), while the biomass composition was dominated by the filter-collector (FC). The abundance/biomass curves (ABC) and biotic index (BI) analysis indicated that the stability of benthic macrofauna and the water quality in upstream Manwan reservoir were better than that in downstream Dachaoshan reservoir, implying that the water quality gradually got worse in these cascading reservoirs along the longitudinal gradient of the river. The structure, distribution pattern of benthic macrofauna, and water quality revealed obviously cumulative and eutrophication effects caused by cascading reservoir operation in Lancang River.
Key words: benthic macrofaunacascading reservoirsbioassessment of water qualityLancang River
1 引言(Introduction)河流不仅为人类和淡水生态系统提供了最重要的可再生水资源, 同时也为众多生物提供了最重要的栖息地并作为主要通道参与生物圈的物质循环(董哲仁等, 2010;Helson et al., 2013).近年来, 全球河流生态系统受高强度的人类活动特别是水坝工程建设的干扰严重(Nilsson et al., 2005).大型水坝的建设和运行改变了自然河流的水文节律和泥沙的沉积迁移速率, 并对水生生物造成深远影响, 已成为河流生物多样性丧失的重要原因(Mccartney et al., 2000;Chen et al., 2013).梯级水电开发在河流上形成一系列首尾相接的河道型水库, 虽然可以最大程度的利用水能资源, 但同时也是对河流生态系统的大规模扰动, 破坏了河流的生态连通性和纵向连通性(Petesse et al., 2007; Li et al., 2013), 其生态效应和影响远比单一水坝复杂, 具有复杂性、累积性、潜在性和系统性的特点(翟红娟等, 2007;董哲仁等, 2010;王超, 2017).大型底栖动物作为水生态系统的重要生物群落, 对环境条件变化敏感, 在水体的物质循环和能量流动过程起着重要作用, 是开展梯级水坝水生态系统变化和演替研究的指示生物类群(罗欢等, 2016;李晋鹏等, 2017;慕林青等, 2018).大型底栖动物的物种组成、群落结构、空间分布和季节动态可以很好地反映水环境和水生态条件的变化, 受到河流生态系统保护研究领域的广泛关注(王强等, 2017;张敏等, 2017).
我国西南地区作为世界梯级水电开发的中心, 同时也是国际生物多样性保护的热点地区(Grumbine et al., 2011).区域内的澜沧江中下游作为中国12个重点开发的水电基地, 目前已基本完成梯级开发(Dore et al., 2004).漫湾和大朝山水坝作为澜沧江干流先后投入运行的第1座(1995年)和第2座水坝(2003年), 在上游功果桥(2013年)和小湾(2010年)水坝蓄水运行后, 澜沧江中下游梯级水坝群已连续稳定运行多年, 其库区水环境条件和大型底栖动物群落保持相对稳定(李晋鹏等, 2017).当前, 国内外在大型河流上开展梯级水坝库区大型底栖动物群落结构和时空分布格局研究相对较少(邵美玲等, 2007;陈浒等, 2010;简东等, 2010;Almeida et al., 2013; 李斌等, 2013;Tagliaferro et al., 2013;罗欢等, 2016;李晋鹏等, 2017), 且水库生态系统的变化及演替日益成为水生态学关注的研究热点(张敏等, 2017).因此, 本研究于2016年选取澜沧江干流漫湾和大朝山梯级水坝库区开展大型底栖动物群落调查, 并结合历史调研数据(1996年, 自然河流时期), 分析梯级水坝运行对上下游两个库区大型底栖动物群落的种类组成、优势种、群落密度和生物量、功能摄食群、群落类型及稳定性的影响, 基于生物指数(Biotic Index, BI)开展库区水质生物学评价, 在此基础上阐明梯级水坝运行对大型底栖动物群落的生态效应.
2 研究区域与研究方法(Study area and methods)2.1 研究区域概况选取云南省澜沧江干流先后投入运营的漫湾(M)和大朝山(D)梯级水坝库区为研究区(图 1), 其分别为澜沧江中下游2库8个梯级水电开发中的第3级和第4级(陈丽晖等, 2000), 均为径流式水坝.其中, 漫湾水坝坝高132 m, 正常蓄水位海拔994 m, 总库容10.6×108 m3, 水库面积23.6 km2, 装机容量150×104 kW, 干流回水70 km至上游的小湾水坝附近(王忠泽等, 2000).大朝山水坝位于漫湾水坝下游, 正常蓄水位海拔899 m, 坝高111 m, 总库容9.4×108 m3, 水库面积26.25 km2, 装机容量135×104 kW, 干流回水91 km至漫湾水坝附近(欧阳慧等, 2010;贾志翔, 2011).研究区位于我国西南地区, 地貌为高山峡谷, 气候类型为亚热带低纬度山地季风气候, 年内分为明显的干湿两季, 5—10月受西南季风的影响为雨季, 11月—次年4月受大陆西风影响为旱季.
图 1(Fig. 1)
 |
| 图 1 澜沧江漫湾和大朝山库区及大型底栖动物采样点位置 Fig. 1Location of the Manwan and Dachaoshan reservoir and the sampling sites of benthic macrofauna in the Lancang River |
2.2 采样调查本研究于2016年开展漫湾和大朝山梯级水坝库区大型底栖动物群落及分布格局采样调查.依据建坝河流沿河流纵向梯度上的生境条件划分(Petesse et al., 2007; Perbiche-Neves et al., 2011; 李晋鹏等, 2017), 漫湾和大朝山库区分别设置6个采样点(图 1), 共计12个采样点.其中, M1、M2、D1、D2、D3生境划分位于静水区;M3、M4、D4、D5生境划分位于过渡区;M5、M6和D6生境划分位于河流区(高水位为静水区, 低水位为河流区).每个采样点分别于旱季(4月, 高水位)和雨季(10月, 低水位)进行2期大型底栖动物群落调查.
大型底栖动物调查采用开口面积1/50 m2的Peterson采样器对12个采样点(每个采样点3个重复)采集底泥样品, 经40目尼龙筛淘洗过滤后, 将大型底栖动物用镊子挑出并放入盛有75%的酒精的50 mL塑料标本瓶中保存(王德铭等, 1991;张觉民等, 1991).将调查样品带回实验室参考相关资料(刘月英等, 1979;Morse等, 1994;韩茂森等, 1995)进行物种分类和鉴定(一般鉴定到种, 部分为属), 分别统计大型底栖动物不同种类的个体数量, 并用天平称量其湿重, 最后计算出各个采样点的个体密度(ind·m-2)和生物量(g·m-2).其中, 生境划分位于河流区的漫湾库区M4、M5和大朝山库区D6采样点的2期调查未采到大型底栖动物, D5采样点雨季未采到大型底栖动物.
2.3 数据处理2.3.1 优势种分析采用相对重要性指数(IRI)进行优势种分析(孙伟胜等, 2015;李晋鹏等, 2017):
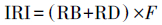 | (1) |
2.3.2 群落聚类分析各采样点的大型底栖动物丰度数据经过对数转换(Y=log(y+1))后, 构建Bray-Curtis相似矩阵, 利用组平均法进行等级聚类(CLUSTER)分析, 并采用相似性百分比分析(SIMPER)计算同一群组的平均相似性和主要贡献物种, 以上分析可通过大型多元统计分析软件Primer5.2.8完成(周红等2003;徐勇等, 2016).
2.3.3 丰度/生物量比较曲线丰度/生物量比较曲线(Abundance/Biomass Curves, ABC)分析可以较好地反映大型底栖动物群落受环境污染和受干扰的程度.据相关研究(孙伟胜等, 2015;王宝强等, 2015), 如生物量曲线位于丰度曲线之上, 表明群落处于未干扰状态;两曲线相交, 表明群落处于中等干扰状态;丰度曲线位于生物量曲线之上, 表明群落处于严重干扰状态.本研究选取漫湾和大朝山库区坝前具有代表性的采样点M1和D1进行ABC曲线分析.
2.3.4 功能摄食群的划分参照Barbour等(1999)关于大型底栖动物功能摄食群(Functional Feeding Group, FFG)的划分标准, 研究区共划分为包括滤食者(Filter-Collector, FC)、掠食者(Predator, PR)、杂食者(Omnivore, OM)、撕食者(Shredder, SH)、收集者(Gather-Collector, GC)和刮食者(Scraper, SC) 6种类型.
2.3.5 水质生物学评价采用生物指数(BI)进行水质生物学评价, 该指数综合了大型底栖动物的耐污能力和物种多样性, 并广泛应用于国内外水质生物学评价研究(吴东浩等, 2011; Chang et al., 2017):
 | (2) |
表 1(Table 1)
| 表 1 生物指数(BI)与水质评价分级标准 Table 1 Classification standard of biotic index (BI) and water quality assessment | ||||||||||
表 1 生物指数(BI)与水质评价分级标准 Table 1 Classification standard of biotic index (BI) and water quality assessment
| ||||||||||
表 2(Table 2)
| 表 2 漫湾(M)和大朝山(D)库区底栖动物种类组成及相对重要性指数(IRI)分析 Table 2 Species composition and index of relative importance (IRI) of benthic macroinvertebrates in Manwan (M) and Dachaoshan (D) reservoir | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 2 漫湾(M)和大朝山(D)库区底栖动物种类组成及相对重要性指数(IRI)分析 Table 2 Species composition and index of relative importance (IRI) of benthic macroinvertebrates in Manwan (M) and Dachaoshan (D) reservoir
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
3 结果与分析(Results and analysis)3.1 大型底栖动物种类组成及优势种分析2016年调查共采集到大型底栖动物16种(表 2), 漫湾库区12种, 其中, 昆虫纲种类(5种, 42%)和软体动物门种类(4种, 33%)占优势, 年内大型底栖动物种类旱季(9种)多于雨季(8种);大朝山库区大型底栖动物11种, 其中, 寡毛纲种类(5种, 45%)和昆虫纲种类(5种, 45%)占优势, 年内大型底栖动物种类雨季(9种)多于旱季(7种).
优势种分析表明, 两库区均以河蚬(Corbicula fluminea)为优势种, 并占绝对优势.具体表现为下游梯级大朝山库区河蚬相对重要性指数明显高于上游梯级漫湾库区, 且各库区旱季均高于雨季.
3.2 大型底栖动物密度和生物量沿河流纵向梯度生境条件的变化, 漫湾和大朝山库区各采样点的大型底栖动物密度和生物量均表现为坝前静水区显著高于过渡区, 河流区均未采集到大型底栖动物(图 2).大型底栖动物密度和生物量年内动态变化表现为:除D3(雨季高于旱季)外, 其余各采样点旱季均明显高于雨季.
图 2(Fig. 2)
 |
| 图 2 漫湾(M)和大朝山(D)库区底栖动物密度和生物量的变化(A:2016-4; O:2016-10) Fig. 2Variance of benthic macrofauna density and biomass in Manwan (M) and Dachaoshan (D) reservoir |
具体来看, 静水区M1和M2样点大型底栖动物群落密度和生物量以软体动物占绝对优势, D1样点大型底栖动物密度以寡毛纲占优势, 生物量以软体动物门和寡毛纲占优势, D2和D3样点大型底栖动物密度以软体动物门和寡毛纲占优势, 生物量则以软体动物门占绝对优势.过渡区M3样点密度和生物量旱季以昆虫纲种类和软体动物门占优势, 雨季以软体动物占绝对优势, D4样点雨季、旱季均以软体动物占绝对优势, D5样点旱季以软体动物和寡毛纲占优势, 雨季则未采到大型底栖动物.
3.3 大型底栖动物群落聚类分析CLUSTER等级聚类分析结果表明, 基于50%的Bray-Curtis相似性水平将两库区大型底栖动物共划分为6个群组(图 3), 其中, 静水区4个群组, 过渡区2个群组, 静水区和过渡区共有1个群组(表 3), 具体结果如下.
图 3(Fig. 3)
 |
| 图 3 漫湾(M)和大朝山(D)库区各采样点大型底栖动物等级聚类(A:2016-4; O:2016-10) Fig. 3The hierarchical cluster of benthic macrofauna sampling sites in Manwan (M) and Dachaoshan (D) reservoir |
表 3(Table 3)
| 表 3 漫湾(M)和大朝山(D)水坝库区底栖动物群落的密度和生物量 Table 3 The abundance and biomass of benthic macrofauna in Manwan (M) and Dachaoshan (D) reservoir | ||||||||||||||||||||||||||||||||||||||||
表 3 漫湾(M)和大朝山(D)水坝库区底栖动物群落的密度和生物量 Table 3 The abundance and biomass of benthic macrofauna in Manwan (M) and Dachaoshan (D) reservoir
| ||||||||||||||||||||||||||||||||||||||||
群组1:分布于漫湾库区静水区, 密度((2490±300) ind·m-2)及生物量((1064±457) g·m-2)最大, 包括采样点M2-A、M1-A, 平均相似性为59.64%, 主要贡献种为河蚬(48.71%)、霍普水丝蚓(29.12%)和苏氏尾鳃蚓(22.17%).
群组2:分布于大朝山库区静水区, 密度((900±264) ind·m-2)及生物量((290±191) g·m-2)次之, 包括采样点D3-O、D1-O、D2-A、D1-A, 平均相似性为68.44%, 主要贡献种为霍普水丝蚓(31.11%)、河蚬(24.91%)、菱跗摇蚊(20.51%)和铁线单向蚓(19.08%).
群组3:分布于漫湾库区和大朝山库区静水区, 密度((390±165) ind·m-2)及生物量((121±58) g·m-2)较高, 包括采样点M2-O、M1-O、D2-O, 平均相似性为60.57%, 主要贡献种为河蚬(29.54%)、水丝蚓(25.27%)、花纹前突摇蚊(23.18%)、苏氏尾鳃蚓(7.59%)、小云多足摇蚊(7.59%).
群组4:分布于大朝山库区过渡区和漫湾库区过渡区, 密度((245±120) ind·m-2)相对较高, 生物量((3.26±0.95) g·m-2)较低, 包括采样点D4-O、D4-A、M3-A, 平均相似性为56.09%, 主要贡献种为河蚬(100%).
群组5:分布于大朝山库区静水区和过渡区, 包括采样点D5-A、D3-A, 密度((180±30) ind·m-2)及生物量((9.69±6.69) g·m-2)较低, 平均相似性为56.40%, 主要贡献种为克拉伯水丝蚓(54.49%)和河蚬(45.51%).
群组6:密度(90 ind·m-2)及生物量(0.45 g·m-2)最小, 分布于漫湾库区过渡区, 包括采样点M3-O.
3.4 丰度/生物量比较曲线(ABC曲线)分析ABC曲线分析表明, 漫湾和大朝山库区坝前静水区M1和D1样点大型底栖动物群落受干扰程度不同(图 4).漫湾库区M1样点大型底栖动物群落较为稳定, 生物量曲线始终位于丰度曲线上方, 雨季丰度曲线起点稍低于旱季.大朝山库区D1样点大型底栖动物群落年内稳定性变化较大, 旱季群落较为稳定, 雨季生物量曲线和丰度曲线相交, 显示大型底栖动物群落受到中等程度的干扰.
图 4(Fig. 4)
 |
| 图 4 漫湾和大朝山水坝库区采样点(D1、M1)大型底栖动物群落ABC曲线(A:2016-4; O:2016-10) Fig. 4Abundance/biomass curves of benthic macrofauna sampling sites (D1、M1) in Manwan and Dachaoshan reservoir |
3.5 大型底栖动物功能摄食群(FFG)变化漫湾库区静水区M1样点功能摄食群密度和生物量组成以滤食者(FC)占绝对优势, M2样点密度组成旱季以滤食者(FC)占绝对优势, 雨季以收集者(GC)和滤食者(FC)占优势, 生物量组成则以滤食者(FC)占绝对优势;过渡区M3样点密度和生物量组成旱季以撕食者(SH)和滤食者(FC)占优势, 雨季则以收集者(GC)占绝对优势.
大朝山库区静水区D1样点密度组成以收集者(GC)占优势, D2样点以滤食者(FC)占优势, D3样点以滤食者(FC)和收集者(GC)占优势, 而D1、D2和D3样点生物量组成均以滤食者(FC)占优势;过渡区D4样点密度和生物量组成以滤食者(FC)占优势, D5样点则以滤食者(FC)和收集者(GC)占优势(图 5).
图 5(Fig. 5)
 |
| 图 5 漫湾(M)和大朝山(D)库区底栖动物功能摄食群(FFG)组成(A: 2016-4; B: 2016-10) Fig. 5The functional feeding group (FFG) composition of benthic macrofauna in Manwan (M) and Dachaoshan (D) reservoir |
3.6 水质生物学评价生物指数(BI)评价结果表明(表 4), 2016年漫湾库区水质(Ⅰ~Ⅱ)明显优于大朝山库区(Ⅰ~Ⅲ).两库区静水区除D3(雨季水质优于旱季)外, M1、M2、D1和D2样点旱季水质均优于雨季;漫湾库区过渡区M3样点则雨季好于旱季, 大朝山库区过渡区D4样点年内变化不大.各库区沿河流纵向梯度, 漫湾库区旱季过渡区M3样点水质相对较差, 雨季静水区M2样点(近坝前)水质相对较差;大朝山库区旱季静水区D3样点(近过渡区)水质相对较差, 雨季静水区D1样点(坝前)水质相对较差.
表 4(Table 4)
| 表 4 漫湾(M)和大朝山(D)水坝库区各采样点的生物指数(BI)及水质评价 Table 4 The biotic index (BI) of sampling sites and water quality assessment in Manwan (M) and Dachaoshan (D) reservoir | |||||||||||||||||||||||||||||||||||||||||||||||||||||
表 4 漫湾(M)和大朝山(D)水坝库区各采样点的生物指数(BI)及水质评价 Table 4 The biotic index (BI) of sampling sites and water quality assessment in Manwan (M) and Dachaoshan (D) reservoir
| |||||||||||||||||||||||||||||||||||||||||||||||||||||
4 讨论(Discussion)4.1 梯级水坝库区的大型底栖动物群落结构及分布格局澜沧江中下游梯级水电开发前, 该河段生境为多样性的急流生境(Li et al., 2013), 大型底栖动物多为适应急流、富氧、贫营养的蜉蝣目(Ephemeroptera)、襀翅目(Plecoptera)和毛翅目(Trichoptera)的种类(王川等, 2013;李晋鹏等, 2017).据1996年历史调研资料, 大型底栖动物密度旱季(80~300 ind·m-2)大于雨季(15~30 ind·m-2), 生物量也表现为旱季(0.18~0.68 g·m-2)大于雨季(0.06~0.07 g·m-2)(王忠泽等, 2000).
梯级水坝运行蓄水后库区的水文、水环境、沉积物、悬浮物和水位波动都是引起大型底栖动物群落结构及演替变化的主要原因(王宝强等, 2015).据2016年调查, 漫湾库区坝前静水区水深40~45 m, 大朝山库区为50~60 m, 淤积明显, 有机物及泥沙的沉积为大型底栖动物的定居、生长和繁殖提供了良好的条件.两库区大型底栖动物物种数增加明显, 物种组成以寡毛纲、软体动物及昆虫纲的摇蚊类为主, 优势种则以软体动物门的河蚬占绝对优势, 这一演替趋势与红水河、乌江和武江梯级的研究结果相同(简东等, 2010;李斌等, 2013;罗欢等, 2016).寡毛纲种类功能摄食群划分为收集者(GC), 指示了库区有机质丰富的淤泥沉积环境, 优势种河蚬则为滤食者(FC), 指示了静水沙质和淤泥质的沉积环境(刘其根等, 2012;罗欢等, 2016).
澜沧江梯级水坝的建设和蓄水运行切割了原自然河流的纵向连通性, 多样性的急流生境转变为包括静水区、过渡区和河流区相间的梯级库区生境类型(Petesse et al., 2007; Perbiche-Neves et al., 2011; 李晋鹏等, 2017).大型底栖动物群落及分布格局表现为不同的生境区差异明显, 静水区密度和生物量显著高于过渡区, 且高水位的旱季明显高于雨季.相对稳定的生境条件是静水区域大型底栖动物高密度和生物量的主要原因(张敏等, 2017).静水区大型底栖动物功能摄食群密度组成以滤食者(FC)和收集者(GC)占优势, 生物量组成则以滤食者(FC)占优势.过渡区由于受水库径流调节及周期性水位变动的影响(张敏等, 2017), 大型底栖动物密度和生物量显著低于静水区, 旱季高水位时期明显高于雨季低水位时期.
4.2 大型底栖动物群落稳定性与梯级水坝库区水质生物学评价大型底栖动物的群落组成和稳定性不仅可以指示库区的沉积环境, 而且可以表明库区水质的变化情况(邵美玲等, 2006).本研究中的ABC曲线分析表明, 上游漫湾库区静水区大型底栖动物群落年内变化较为稳定, 受干扰程度较小, 而下游大朝山库区大型底栖动物旱季较为稳定, 雨季则受中等程度的干扰, 反映梯级水坝沿着河流纵向梯度水环境受到污染的趋势(孙伟胜等, 2015).基于生物指数的水质生物学评价也同时表明, 上游梯级漫湾库区水质明显优于下游大朝山库区, 且大多数静水区采样点旱季水质均优于雨季, 过渡区采样点雨季好于旱季或变化不大.这一研究结果与香溪河流域梯级水库结论相同, 表现为沿着梯级水库水质有逐渐恶化的趋势(邵美玲等, 2007).从生物指数的年内变化来看, 各库区沿河流纵向梯度, 两库区均表现为旱季静水区近过渡区及过渡区水质相对较差, 雨季则静水区坝前及近坝前水质相对较差.ABC曲线和生物指数分析清晰表明了大型底栖动物对梯级水坝库区水质变化有较高的敏感性(吴东浩等, 2011;霍堂斌等, 2012; Chang et al., 2017).
4.3 梯级水坝运行对大型底栖动物群落的生态效应梯级水坝对河流生态系统的影响具有明显的累积效应和富营养化效应(Naiman et al., 2000).澜沧江梯级水坝沿河流纵向梯度, 大型底栖动物优势种河蚬的相对重要性指数为下游梯级大朝山库区明显高于上游梯级漫湾库区, 表现出明显的累积效应.ABC曲线和生物指数分析表明, 下游梯级库区大型底栖动物群落稳定性和水质相比上游梯级库区相对较差, 表现为水质和大型底栖动物群落稳定性的累积效应.同时, 梯级水坝运行对水生生态系统也表现出富营养化效应, 本研究下游大朝山库区静水区寡毛纲及摇蚊种类丰度相对上游库区较高, 显示出库区静水区水体的富营养化及有机污染相对较重的生态特征(邵美玲等, 2006;简东等, 2010;吴涛等, 2016).
此外, 研究区的漫湾和大朝山梯级均为径流式水坝, 其库区的水文、水环境及泥沙的沉积状况除受自身调节影响, 还受上游梯级小湾蓄水式水坝及流域梯级的调度运行影响.梯级水坝上下游不同的水库类型及其调度运行方式、水库形态、水体滞留时间、营养物质的沉积及分布、水位及其波动状况均可引起底栖动物分布格局及动态变化趋于复杂(张敏等, 2017).
5 结论(Conclusions)1) 2016年在澜沧江的大朝山库区和漫湾库区调查共采集到大型底栖动物16种, 物种组成以寡毛纲、软体动物门及昆虫纲的摇蚊类为主.优势种均以河蚬占绝对优势, 下游梯级大朝山库区河蚬的相对重要性指数明显高于上游梯级漫湾库区, 表现为明显的累积效应.
2) 沿河流纵向梯度生境条件的变化, 两梯级库区大型底栖动物密度和生物量均表现为坝前静水区显著高于过渡区.下游大朝山库区静水区寡毛纲及摇蚊种类丰度显著高于上游漫湾库区, 表现出水体的富营养化及有机污染相对较重的生态特征.
3) 两梯级库区静水区大型底栖动物功能摄食群密度组成以收集者(GC)和滤食者(FC)占优势, 生物量组成则以滤食者(FC)占优势.寡毛纲种类为收集者(GC), 指示了库区有机质丰富的淤泥沉积环境, 优势种河蚬则为滤食者(FC), 指示了库区静水沙质和淤泥质的沉积环境.
4) CLUSTER等级聚类将两梯级库区大型底栖动物共划分为6个群组, 其中, 静水区4个群组, 过渡区2个群组, 静水区和过渡区共有1个群组, 大型底栖动物群落主要分布于静水区.
5) ABC曲线分析表明, 上游梯级漫湾库区静水区大型底栖动物群落年内较为稳定, 下游大朝山库区旱季较为稳定, 雨季则受中等程度的干扰, 反映了梯级水坝沿着河流纵向梯度大型底栖动物群落稳定性的累积效应.
6) 生物指数评价结果表明, 2016年漫湾库区水质(Ⅰ~Ⅱ)明显优于大朝山库区(Ⅰ~Ⅲ), 反映了梯级水坝沿着河流纵向梯度水环境受到污染的趋势, 表现出水质的累积效应.
参考文献
| Almeida D, Merino-Aguirre R, Angeler D G. 2013. Benthic invertebrate communities in regulated Mediterranean streams and least-impacted tributaries[J]. Limnologica-Ecology and Management of Inland Waters, 43(1): 34–42.DOI:10.1016/j.limno.2012.02.003 |
| Barbour M T, Gerritsen J, Snyder B D, et al. 1999. Rapid bioassessment protocols for use in streams and wadeable rivers: Periphyton, benthic macroinvertebrates and fish(2nd Edition)[R]. EPA 841-B-99-002. Washington, D. C. : U. S. Environmental Protection Agency, Office of Water |
| Chang T, Gao C, Ye C, et al. 2017. Bioassessment of sediment quality using macrofauna in the wetlands of lake buffer zone of Zhushan Bay, Taihu Lake, China[J]. Fresenius Environmental Bulletin, 26(3): 2410–2421. |
| 陈浒, 李厚琼, 吴迪, 等. 2010. 乌江梯级电站开发对大型底栖无脊椎动物群落结构和多样性的影响[J]. 长江流域资源与环境, 2010, 19(12): 1462–1470. |
| 陈丽晖, 何大明. 2000. 澜沧江-湄公河水电梯级开发的生态影响[J]. 地理学报, 2000, 55(5): 577–586.DOI:10.11821/xb200005007 |
| Chen Q, Yang Q, Li R, et al. 2013. Spring micro-distribution of macroinvertebrate in relation to hydro-environmental factors in the Lijiang River, China[J]. Journal of Hydro-environment Research, 7(2): 103–112.DOI:10.1016/j.jher.2012.03.003 |
| 董哲仁, 孙东亚, 赵进勇, 等. 2010. 河流生态系统结构功能整体性概念模型[J]. 水科学进展, 2010, 21(4): 550–559. |
| Dore J, Yu X. 2004. Yunnan hydropower expansion: update on China's energy industry reforms and the Nu, Lancang and Jinsha Hydropower dams[R]. Chiang Mai, Thailand: Chiang Mai University's Unit for Social and Environment Research, and Green Watershed |
| Grumbine R E, Xu J. 2011. Mekong hydropower development[J]. Science, 332: 178–179.DOI:10.1126/science.1200990 |
| 韩茂森, 束蕴芳. 1995. 中国淡水生物图谱[M]. 北京: 海洋出版社. |
| Helson J E, Williams D D. 2013. Development of a macroinvertebrate multimetric index for the assessment of low-land streams in the neotropics[J]. Ecological Indicators, 29: 167–178.DOI:10.1016/j.ecolind.2012.12.030 |
| 霍堂斌, 刘曼红, 姜作发, 等. 2012. 松花江干流大型底栖动物群落结构与水质生物评价[J]. 应用生态学报, 2012, 23(1): 247–254. |
| 贾志翔. 2011. 大朝山水电站库区污染成因及防治对策[J]. 环境科学导刊, 2011, 30(6): 37–39. |
| 简东, 黄道明, 常秀岭, 等. 2010. 红水河干流梯级运行后底栖动物的演替[J]. 水生态学杂志, 2010, 3(6): 12–18. |
| 李斌, 申恒伦, 张敏, 等. 2013. 香溪河流域梯级水库大型底栖动物群落变化及其与环境的关系[J]. 生态学杂志, 2013, 32(8): 2070–2076. |
| Li J, Dong S, Liu S, et al. 2013. Effects of cascading hydropower dams on the composition, biomass and biological integrity of phytoplankton assemblages in the middle Lancang-Mekong River[J]. Ecological Engineering, 60: 316–324.DOI:10.1016/j.ecoleng.2013.07.029 |
| 李晋鹏, 董世魁, 彭明春, 等. 2017. 梯级水坝运行对漫湾库区底栖动物群落结构及分布格局的影响[J]. 应用生态学报, 2017, 28(12): 4101–4108. |
| 刘其根, 查玉婷, 陈立侨, 等. 2012. 浙江分水江水库大型底栖动物群落结构及水质评价[J]. 应用生态学报, 2012, 23(5): 1377–1384. |
| 刘月英, 张文珍, 王跃先, 等. 1979. 中国经济动物志:淡水软体动物[M]. 北京: 科学出版社. |
| 罗欢, 陈骞, 吴琼, 等. 2016. 武江梯级水电站开发对底栖动物群落结构的影响[J]. 人民珠江, 2016, 37(5): 80–84. |
| Mccartney M P, Sullivan C, Acreman M C. 2000. Ecosystem impacts of large dams. In: Background Paper No. 2 Prepared for IUCN/UNEP/WCD[R]. Wallingford, UK: Center for Ecology and Hydrology |
| MorseJ C, 杨莲芳, 田立新. 1994. 中国水生昆虫及其水质监测应用[M]. 南京: 河海大学出版社. |
| 慕林青, 张海萍, 赵树旗, 等. 2018. 永定河底栖动物生物完整性指数构建与健康评价[J]. 环境科学研究, 2018, 31(4): 697–707. |
| Naiman R J, Bilby R E, Bisson P A. 2000. Riparian ecology and management in the Pacific coastal rain forest[J]. Bioscience, 50: 996–1011.DOI:10.1641/0006-3568(2000)050[0996:REAMIT]2.0.CO;2 |
| Nilsson C, Reidy C A, Dynesius M, et al. 2005. Fragment and flow regulation of the world's large river system[J]. Science, 308: 405–408.DOI:10.1126/science.1107887 |
| 欧阳慧, 邵骏, 李秀峰. 2010. 大朝山电站入库径流变化及预测研究[J]. 水电能源科学, 2010, 28(7): 6–8. |
| Petesse M L, Petrere Jr M, Spigolon R J. 2007. Adaptation of the reservoir fish assemblage index (RFAI) for assessing the Barra Bonita reservoir (S?o Paulo, Brazil)[J]. River Research and Applications, 23(6): 595–612.DOI:10.1002/(ISSN)1535-1467 |
| Perbiche-Neves G, Ferreira R A R, Nogueira M G. 2011. Phytoplankton structure in two contrasting cascade reservoirs (Paranapanema River, Southeast Brazil)[J]. Biologia, 66(6): 967–976. |
| 邵美玲, 韩新芹, 谢志才, 等. 2007. 香溪河流域梯级水库底栖动物群落比较[J]. 生态学报, 2007, 27(12): 4963–4971.DOI:10.3321/j.issn:1000-0933.2007.12.004 |
| 邵美玲, 谢志才, 叶麟, 等. 2006. 三峡水库蓄水后香溪河库湾底栖动物群落结构的变化[J]. 水生生物学报, 2006, 30(1): 64–69. |
| 孙伟胜, 顾钱洪, 董静, 等. 2015. 淮河流域板桥水库大型底栖动物群落结构及水质生物学评价[J]. 应用生态学报, 2015, 26(9): 2843–2851. |
| Tagliaferro M, Miserendino M L, Liberoff A, et al. 2013. Dams in the last large free-flowing rivers of Patagonia, the Santa Cruz River, environmental features, and macroinvertebrate community[J]. Limnologica-Ecology and Management of Inland Waters, 43(6): 500–509.DOI:10.1016/j.limno.2013.04.002 |
| 王宝强, 刘学勤, 彭增辉, 等. 2015. 三峡水库底栖动物群落结构特征及其与蓄水前资料的比较[J]. 水生生物学报, 2015, 39(5): 965–972.DOI:10.7541/2015.126 |
| 王备新, 杨莲芳. 2004. 我国东部底栖无脊椎动物主要分类单元耐污值[J]. 生态学报, 2004, 24(12): 2768–2775.DOI:10.3321/j.issn:1000-0933.2004.12.014 |
| 王超. 2017. 西南水电高坝大库梯级开发的生态保护与恢复冶研究构想[J]. 四川大学学报(工程科学版), 2017, 49(1): 1–8. |
| 王川, 李斌, 谢嗣光, 等. 2013. 澜沧江大型底栖动物群落结构及分布格局[J]. 淡水渔业, 2013, 43(1): 37–43. |
| 王德铭, 王明霞, 罗森源. 1991. 水生生物监测手册[M]. 南京: 东南大学出版社. |
| 王强, 庞旭, 王志坚, 等. 2017. 城市化对河流大型底栖动物群落的影响研究进展[J]. 生态学报, 2017, 37(18): 1–14. |
| 王忠泽, 张向明. 2000. 云南澜沧江漫湾水电站库区生态环境与生物资源[M]. 昆明: 云南科技出版社. |
| 吴东浩, 王备新, 张咏, 等. 2011. 底栖动物生物指数水质评价进展及在中国的应用前景[J]. 南京农业大学学报, 2011, 34(2): 129–134. |
| 吴涛, 李强, 张凯, 等. 2016. 底栖动物完整性指数(BIBI)在于桥水库水质评价中的应用[J]. 湖北农业科学, 2016, 55(8): 1979–1984. |
| 徐勇, 李新正, 王洪法, 等. 2016. 长江口邻近海域丰水季大型底栖动物群落特征[J]. 生物多样性, 2016, 24(7): 811–819.DOI:10.17520/biods.2016039 |
| 翟红娟, 崔保山, 胡波, 等. 2007. 纵向岭谷区不同水电梯级开发情景胁迫下的区域生态系统变化[J]. 科学通报, 2007, 52(S2): 93–100. |
| 张觉民, 何志辉. 1991. 内陆水域渔业自然资源调查手册[M]. 北京: 农业出版社. |
| 张敏, 蔡庆华, 渠晓东, 等. 2017. 三峡成库后香溪河库湾底栖动物群落演变及库湾纵向分区格局动态[J]. 生态学报, 2017, 37(13): 1–12. |
| 周红, 张志南. 2003. 大型多元统计软件PRIMER的方法原理及其在底栖群落生态学中的应用[J]. 青岛海洋大学学报(自然科学版), 2003, 33(1): 58–64. |
