 ,1,2
,1,2A new species of Scleropages (Osteoglossidae, Osteoglossomorpha) from the Eocene of Guangdong, China
ZHANG Jiang-Yong,1,2收稿日期:2019-11-19网络出版日期:2020-04-20
| 基金资助: |
Received:2019-11-19Online:2020-04-20
作者简介 About authors
zhangjiangyong@ivpp.ac.cn

摘要
记述了产于广东三水盆地华涌组的金龙鱼化石新材料,定为骨舌鱼科金龙鱼属一新种: 三水金龙鱼(Scleropages sanshuiensis sp. nov.)。新种在头部骨骼、尾骨骼、各鳍形状和位置以及具有网状鳞片等方面与中华金龙鱼(S. sinensis)和现生金龙鱼(Scleropages)极为相似,因而归入该属。在金龙鱼属的4个现生种和中华金龙鱼之中,三水金龙鱼和中华金龙鱼更为接近。但是,三水金龙鱼在下列特征上明显不同于中华金龙鱼:鼻骨上有纹饰,第三眶下骨略小于第四眶下骨,鳃盖骨后腹缘的内凹浅,胸鳍往后延伸到腹鳍起点,躯椎上的神经棘不愈合,椎体横突长。三水金龙鱼与现生的金龙鱼也有相似之处,如鼻骨有纹饰、椎体横突长。同样,中华金龙鱼也有与现生金龙鱼相似的地方,如第三眶下骨略大于第四眶下骨、大多数躯椎上的神经棘愈合。尚不清楚三水金龙鱼和中华金龙鱼中哪一个与现生种更接近,这两个化石种可能处于相同的进化水平。中生代骨舌鱼类主要发现于中国,始新世骨舌鱼类化石在中国的发现也在逐渐增多,金龙鱼属(Scleropages)和骨舌鱼属(Osteoglossum)的共同祖先在始新世以前可能生存于东亚,并随后扩散到澳洲和世界其他地方。华涌组的骨舌鱼类和湖北松滋洋溪组的可以直接比较,因此,本项研究支持华涌组的时代为早始新世的观点。
关键词:
Abstract
A new species of Scleropages, an extant genus of the Osteoglossidae, is described from the Lower Eocene Huayong Formation in the Sanshui Basin, Guangdong, China. The new species, Scleropages sanshuiensis sp. nov., was attributed to Scleropages because it possesses the diagnostic features of the genus in skull bones, caudal skeleton, the shape and position of fins, and reticulate scales. Compared with the four extant and one fossil species of Scleropages, the new species is most similar to the fossil one, S. sinensis. However, S. sanshuiensis is different from S. sinensis in many aspects: the nasal is ornamented; the third infraorbital is slightly smaller than the fourth; the posteroventral margin of the opercle is slightly concave; the pectoral fin extends just to the beginning of the pelvic fin; the distal ends of the neural arches on abdominal centra are not fused; the parapophyses are much longer than in S. sinensis. Scleropages sanshuiensis is closer to S. sinensis but it shares two characters with the living species (ornamented nasal and long parapophyses) while S. sinensis shares two characters with the extant ones too (the third infraorbital slightly larger than the fourth and the distal ends of the neural arches on most abdominal centra fused). It is not clear which one of the two fossil fishes is more closely related to the living fishes. The two fossils are probably in the same evolutionary level. In addition to the rich Mesozoic osteoglossomorphs, many Eocene ones were also found from China. The common ancestor of Scleropages and Osteoglossum may live before the Eocene in East Asia and then dispersed to Australia and other places of the World. Huayong Formation and Yangxi Formation, Songzi, Hubei are comparable and possibly deposited contemporaneously. Therefore, this work supports the view that the Huayong Formation is of Early Eocene.
Keywords:
PDF (5725KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
张江永. 广东始新世金龙鱼(Scleropages, Osteoglossomorpha)属一新种. 古脊椎动物学报[J], 2020, 58(2): 100-199 DOI:10.19615/j.cnki.1000-3118.191213
ZHANG Jiang-Yong.
1 Introduction
Scleropages (Asian arowanas) are superstars in an aquarium for their ornate coloration in some color variants. The fans from Thailand, China (Taiwan and Hong Kong), and Japan believe that the arowanas bring them good luck and fortune. The arowana fishes exhibit a trans-marine distribution, living on both sides of the Wallace’s line. Among the four extant species of the genus, S. formosus (Müller and Schlegel, 1844) distributed in Sumatra, Kalimantan, Peninsular Malaysia, Thailand, and Cambodia, S. inscriptus (Roberts, 2012) only in Myanmar, S. jardinii (Saville-Kent, 1892) and S. leichardti (Günther, 1864) in Australia and New Guinea. This distribution of Arowanas has attracted the interests of some molecular biologists (Kumazawa and Nishida, 2000; Lavoué and Sullivan, 2004) who reconstructed the biogeography based on a molecular phylogeny and molecular-clock.Fossil remains of scales, otoliths and fragments of bones of Scleropages have long been known from the Maastrichtian of India (Hora, 1938; Rana, 1988; Kumar et al., 2005; Nolf et al., 2008), the Maastrichtian/Late Paleocene of Africa (Taverne, 2009), the Paleocene of Europe (Taverne et al., 2007), the Eocene of Sumatra (Sanders, 1934; Forey and Hilton, 2010), the Oligocene of Australia (Hills, 1934, 1943; Unmack, 2001). Taverne et al. (2007) proposed that a fragment of a toothed jaw and some squamules of an undetermined osteoglossid from the continental Campanian (Late Cretaceous) of Champ-Garimond in southern France (Sigé et al., 1997) may represent as the earliest fossil record of the genus Scleropages. Recently, a complete fossil Scleropages was described from the Lower Eocene strata in Xiangxiang, Hunan Province and Songzi, Hubei Province, China (Zhang and Wilson, 2017) that made it possible to carry out a detailed comparative analysis between the fossil and extant species for the first time. Here the author reports on the second fossil species of Scleropages from the Lower Eocene Huayong Formation in the Sanshui Basin of Guangdong Province, China.
The Sanshui Basin is located in the Northwest of the Pearl River Delta. Most of the area belongs to Sanshui, Nanhai, Chancheng and Shunde Districts of Foshan City, covering an area of about 3300 km2. The large regions of the surface of the Sanshui Basin are covered by Neogene loose sediments and vegetation. Paleogene strata are only sporadically exposed and are divided into four formations from bottom to top: Xinzhuangcun, Buxin, Baoyue and Huayong Formations (Zhang et al., 2008). The lithology of the lower part of the Huayong Formation is the interbedding of volcanic and sedimentary rocks, different from the underlying the Baoyue Formation that has a large suit of volcanic rock as its marker. The upper part of the Huayong Formation is composed of conglomerate, sandy gravel with brownish red siltstone, fine sandstone, dark gray silty mudstone and scattered purple gray basalt, with a thickness of nearly 1000 m.
The Paleogene fish fossils in the Sanshui Basin are abundant and widely distributed. Fishes were mainly collected from the Buxin Formation. Only one genus, Tungtingichthys, is from individual drill holes of the Xinzhuangcun Formation and scales and fragments of bones are from the Baoyue and Huayong Formations. Wang et al. (1981) described three families, 8 genera, 15 species, including 10 new species from the Buxin Formation except the Leuciscinae gen. et sp. indet.1 and the Leuciscinae gen. et sp. indet. 2 from the Huayong Formation. However, most of their specimens are not well preserved and need to be re-identified. Two new fish faunas have been discovered since 2006 (Zhang et al., 2009): 1) Tungtingichthys fauna can be seen from the upper part of the Xinzhuangcun Formation to the bottom of the Buxin Formation. The fossil fishes are rich in a set of hard oil shales, which are well-preserved but poor in diversity. This fauna is characterized by the occurrence of a large number of Tungtingichthys and occasionally cyprinids and bagrids. 2) Osteoglossid fauna can be seen in the black tuffaceous shale of the Huayong Formation in the central basin. The fossil fishes from the second fauna are rich in both individual number and taxonomy, including osteoglossids, Jianghanichthys, Aoria lacus, Tungtingichthys and amiiforms. Some members of this fauna are also seen in the sandstone of upper part of the Huayong Formation including Scleropages. In addition to fishes, other fossil vertebrates were also found in the Huayong Formation recently, including Sanshuibatrachus sinensis (Anura, Pelobatoidea. Wang et al., 2017), Sanshuiornis zhangi (Aves, cf. Ciconiiformes. Wang et al., 2012), Meridiolophus expansus (Mammalia, Perissodactyla. Bai et al., 2014), chelonia, crocodilia and lizards (Zhang et al., 2009). Ostracodes, gastropods, spores and pollen, charophytes and plants were also seen in this formation.
2 Material and methods
The specimens studied are deposited in the collection of Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences (IVPP). The comparative materials of Scleropages sinensis (IVPP V 12749.1-8, V 12750, V 13672.1-3, V 26536.1-2), dried skeletons of the extant species of the genus (S. formosus, IVPP V OP 80; S. leichardti, V OP 81; S. jardinii OP 303-307), and a dried skeleton of Osteoglossum (OV 2712) are also deposited in IVPP. Scleropages inscriptus is not available at present.Some impression fossils were scanned with a flatbed Epson Scanner and then the photos were flipped vertically, making the impression like the body fossil visually.
3 Systematic paleontology
Teleostei Müller, 1846Osteoglossomorpha Greenwood et al., 1966
Osteoglossiformes Regan, 1909
Osteoglossidae Bonaparte, 1832
Scleropages Günther, 1864
Scleropages sanshuiensis sp. nov.
(Figs. 1-9)
Fig. 1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1Scleropages sanshuiensis sp. nov. from Sanshui Basin, Guangdong
A. holotype (IVPP V 26533.1) in right lateral view; B. a nearly complete fish (V 26534.1) in left lateral view; C. a skeleton missing head (V 26533.5) in left lateral view All scale bars in figures of this paper equal 1 cm
Fig. 2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Skulls of Scleropages sanshuiensis sp. nov., impressions were scanned with a flatbed Epson Scanner and then flipped vertically
A.IVPP V 26533.6; B. IVPP V 26533.8
Fig. 3
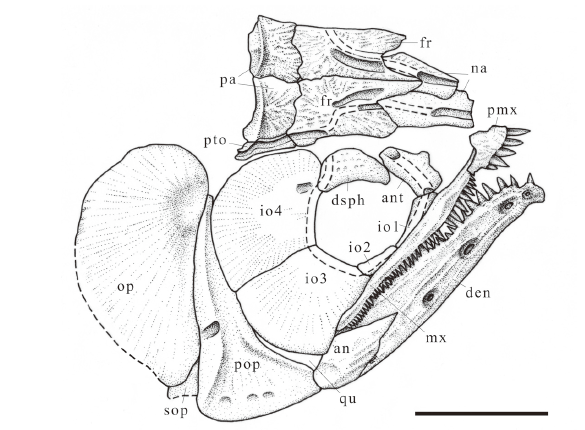 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3Sketch of Scleropages sanshuiensis sp. nov., skull as preserved in holotype, right lateral view
Abbreviations: an. angular; ant. antorbital; den. dentary; dsph. dermosphenoid; fr. frontal; io1-4. first to fourth infraorbitals; mx. maxilla; na. nasal; op. opercle; pa. parietal; pmx. premaxilla; pop. preopercle; pto. dermopterotic; qu. quadrate; sop. subopercle
Fig. 4
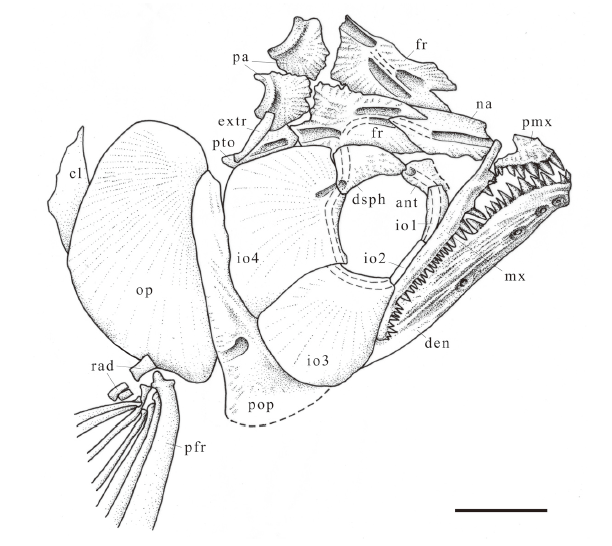 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 4Sketch of Scleropages sanshuiensis sp. nov., skull based on a scanned and flipped photo of IVPP V 26533.6, right lateral view
Abbreviations: cl. cleithrum; extr. extrascapular; pfr. pectoral fin ray; rad. radials; other abbreviations see
Fig. 5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 5Skull bones of Scleropages sanshuiensis sp. nov. and S. sinensis
A. opercle of S. sanshuiensis sp. nov. (IVPP V 26533.19A); B. opercle of S. sinensis (V 26536.2.); C. skull bones of S. sanshuiensis sp. nov. (V 26533.19A, impression scanned with a flatbed Epson Scanner and then flipped vertically)
Fig. 6
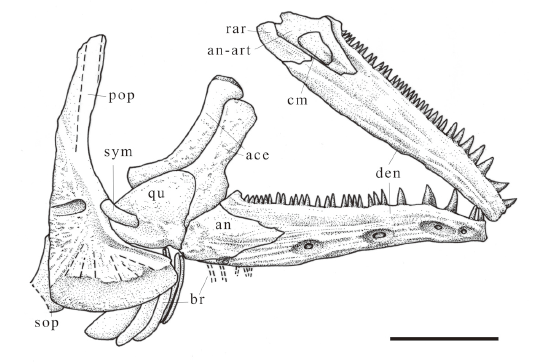 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 6Sketch of Scleropages sanshuiensis sp. nov., skull bones based on a scanned and flipped photo of IVPP V 26533.19A
Abbreviations: ace. anterior ceratohyal; an-art. angulo-articular; br. branchiostegals; cm. coronomeckelian; rar. retroarticular; sym. symplectic; other abbreviations see
Fig. 7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 7Scleropages sanshuiensis sp. nov. from Sanshui Basin, Guangdong
A. parapophysis (IVPP V 26534.5); B. a caudal skeleton (IVPP V 26534.1)
Fig. 8
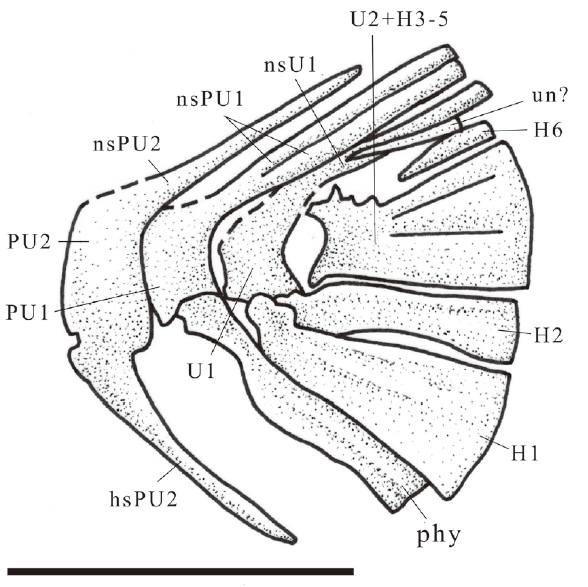 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 8Sketch of Scleropages sanshuiensis sp. nov., caudal skeleton, IVPP V 26534.1
Abbreviations: H1-6. hypurals 1-6; hsPU2. haemal spine on PU2; nsPU1, 2. neural spines on PU1, 2; nsU1. neural spine on U1; phy. parhypural; PU1, 2. preural vertebrae 1, 2; U1, 2. ural centrum 1, 2; un. uroneural
Fig. 9
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 9Scales of Scleropages sanshuiensis sp. nov. from Sanshui Basin, Guangdong
A. IVPP V 26533.23, B. IVPP V 26533.22
Etymology The specific name refers to the Sanshui Basin where the specimens were found.
Holotype IVPP V 26533.1, a nearly complete skeleton.
Referred specimens IVPP V 26533.2-23, V 26534.1-10.
Locality and horizon Specimens V 26533.1-23 are from Niuweigang, Zidong, Nanzhuang Town, Chancheng District, Foshan City; Lower part of the Huayong Formaion, Lower Eocene. Specimens V 26534.1-10 are from Shicheng Middle School, Shishan Town, Nanhai District, Foshan City; upper part of the Huayong Formaion, Lower Eocene.
Diagnosis A fossil species of Scleropages different from S. sinensis and extant species in: sensory canal of nasal in a groove on the anterior half but in a tube of the posterior half, third infraorbital slightly smaller than fourth, posteroventral margin of opercle concave slightly, sensory canal opening in a groove along the whole dermopterotic, last branchiostegal ray is much wider than the penultimate, pectoral fin extending just to the beginning of pelvic fin, distal ends of neural arches on abdominal centra not fused, parapophyses much longer than in S. sinensis but not as long as in extant species, hypural 2 unconnected with ural centrum 1.
4 Description
The body of the fish is fusiform (Fig. 1). The standard length of the holotype is 116 mm, 3.5 times of the length of the head. The standard length of the largest specimen (V 26534.1) is 182 mm, 3.3 times of the head length.Cranium The nasals (Figs. 2-4) are large and sutured in the midline along the anterior two thirds of their length, but are separated by the tapered frontals posteriorly as in S. sinensis. There is a notch at the anterior medial corner of the bone probably to receive the dermethmoid. This notch can be seen in S. sinensis (relatively small) but not seen in the extant species (IVPP dried skeleton). The nasals are ornamented like the condition in extant species (IVPP dried skeleton) but unlike that in S. sinensis. The sensory canal is quite different from other species of the genus. A groove is exposed in the anterior half of the nasal and goes into a tube in the posterior half of the bone.
The canal is exposed in a groove along the whole length of the nasal in S. sinensis and S. leichardti, but is presented as a tube in the nasal in S. formosus.
The frontal (Figs. 3, 4) is similar in shape and ornamentation to other species of Scleropages. It is long and subrectangular, with an anterior embayment for the reception of the nasal, and a posterior sinuous suture with its opposite member. The sensory canal is enclosed in bone for the middle third of its length, while it is exposed in grooves for the anterior and posterior thirds of its length in the frontal, similar to that in S. sinensis and S. leichardti but unlike in S. formosus, in which the canal is enclosed in bone to, or almost to, its entry into the nasal.
The parietal (Figs. 3, 4) is subrectangular and sutures with its opposite at the midline and with the pterotic laterally. The bone is sculptured except for the posterior quarter of the lower level. The bone is slightly wider than it is long, similar to that of S. formosus but unlike the ratio seen in S. leichardti.
The dermopterotic (Figs. 3, 4, V 26533.8) is a little larger and thicker than it is in extant species of Scleropages but smaller than in S. sinensis. The bone seems not sculptured and bears the temporal sensory canal in an open groove, different from that in other species of the genus where only the anterior half of the canal is in an open groove (the holotype of S. sinensis, IVPP dried skeleton).
The extrascapular (Fig. 4) is a tubular bone. The epiotic, supratemporal and supraoccipital are not visible in the available specimens.
The orbital portion of the parasphenoid is toothless, moderately broad, and parallel-sided (V 26533.8). The parasphenoid is nearly complete in V 26533.14, with strong basipterygoid processes and a small tooth patch at the base of the process. The bone joins with the basioccipital posteriorly. The vomer is seen in V 26533.14 in lateral view, with four teeth, one much larger than the other three.
The circumorbital series (Figs. 2-4, V 26533.8) is composed of six bones: an antorbital, four infraorbitals and a dermosphenotic. The supraorbital is absent. The antorbital, infraorbitals and the dermosphenotic are all prominently sculptured.
The antorbital is well-preserved in V 26533.8 and is polygonal, making contact with the dermopterotic posterodorsally, the nasal and frontal dorsomedially, and the first infraorbital ventrally. The concave anterior and orbital margins are free. Dorsally the circumorbital sensory canal enters the antorbital via a short and small groove, and then passes through the bone in a tube, entering the first infraorbital where a pore communicates with the exterior as in S. sinensis. In the extant species of the genus (IVPP dried skeleton) and Osteoglossum (IVPP OV 2712), the canal is completely enclosed in bone. The antorbital of the two fossil species is relatively larger than in the extant species of Scleropages and in Osteoglossum.
The first and the second infraorbitals are narrow and tubular. The first is slightly expanded, longer, and more ornamented than the second.
The two posterior infraorbitals (third and fourth) are very large as in S. sinensis but do not quite reach the size of those in extant Scleropages and Osteoglossum, in which they extend posteriorly to the articulation of the opercle, completely concealing the dorsal end of the preopercle. In both fossil species of Scleropages there is a narrow gap through which the dorsal limb of the preopercle may be seen. The third infraorbital is slightly smaller than the fourth while it is slightly larger than the fourth in other species of the genus including S. sinensis. In Osteoglossum the lower one is much larger than the upper. The infraorbital sensory canal is carried in a tube near the orbital margin of both posterior infraorbitals. Near the anterodorsal corner of the fourth infraorbital where the sensory canal enters the dermosphenotic, there is a short backward groove. This groove is also seen in other species of Scleropages and Osteoglossum.
The dermosphenotic (Figs. 2-4) is a nearly triangular bone, large, thick, and sculptured. A pore of the sensory canal is seen at the posteroventral corner of the bone. This bone is larger in S. sanshuiensis sp. nov. and S. sinensis than in the two extant species of the genus and in Osteoglossum.
Jaws The premaxilla (Figs. 3, 4) is small and sculptured. There is a small ascending process in its anterior half. Seven teeth are seen on the specimens V 26533.1 and V 26533.6, but eight on specimen V 26533.10. The anterior four teeth are much larger than the posterior ones while the number of the large teeth on the bone in S. sinensis is three. The number of teeth on the premaxilla of S. sinensis is seven, but in extant Scleropages the number has been counted differently, 4-5 by Ridewood (1905), 3-5 by Kershaw (1976), and 11 by Taverne (1977). In the IVPP dried specimens of extant Scleropages this number is 4-8.
The maxilla (Figs. 2-4) is long and slender, and takes an angle of about 45° with the long axis of the fish when the mouth is closed as it is in S. sinensis. It extends posteriorly nearly to the level of the mandibular articulation and ends well behind the posterior margin of the orbit. Ornament is most prominent at the anterior and posterior ends. There is no supramaxilla.
The maxilla bears 40 conical teeth in the holotype and V 26533.6, a resemblance in number with S. sinensis and S. formosus and a difference from S. leichardti, in which the tooth number is about 35 (IVPP dried skeletons). The teeth decrease in size steadily from anterior to posterior, with 4-5 teeth much larger than the others.
The mandible (Figs. 5C, 6) is long and lacks a distinct coronoid process, consisting of dentary, angulo-articular, and retroarticular. The dentary forms the great majority of the length of the mandible. Anteriorly, the dentary curves inwards to form the shallow symphysis. 33 teeth are counted on the oral margin of the dentary in V 26533.19A, with the anterior five or six much larger than the posterior ones. Dense long ridges are ornamented on the lower part of the lateral surface of the dentary.
The angulo-articular (Figs. 5C, 6) is short, and articulates with the quadrate as seen in lateral view. The anterior part of the angulo-articular is thin and pointed overlapping the recess on the internal surface of the dentary. Posteriorly the bone is thick and deeply sculptured. The articular facet for the quadrate is formed by the bone alone and lies in the upper part of the posterior end of the bone. The facet clearly showing in V 26534.5 is a little deeply excavated. The coronomeckelian (Figs. 5C, 6) can be seen on the medial surface of the angulo-articular in V 26533.19A. The coronomeckelian is also visible on the medial surface of the mandible of S. sinensis (V 26536.2).
The retroarticular (Figs. 5C, 6) is very small and applies to the postero-ventral surface of the angulo-articular.
The mandibular sensory canal extends the length of the dentary and angulo-articular within a canal, with five pores located near the ventral margin of the dentary.
Palato-quadrate arch The palato-ectopterygoids are seen in V 26533.10, in which there are small teeth on the lateral margins and a patch of much smaller teeth behind the margin.
Two detached entopterygoids are preserved in specimens V 26533.17-18, one in lateral view and the other in medial. The bone is triangular, with a row of 20 large conical teeth existing on the medial edge, and fine denticles covering the remainder of the surface. The metapterygoid is not visible. The whole quadrate (Figs. 5C, 6) can be seen in V 26533.19A, B. It is a small fan-shaped bone, with the postero-ventral process a little short and the incision for the symplectic somewhat shallow. A short, rod-like symplectic is visible in V 26533.19A, which inserts into the incision of the quadrate.
Hyoid arch and branchiostegals The hyomandibula (V 26534.5) is vertical and articulates with the cranium by one head. The opercular process is strong and lies in the upper third of the bone. Anteriorly, the bone has a broad wing. Two foramina opened in the external surface of the bone, one at the upper third and the other near the lower end.
The anterior ceratohyal (Figs. 5C, 6) is seen in V 26533.19A and V 26534.4. Its proximal part is rod-like while the distal part is nearly rectangular. The hypohyal and the basihyal are not visible. A small urohyal can be seen in V 26534.4. There are 10 slender, acinaciform branchiostegal rays in V 26533.19A, with the last three gradually broadened. The last branchiostegal ray is much wider than the penultimate, differing from what is seen in other species of the genus.
Opercular series The preopercle is similar to that in S. sinensis and the extant species of Scleropages. The upper limb is not completely covered by the posterior infraorbitals as it is in the extant species of the genus (Taverne, 1977:fig. 71). The dorsal limb is slightly more than twice the length of the ventral limb. The latter is bluntly rounded anteroventrally. The posteroventral angle of the preopercle is not produced posteriorly to a point. Zhang and Wilson (2017) described that the posteroventral angle of S. sinensis is produced posteriorly to a point just like that in Chauliopareion and Singida (Murray and Wilson 2005). On the newly found specimens (V 26536.1-2) of S. sinensis from Songzi, the type locality of the species, the posteroventral angle of the preopercle is complete and does not show this feature.
The preopercular sensory canal is similar to that of S. sinensis and is open ventrally beneath a long, horizontal shelf. The surface ventral to the shelf is smooth but with some granules near the ventral margin for the preopercle (V 26533.19A). The sensory canal is enclosed in bone on the anterior edge, but opens ventrally with five pores beneath the shelf and posteriorly with a large pore at about the lower third height of the preopercle as that in S. sinensis and the extant species of the genus.
The opercle (Fig. 5A, V 26533.10, V 26533.19A) is nearly semicircular. The ratio depth /width of the bone is 1.89 in V 26533.10 and 1.98 in V 26533.19A. The ratio of that in S. sinensis is 1.80 in the holotype and 1.91 in V 26536.2. The ratio of extant species of the genus is of two opposing extremes, 2.25 in S. formosus and 1.72 in S. leichaditi. The posteroventral margin of the opercle is concave slightly (Fig. 5A), whereas it is concave sharply in S. sinensis (Fig. 5B). In extant species of genus, by contrast, the margin is not concave. The opercle is prominently sculptured with radial ornamentation on the lateral surface except for its anterior margin and dorsal extremity. The medial surface is smooth (V 26533.19A). The subopercle is only partially shown in V 26533.1 and V 26533.8. The interopercle is not visible.
Appendicular skeleton The upper part of the pectoral girdle is seen in V 26533.10 and V 26534.5. The posttemporal is not visible. The supracleithrum is long and thickened along the anterior margin. The anterior margin is a little straight but the posterior margin is rounded. A small postcleithrum is present and lies medial to the junction between the supracleithrum and cleithrum. The cleithrum shows only its upper part, terminating dorsally in a long, rod-like process. The lower part of the girdle can be seen in V 26533.11, but the coracoid, scapula, and mesocoracoid have not been seen. Three proximal pectoral radials are visible in V 26533.6, one large and the rest two small (Fig. 4).
The pectoral fin (V 26533.2, 11) is very long and extends just to the beginning of the pelvic fin, as compared with that in S. sinensis in which it extends well behind the beginning of the pelvic fin. In the extant species of Scleropages it does not reach the beginning of the pelvic fin. There are seven pectoral fin rays (V 26533.1, 11). This number is also seven in S. sinensis and S. formosus but eight in S. leichardti (IVPP dried skeletons). All rays are branched and segmented except the first one which is exceptionally thick and unbranched, though segmented. A claw-shaped bone is showing in V 26534.2. The same bone is also found adjacent to the base of the smallest ray of S. sinensis.
The pelvic girdle and fins are very small (V 26533.11). Each pelvic fin originates in about the middle length between the pectoral and anal fins, different from that in S. sinensis in which the pelvic fin originates slightly closer to the anal fin than to pectoral fin. The pelvic girdle (V 26533.11) is short and flat. There are six pelvic fin rays, all branched but the first, a condition agreeing with that of S. sinensis and S. leichardti but differing from S. formosus, which has five fin rays.
Dorsal and anal fins The dorsal and anal fins are rounded in outline and located posteriorly (V 26533.4, 5). The dorsal fin is small and originates posterior to the origin of anal fin, opposite the middle of the anal fin. There are three short procurrent dorsal rays, one long segemented but not branched ray and 11 branched rays, for a total of 12 principal rays in V 26533.4-5. The dorsal pterygiophores can be counted only in V 26533.4, about 14 in number. The anal fin is much larger than the dorsal fin, with three small, unsegmented procurrent rays and 22 principal rays. The anal pterygiophores cannot be counted.
Vertebral column and caudal fin There are 48 vertebrae in V 26533.5, of which about 22 are abdominal and 26 are caudal including the two ural centra. This number is much less than in extant osteoglossids and agrees with that of S. sinensis and early osteoglossomorphs such as Kuntulunia and Xixiaichthys (Zhang, 1998, 2004). The first three centra are covered by the opercle. The centra are slightly deeper than long. The neural spines are paired on abdominal centra (V 26533.2), in which some paired spines may fuse at the distal end, the case different from that of S. sinensis in which only the first four neural spines are fused.
The parapophyses (Fig. 7A, V 26534.5) are much longer than in S. sinensis and most early osteoglossomorphs (Zhang, 1998, 2004; Zhang and Jin, 1999).
There are 22 pairs of pleural ribs (V 26533.2), which extend to the ventral margin of the trunk, except for the last pair, which is only about half the length of the more anterior ones.
The epineurals are not visible. Some long, slender supraneurals are seen in specimen V 26533.2.
The caudal skeletons (Figs. 7B, 8) are very similar to those of S. sinensis and the extant species of Scleropages with certain disparities. These are best preserved in the specimen V 26534.1. Three haemal spines in S. sanshuiensis are lengthened to support the caudal fin rays. The first preural centrum bears two complete neural spines as in S. leichardti, contrasting single in S. sinensis and S. formosus.
The first ural centrum (U1) has a complete neural spine. The second ural centrum (U2) is fused with the proximal ends of hypurals 3-5 (H3-5). There are six hypurals. H1 is very deep and joints with U1 proximally. H2 is less than half the width of H1 and is probably unconnected with U1. Hypurals 3 through 5 are fused proximally and fit tightly together distally. A free rod-like bone dorsal to hypurals 3-5 is probably the sixth hypural. Just above this bone, a similarly shaped bone is interpreted here as an uroneural. A greatly enlarged first hypural is otherwise present only in S. sinensis among this genus. This hypural is as deep as the first two (of three lower) hypurals in S. leichardti.
The caudal fin has a rounded posterior profile. There are 16 principal rays, as in S. sinensis and S. leichardti, versus 14 in S. formosus. The first and the last principal rays are unbranched and almost as long as the remaining rays just as in S. sinensis, whereas in living species of Scleropages and Osteoglossum, the upper and lower rays are only half the length of the innermost ones. One or two procurrent rays are present anterior to the principal rays.
Squamation The scales are large (Fig. 9), cycloid or oval, and exhibit the reticulate pattern, involving small units called squamules, typical of osteoglossids. The external surface of the scale shows circuli in the basal portion and granular ornamentation in the apical area. The squamules (Gayet and Meunier, 1983) are rhombic, polygonal, or irregular in shape, usually larger in basal portion than in apical. The mesial surface of squamules in the apical area bears rounded, raised tubercles, each of which has a minute transversal-pore at its center while that in the basal portion is smooth, but with large pores in several squamules.
The lateral line (Fig. 9B, V 26533.22,) runs just below the vertebral column, but the number of scales along the lateral line cannot be counted. The size of the scales in S. sanshuiensis is about equal to that of S. sinensis and therefore, the same number of lateral line scales of 24 (Zhang and Wilson, 2017) is expected.
5 Discussion
The materials from the Sanshui Basin, Guangdong province of China very much resemble Scleropages in skull bones, caudal skeleton, the shape and position of fins, and reticulate scales. Therefore, it is attributed to the genus.Among the four extant and one fossil species of Scleropages, the new fish is very similar to the fossil one, S. sinensis in: there is a notch at the anterior medial corner of the nasal, the teeth on anterior oral margin are much larger than those on the posterior; the upper limb of the preopercle is not completely covered by posterior infraorbitals; the posteroventral margin of the opercle is concave; the pectoral fin is very long; a claw-shaped bone is present adjacent to the base of the smallest ray of the pectoral fin; the ray number of all the fins is equal respectively; vertebrae is same in number; hypural 1 is very deep; the number of lateral line scales is probably same.
Although S. sanshuiensis shares some similarities with S. sinensis, it is different from the latter in many aspects: the nasal is ornamented; the sensory canal of the nasal is exposed in a groove on the anterior half of the bone but in a tube in the posterior half unlike in the groove of whole length of the nasal of S. sinensis; the temporal sensory canal of the dermopterotic is in an open groove, different from that in other species of the genus where only the anterior half of the canal is in an open groove; dorsally the circumorbital sensory canal enters the antorbital via a small groove rather than a broad groove in S. sinensis; the third infraorbital is slightly smaller than the fourth while it is slightly larger than the fourth in other species of the genus including S. sinensis; the posteroventral margin of the opercle is concave slightly vs concave sharply in S. sinensis; the pectoral fin extends just to the beginning of the pelvic fin rather than extending well behind the beginning of the pelvic fin in S. sinensis; the distal ends of the neural arches on abdominal centra are not fused unlike the condition in S. sinensis that only the anterior four neural arches are not fused; the parapophyses are much longer than in S. sinensis; H2 is probably unconnected with U1 vs either articulates with or fused to U1 in S. sinensis and extant species of Scleropages.
Taverne et al. (2007) described Scleropages sp. based on some bony remains, otoliths and squamules from the continental Paleocene of Hainin (Mons Basin, Belgium). The bone remains include a right premaxilla, an incomplete right maxilla, a fragment of a right entopterygoid, a fragment of a left palato-ectopterygoid and an abdominal vertebra. The otoliths are not seen in S. sanshuiensis, and the squamules are nearly the same as in osteoglossids. The bone remains are difficult to compare with those of S. sanshuiensis except for the premaxilla on which the anterior teeth of the bone are very large in S. sanshuiensis and S. sinensis but small in Scleropages sp. of Hainin.
Based on these comparisons, a new species, Scleropages sanshuiensis is established.
Sharing many characters, the two fossil species of Scleropages are clearly more closely related to each other than either of them to the extant species. As to the relationship between the two fossil species and the extant ones, S. sanshuiensis shares two characters with living species (ornamented nasal and long parapophyses) and S. sinensis shares two characters with living species (the third infraorbital is slightly larger than the fourth and the distal ends of the neural arches on most vertebrae fused). Therefore, it is not clear which one of the two fossil fishes is more closely related to the living fishes. The two fossils are probably in the same evolutionary level.
Sexual dimorphism may exist in S. sinensis (Zhang and Wilson, 2017), with males have a slimmer and shallower body depth while females have a more rounded body. This is not seen in the available specimens of S. sanshuiensis.
The biogeographic history of the Osteoglossomorpha was undoubtedly complex. The remarkable diversity of basal osteoglossomorphs and primitive osteoglossiforms in SE Asia and the centre of modern biodiversity in Africa are two phenomena that, on present knowledge, seem inexplicable by simple hypotheses of centres of origin, or of Pangaean or Gondwanan continental breakup (Wilson and Murray, 2008). Biogeography of Scleropages seems simple, but in fact, it is not.
Kumazawa and Nishida (2000) first tested biogeographical hypotheses for Scleropages using a molecular phylogenetic time-scale and they concluded that the divergence time between Asian arowana (S. formosus) and Australia arowana (S. leichardti and S. jardinii) is about 138 million years, which is close to or slightly older than the probable time of the India-Madagascan separation from Gondwanaland (120-130 Ma, Smith et al., 1994). They consequently argued that the Asian arowana originated on a part of Gondwanaland and was carried to Eurasia by the Indian subcontinent. However, Lavoué (2015) reported an age CI of Scleropages ranging from 79.9 to 101.4 Ma, which is significantly younger than the age ((138 ± 18) Ma) inferred by Kumazawa and Nishida (2000) and even younger than 115.0 Ma (latest possible age for a direct connection of the Indian subcontinent to Australia-Antarctica) (Gibbons et al., 2013; Scotese, 2014). Therefore, Lavoué’s result rejects the Gondwanan origin hypothesis to explain the distribution of Scleropages. More recently, Cioffi et al. (2019) reported that a time-calibrated phylogenetic tree revealed that Osteoglossum and Scleropages divergence occurred approximately 50 million years ago (Mya), at the time of the final separation of Australia and South America (with Antarctica). Asian S. formosus and Australian Scleropages diverged about 35.5 Mya, substantially after the latest terrestrial connection between Australia and Southeast Asia through the Indian plate movement.
Since a vicariant event is not available to explain the transmarine distribution of Scleropages, a marine dispersal between Australia and Asian across Wallace’s Line was suggested (Bonde, 2008; Wilson and Murray, 2008; Hilton and Lavoué, 2018; Capobianco and Friedman, 2019; Cioffi et al., 2019). Although the authors mentioned above support the marine dispersal hypothesis, none of them pointed out a direction for the dispersal. Is it from south to north or just the opposite? Traditionally, it was supposed that the ancestor of Scleropages lived in Australia during the Oligocene and then moved to East Asia by a marine dispersal event (Nelson, 1969; Cracraft, 1974). Eocene S. sanshuiensis and S. sinensis from China predate this hypothetical dispersal and therefore, the direction of the dispersal need to be reconsidered.
Early osteoglossomorphs have mostly been recovered from China (Zhang, 1998, 2004), including a Late Jurassic Paralycoptera (Tse et al. 2015), one of the oldest osteoglossoids (Xu and Chang, 2009), from Hong Kong, and consequently, Greenwood (1970) and Chang and Chou (1976) supposed that East Asia might be the ancestral of osteoglossomorphs. In addition to these Mesozoic ones, the Eocene osteoglossomorphs were also found from China, such as Sinoglossus lushanensis (Su, 1986), Eohiodon shuyangensis (Shen, 1989), Phareodus songziensis (Zhang, 2003), Scleropages sinensis (Zhang and Wilson, 2017) and S. sanshuiensis It is reasonable to suppose that the common ancestor of Scleropages and Osteoglossum lived before the Eocene in East Asia and then dispersed to Australia and other places in the world.
To firmly support the marine dispersal hypothesis of Scleropages, a marine fossil of the genus may well be needed. The Sanshui Basin, where S. sanshuiensis was found, was probably connected with the sea at the time when the Paleocene Buxin Formation was deposited (Hou et al., 2007). It is difficult but not impossible to find marine fossil Scleropages in the Sanshui Basin.
About the geological time of the Huayong Formation in the Sanshui Basin has not formed a unified view. It was considered to be Late Eocene by the majority of researchers (Xu et al., 2000; Li et al., 2005; Hou et al., 2007) and to be Early Eocene (Zhang et al., 2008) or Oligocene (Wei et al., 2018) by a few other researchers. The isotopic age of the volcanic rocks in the Huayong Formation is 43-55 Ma, but the K-Ar age of the trachyte is (51.5±2.6) Ma (Zhang et al., 2008).
Scleropages sanshuiensis from the Huayong Formation of the Sanshui Basin and S. sinensis from the Yangxi Formation of Songzi are very close to each other and they are probably in same evolutionary level. Therefore, the Huayong and Yangxi Formations are comparable and possibly deposited contemporaneously. At present, the Early Eocene age of the Yanxi Formation is not disputed (Editorial Committee of Stratigraphical Lexicon of China, 1999; Wang, 2015). In view of this, this work supports the view that the Huayong Formation is of the Early Eocene.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 5]
[本文引用: 3]
[本文引用: 2]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}