 ,1, 邱占祥1, 李绿洲2,3
,1, 邱占祥1, 李绿洲2,3A Late Miocene Huerzelerimys (Rodentia: Muridae) skull from Hezheng, Gansu, China
WANG Ban-Yue,1, QIU Zhan-Xiang1, LI Lü-Zhou2,3收稿日期:2019-12-29网络出版日期:2020-04-20
| 基金资助: |
Received:2019-12-29Online:2020-04-20
作者简介 About authors
wangbanyue@ivpp.ac.cn

摘要
描述了甘肃临夏盆地晚中新世柳树组中采集到的一件鼠亚科具下颌骨的头骨和数节颈椎化石(IVPP V 16288)。该头骨代表许氏鼠的一新种,被命名为亚洲许氏鼠(Huerzelerimys asiaticus sp. nov.)。这是许氏鼠头骨的首次发现。其主要特征是:个体较小;头骨较宽短,具较宽短的吻部和较窄的眶间区;有较发达的前颌骨侧背嵴;两额嵴弱,近于平行;门齿孔长,向后延伸至M1前齿根舌侧;后颚孔位于M2舌侧;硬颚后缘位于M3之后;无翼间孔;具骨质的翼蝶管和膨大的听泡;内颈动脉孔的位置靠近基结节;下颌骨水平支较低,齿隙凹入较深;咬肌嵴伸至m1前缘下方;颏孔位于m1的前下方,咬肌嵴前方,靠近齿隙;I2为垂直伸型;M1的t1位置较H. exiguus靠前;M1和M2的t6和t9的连接脊明显,t12为低的脊形;M2的t1和t3以及M3的t1均与t5连;M3有t3和较大而孤立的t8; m1的下前中尖(Acc)明显,与颊侧下前边尖(Abc)和舌侧下前边尖(Alc)连;m1和m2颊侧齿带明显,有较大的c2与下原尖相连,后根为低弱的脊形;m2和m3的Abc孤立;m3无c1等。根据H. asiaticus的进化程度,H. aisaticus的产出时代可能比H. exiguus的稍晚,为灞河期晚期,大致与欧洲MN11上部或MN12下部的时代相当。
关键词:
Abstract
A skull with mandible and several cervicals of a new species of Huerzelerimys, H. asiaticus, collected from the Late Miocene Liushu Formation in Linxia Basin, Gansu Province, is described in this paper. The skull is the first one ever found for the genus Huerzelerimys. Its main characters are: size small; skull broad and short with stout rostrum; interorbital roof narrow; premaxillary laterodorsal crest well developed; frontal crests weak and subparallel; incisive foramina long, with their posterior ends lined up with anterior root of M1; posterior palatal foramina located mesial to M2; caudal border of hard palate lying posterior to M3; interpterygoid foramen absent; alisphenoid canal bony; bulla large and inflated; internal carotid foramen located near the basilar tubercle; mandible having low horizontal ramus and deeply concave diastema; anterior end of masseteric ridge lined up with anterior margin of m1; mental foramen situated slightly anterior to masseteric ridge and anteroventral to m1, near to mandibular diastema; upper incisors orthodont; M1 having slightly anteriorly located t1; in M1 and M2 t6 and t9 connected by distinct crest and t12 crest-like; t1 and t3 of M2 and t1 of M3 connected to t5; M3 having t3 and a large isolated t8; small Acc of m1 connected with both Alc and Abc; m1 and m2 having distinct buccal cingula, larger c2 attached to protoconid, and low crest-like posterior heel; m2 and m3 having isolated Abc; c1 absent in m3. The evolutionary level of the described skull tends to show that the age of the upper part of the Liushu Formation yielding H. asiaticus may belong to late Bahean, corresponding to European upper MN11 or lower MN12.
Keywords:
PDF (2063KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
王伴月, 邱占祥, 李绿洲. 甘肃和政晚中新世许氏鼠(啮齿类:鼠科)头骨化石. 古脊椎动物学报[J], 2020, 58(2): 120-133 DOI:10.19615/j.cnki.1000-3118.200319
WANG Ban-Yue, QIU Zhan-Xiang, LI Lü-Zhou.
Huerzelerimys is a genus of the Murinae lived in Eurasia in Late Miocene. In the past, fossils of Huerzelerimys were mainly found in Europe, only recently they were found in Asia (Mein et al., 1993; Qiu and Li, 2008). However, most of the known fossils are isolated teeth, only a few are fragmentary jaws. In 2005, a skull with mandible and several cervicals of Huerzelerimys were collected from the Late Miocene Liushu Formation in Linxia Basin, Gansu Province by a field team of the IVPP (Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences). The skull is the first one ever found among the Huerzelerimys fossils and provides us with important new information about this genus.
The skull morphological terminology follows that of Greene (1935), Yang et al. (1985) and Wang and Qiu, 2018, and the molar terminology follows that of Qiu and Li (2016) (Fig. 1).
Fig. 1
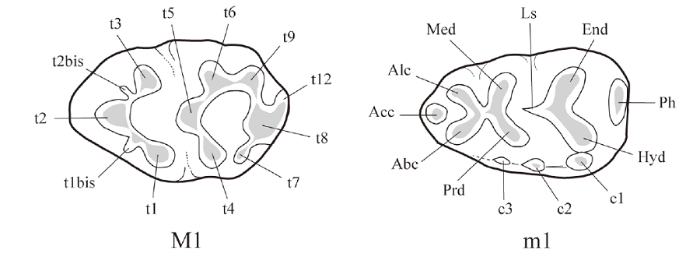 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1Nomenclature used for molars of MuridaeAbbreviations: Abc. anterobuccal cusp; Acc. anterocentral cusp; Alc. anterolingual cusp; c1?c3. accessory cusps; End. entoconid; Hyd. hypoconid; Ls. longitudinal spur; Med. metaconid; Ph. posterior hell; Prd. protoconid; t1?t12. tubercles; t1bis and t2bis. appendices to t1 and t2 (After Qiu and Li, 2016)
1 Systematic paleontology
Muridae Gray, 1821Murinae Gray, 1821
Huerzelerimys Mein et al., 1993
Type speciesParapodemys vireti Schaub, 1938.
Diagnosis (adopted from Mein et al., 1993) “Molars smaller than or similar in size to those of extant Rattus rattus, and with a poor development of the longitudinal connection between tubercles. Upper molars without t7, but with t4 and t8 connected by a weak crest. M1 and M2 with a well-developed t9, and with t6 and t9 united in more than 50% of the specimens. M3 without t9. m1 with a reduced tma (= Acc), and with a connection between the two anterior pairs of tubercle; three roots; cingular margin moderately developed. Tendency towards a strong size increase in the course of time.”
Huerzelerimys asiaticus sp. nov.
(Figs. 2?5; Tables 1?2)
Fig. 2
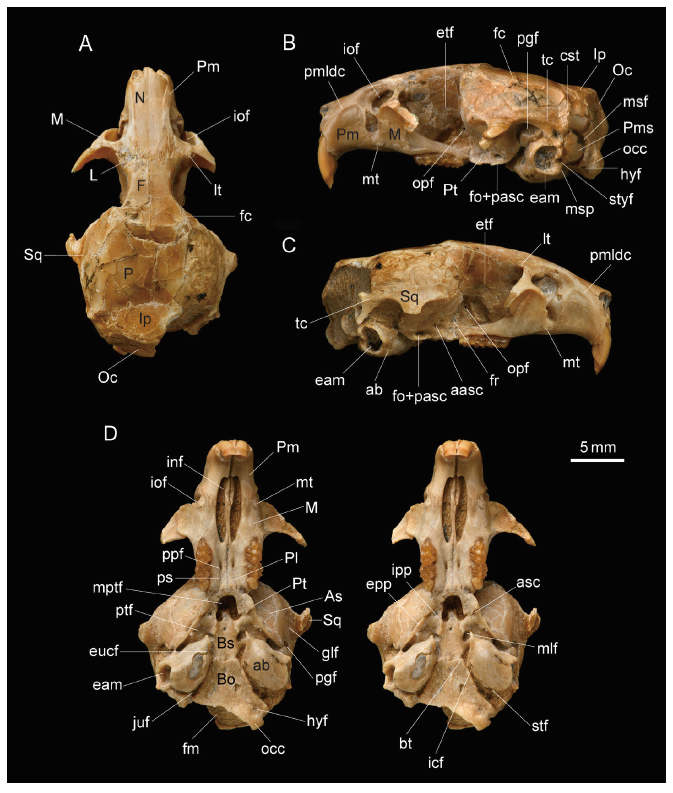 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Skull of Huerzelerimys asiaticus sp. nov. (IVPP V 16288, holotype) from Hezheng, Gansu
A. dorsal view; B. left lateral view; C. right lateral view; D. ventral view (stereopair)
Abbreviations of foramina and other structures: aasc. anterior foramen of alisphenoid canal; ab. auditory bulla; asc. alisphenoid canal; bt. basilar tubercles; cst. crista supratympanica; eam. external auditory meatus;epp. external pterygoid process; etf. ethmoidal foramen; eucf. foramen of Eustachian canal; fc. frontal crest; fm. foramen magnum; fo. foramen ovale; fr. foramen rotundum; glf. glenoid fossa; hyf. hypoglossal foramen; icf. internal carotid foramen; inf. incisive foramen; iof. infraorbital foramen; ipp. internal pterygoid process; juf. jugular foramen; lt. lacrimal tubercle; mlf. middle lacerate foramen; mptf. mesopterygoid fossa;msf. mastoid foramen; msp. mastoid process; mt. masseteric tubercle; occ. occipital condyle;opf. optic foramen; pasc. posterior foramen of the alisphenoid canal; pgf. postglenoid foramen; pmldc. premaxillary laterodorsal crest; ppf. posterior palatine foramen; ps. palatine sulcus; ptf. pterygoid fossa;stf. stapedial foramen; styf. stylomastoid foramen; tc. temporal crest Bones: As. alisphenoid; Bo. basioccipital; Bs. basisphenoid; F. frontal; Ip. interparietal; L. lacrimal; M. maxilla; N. nasal; Oc. occipital; P. parietal; Pl. palatine; Pm. premaxilla; Pms. petromastoid portion; Pt. pterygoid; Sq. squamosal
Fig. 3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3Mandible of Huerzelerimys asiaticus sp. nov. (IVPP V 16288, holotype) from Hezheng, Gansu
A1-3. left mandible; B1-3. right mandible; A1, B1. occlusal view; A2, B2. buccal view; A3, B3. lingual view+Abbreviations: ap. angular process; cdp. condyloid process; crp. coronoid process; eptf. external pterygoid fovea; i2b. bulge formed by i2; iptf. internal pterygoid fovea; mdf. mandibular foramen; mn. mandibular notch; mr. masseteric ridge; mstf. masseteric fossa; mtf. mental foramen; tf. temporal fovae
Fig. 4
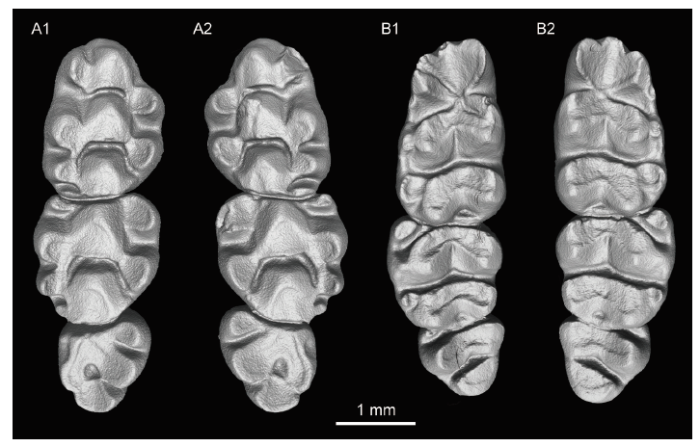 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 4Occlusal view of molars of Huerzelerimys asiaticus sp. nov. (IVPP V 16288) from Hezheng, Gansu
A1. right M1?3; A2. left M1?3; B1. left m1?3; B2. right m1?3
Fig. 5
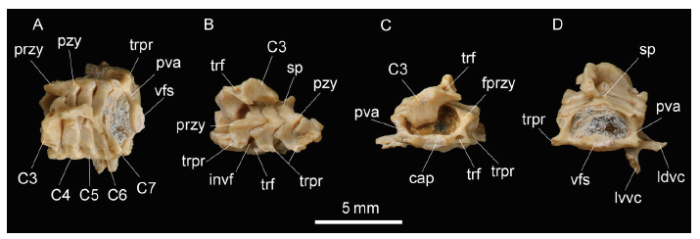 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 5Cervical vertebrae of Huerzelerimys asiaticus sp. nov. (IVPP V 16288, holotype) from Hezheng, Gansu
A. dorsal view; B. left lateral view; C. anterior view; D. posterior view+Abbreviations: C3-C7. 3rd-7th cervical vertebrae; cap. caput; fprzy. facet on prezygapophysis;invf. intervertebral foramen; ldvc. lamina dorsalis vertebrae cervicalis VI; lvvc. lamina ventralis vertebrae cervicalis VI; przy. prezygapophysis; pva. pedicle of vertebral arch; pzy. postzygapophysis;sp. spinous process; trf. transverse foramen; trpr. transverse process; vfs. verterbral fossa
Table 1
Table 1Measuremens of skull and mandible of Huerzelerimys asiaticus sp. nov. (IVPP V 16288, holotype) (mm)
| Parameter | Left | Right | Parameter | Left | Right | |
|---|---|---|---|---|---|---|
| 1. Condylonasal length (CNL) | 27+ | (15/3) (%) | 89 | |||
| 2. Condylobasal length (CBL) | 26.6+ | 16. Nasal length (NL) | 9.5+ | |||
| 3. Length of anterior part of skull (LAS) | 9 | (16/2) (%) | 36 | |||
| 4. Length of posterior part of skull (LPS) | 17.5 | 17. Anterior width of nasals (AWN) | 1.8 | 1.8 | ||
| (3/4) (%) | 51 | 18. Posterior width of nasals (PWN) | 0.7 | 0.7 | ||
| 5. Palatal length (PL) | 14.5 | 19. Interorbital width (IOW) | 4.2 | |||
| 6. Length of maxillary diastema (LMXD) | 7.3 | 20. Zygomatic width (ZW) | 15+ | |||
| 7. Length of palatal bridge (LPB) | 5 | (20/2) (%) | 56 | |||
| 8. Anterior width of palatal bridge (AWPB) | 3 | 21. Width of cranial part at mastoid processes (WCM) | 13 (6.5 × 2) | |||
| 9. Posterior width of palatal bridge (PWPB) | 3.2 | (21/2) (%) | 49 | |||
| (9/7) (%) | 64 | 22. Length of auditory bulla (LAB) | 5.4 | 5.4 | ||
| 10. Length of incisive foramen (LIF) | 6 | 6 | (22/2) (%) | 20.3 | 20.3 | |
| 11. Rostrum length (RL) | 6 | 23. Width of auditory bulla (WA) | 4 | 4 | ||
| (11/2) (%) | 22.5 | 24. Distance between two auditory bullae (DAB) | 2.7 | |||
| (10/11) (%) | 100 | 25. Nuchal height (NH) | 8e | |||
| 12. Anterior width of rostrum (AWR) | 4.5 | 26. Mandibular length (MDL) | 15.8 | 14.7+ | ||
| 13. Posterior width of rostrum (PWR) | 5.9 | 27. Height of condyloid process of mandible (HCPM) | 8.4 | |||
| (13/11) (%) | 98 | 28. Length of mandibular diastema (LMD) | 4 | 4 | ||
| 14. Rostrum height (RH) | 5 | 29. Height of mandibular diastema (HMD) | 2 | 2 | ||
| 15. Height of middle part of skull (HMS) | 8 | 30. Height of horizontal ramus of mandible (HHR) | 4.2 | 4.3 | ||
新窗口打开|下载CSV
Table 2
Table 2Measurements of teeth of Huerzelerimys asiaticus sp. nov. (IVPP V 16288, holotype) (mm)
| Parameter | Left | Right | Parameter | Left | Right | |
|---|---|---|---|---|---|---|
| M1-3 L | 4.6 | 4.6 | m1-3 L | 4.5 | 4.5 | |
| M1 L | 2.3 | 2.25 | m1 L | 2 | 1.9 | |
| W | 1.5 | 1.5 | W | 1.3 | 1.3 | |
| W/L (%) | 65 | 67 | W/L (%) | 65 | 68 | |
| M2 L | 1.7 | 1.7 | m2 L | 1.5 | 1.5 | |
| W | 1.55 | 1.5 | W | 1.46 | 1.4 | |
| W/L (%) | 91 | 88 | W/L (%) | 97 | 93 | |
| M3 L | 1.1 | 1.1 | m3 L | 1.2 | 1.3 | |
| W | 1.1 | 1.1 | W | 1.2 | 1.15 | |
| W/L (%) | 100 | 100 | W/L (%) | 100 | 88 | |
| I2 L | 1.6 | 1.6 | i2 L | 1.4 | 1.5 | |
| W | 1.1 | 1.1 | W | 0.9 | 1 |
新窗口打开|下载CSV
Holotype A complete skull with mandible and 5 cervicals (IVPP V 16288).
Locality and horizon Niuzhawan, Sanhe Village, Hezheng County (IVPP Loc. LX 200503), Linxia Hui Autonomous Prefecture, Gansu Province, China; upper part of the Liushu Formation, Late Miocene late Bahean ALMA/S (Asian Land Mammal Age/Stage), about 8 Ma.
Diagnosis Small-sized Huerzelerimys. Skull broad and short with short and broad rostrum, narrow interorbital roof, more developed premaxillary laterodorsal crest, weak and subparallel frontal crests; incisive foramen long, terminated posteriorly near the anterior root of M1; paired posterior palatal foramina situated mesial to M2; posterior border of hard palate lying posterior to M3; interpterygoid foramen absent; alisphenoid canal bony; bulla large and inflated; internal carotid foramen located near the basilar tubercle; horizontal ramus of mandible low, having a deeply concave diastema; masseteric ridge extending anteriorly below the anterior margin of m1; the mental foramen situated anteroventral to m1, anterior to the masseteric ridge, and near to mandibular diastema.
I2 orthodont; M1 with slightly anteriorly located t1; M1 and M2 with t6 and t9 connected by distinct crest and crest-like t12; t1 and t3 of M2 and t1 of M3 connected with t5; M3 with t3 and a large isolated t8; m1 with small Acc connected with both Alc and Abc; m1 and m2 having distinct buccal cingula, larger c2 attached to protoconid, and low crest-like posterior heel; m2 and m3 having isolated Abc; c1 absent in m3.
Etymology Asia, name of the continent where the fossil was collected.
2 Description
2.1 Skull
The skull is well preserved, but the zygomatic arches are broken and the posterior part of the skull is damaged. The cranial bone sutures are obscure, especially those on the skull roof (Fig. 2).The skull is small-sized among the muroid rodents, with myomorphous zygomasseteric structure. It is relatively short and broad. The condylobasal length is subequal to that of a small-sized Rattus (R. rattus). The rostral part is rather short and broad, relative to the cranial part. The length of maxillary diastema is slightly shorter than the height of the middle part of the skull.
Dorsal view The skull is roughly oval in outline. As in Rattus, the rostrum is rather short and broad, with its two lateral sides slightly convergent forwards. Rostral end of the rostrum is nearly as wide as the interorbital part. The nasal (N) is wedge-shaped. Caudally, it is lined up with the posterior end of the premaxillo-frontal sutures. The premaxillary laterodorsal crest (pmldc) is well developed, nearly parallel to the naso-premaxillary suture and separates the premaxilla (Pm) into dorsal and lateral parts. The dorsal part of the premaxilla is narrow and long, forming a strip. The premaxillo-frontal suture is strongly serrated, extending anterolaterally to meet the premaxillo-maxillary and maxillo-frontal sutures. The posterolateral end of the maxillo-frontal suture extends to the lacrimal (L). The lacrimal is situated mainly in the orbit, exposed only a little on the dorsal side, forming a distinct lacrimal tubercle (lt) at the anteromesial corner of the orbit. The coronal suture is convex posteriorly. The dorsal parts of the parietals (P) are slightly convex, much broader than those of the frontals (F). The parietals are displaced about 0.5 mm to the right side relative to the frontals, so that the parietal suture and the frontal suture are not in the same longitudinal line. The interparietal (Ip) is a large bone, elliptic in outline, wider than long.
As in other murines, the frontal crest (fc) extends backwards to the crista supratympanica (cst). The two slightly curved frontal crests are roughly parallel with each other as in Rattus, but not so well developed as in the latter. Though broken away, the zygomatic arches must be slender and their anterior roots are situated in front of the posterior ends of the nasal and premaxilla.
Lateral view The skull roof is convex dorsally. The anterior ends of the nasal and premaxilla are situated almost in the same vertical line. The diastema is much longer than the length of the upper molar row. The anterior part of the skull (anterior to M1) is shorter and lower than the posterior part of the skull (including M1), only about 1/2 as long as the latter. The lateral part of the premaxilla, bordered anterodorsally by the arched pmldc, is broad and slightly concave. The premaxillo-maxillary suture runs roughly vertically in front of the infraorbital foramen. The infraorbital foramen (iof) lies within the maxilla (M), having an oval upper part and an infraorbital fissure below. The masseteric tubercle (mt) is located below the infraorbital fissure. The insertion of masseter profundus is confined within the infraorbial foramen. The zygomatic plate is located ventrolateral to the infraorbial foramen and its surface is concave facing anteroventrally and laterally.
The posterior border of the large orbit is formed by the squamosal (Sq). The distinct temporal crest (tc) extends to the crista supratympanica (cst). The ventral surface of the large auditory bulla (ab) is lower than that of the occipital condyle (occ). The external auditory meatus (eam) is formed by a very short bony tube. The stylomastoid foramen (styf) is located behind the external auditory meatus and above the mastoid process (msp). The nuchal surface is vertical and convex posteriorly. The left petromastoid portion (Pms = os mastoideum) is preserved and nearly round in outline and has a convex surface. The mastoid foramen (msf) is situated at the mesosuperior corner of the petromastoid portion.
In the orbital area, several foramina can be observed. The optic foramen (opf) is large and located above the M3. The ethmoidal foramen (etf) is small, situated anterosuperior to the optic foramen and above the M2. The area below the etf and anteroventral to opf is broken, thus the sphenopalatine foramen cannot be fixed. The foramen rotundum (fr) is located ventroposterior to the optic foramen and superoposterior to the M3. Differing from Rattus, but similar to Apodemys, the alisphenoid canal (asc, canalis alisphenoidale) is a bony canal penetrating the pterygoid. The anterior foramen of alisphenoid canal (aasc) opens on the lateral side of the external pterygoid process (epp). The large foramen ovale (fo) penetrates the superoposterior part of the external pterygoid process, confluent with the posterior foramen of the alisphenoid canal (pasc).
Ventral view The incisive foramen (inf) is very long, about 4/5 as long as the diastema, extending posteriorly to the level of the anterior root of the M1. The zigzag premaxillo-maxillary suture intersects the incisive foramen at around the anterior 1/3 of the foramen. The zygomatic plate is broad and concave, extending anterodorsally and facing anteroventrally and laterally. The masseteric tubercle (mt) is located at the anteromesial corner of the zygomatic plate, behind the premaxillo-maxillary suture. The left and right cheek tooth rows are nearly parallel to each other. The posterior palatine foramen (ppf) is situated within the palatine (Pl), lying mesial to the M2. The distinct palatine sulcus (ps) extends to the ppf. The posterior border of the hard palate is located behind the M3. The mesopterygoid fossa (mptf) is subequal to the pterygoid fossa (ptf) in width. In the pterygoid fossa, no interpterygoid foramen (or sphenopterygoid vacuity) is observed. The middle lacerate foramen (mlf) is located at the mesioposterior corner of the pterygoid fossa. The glenoid fossa (glf) is formed by the zygomatic process of squamosal, and transversely concave. The postglenoid foramen (pgf) is long and large, extending along the petro-squamosal suture on the posterior part of the glenoid fossa. The auditory bulla is very large (about 1/5 of the condylobasal length) and strongly inflated. No bony septum is present in the broken bulla. The foramen of the Eustachian canal (eucf) opens anteromesially to the bulla. On the mesial side of the bulla, the internal carotid foramen (icf) is located on the anterior 1/3 of the bulla, nearly in the same transverse level of the basilar tubercle (bt). The jugular foramen (juf) is located between the bulla and occipital. There is a distinct stapedial foramen (stf) at the mesioposterior corner of the bulla as in Rattus (Guthrie, 1963). The hypoglossal foramen (hyf) is located anterolateral to the occipital condyle.
2.2 Mandible
The left and right hemimandibles are well-preserved, with only the tops of the coronoid processes broken away. The mandible is sciurognathous. The horizontal ramus is low, with its diastema deeply concave. As in Rattus and Huerzelerimys exiguus, the masseteric ridge (mr) extends anteriorly to below the anterior margin of m1. The mental foramen (mtf) is located anteroventral to the m1, lying near the diastema and anterior to the masseteric ridge (Fig. 3).The ascending ramus of the mandible is long. The anterior border of the coronoid process (crp) rises from the buccal side of the mandible below the anterior part of the m2, and its upper part curves slightly posteriorly. The condyloid process (cdp) is slightly lower than the coronoid process. The articular facet of the condyle is narrow-ovoid in shape, much longer than wide. The mandibular notch (mn) is very shallow. On the buccal side of the ascending ramus, the masseteric fossa (mstf) is broad. The bulge (i2b) formed by the posterior end of the lower incisor alveolus is prominent and situated below the mandibular notch on the buccal side. The angular process (ap) extends posteriorly below the condyloid process. On the lower half of the lingual side of the ascending ramus, the internal pterygoid fovea (iptf) is large, triangular in shape, with its anterior angle reaching to below the m2 and the large posterior portion deeply concave, which is sharply bordered by curved upper and lower ridges. The slightly concave temporal fovae (tf) is located in the lower part of the coronoid process and posterolateral to m3. The mandibular foramen (mdf) is situated below the mandibular notch, and the external pterygoid fovea (eptf) is slightly concave, lying anteroinferior to the condyloid process.
2.3 Teeth
The dental formula is 1·0·0·3/1·0·0·3. The molars are brachyodont and rooted. The molars are moderately worn (Fig. 4).The M1 is oval-shaped (Width/Length = 0.65, 0.67). The t1 is rounded in outline and located slightly anteriorly, with the anterior border being nearly opposite to the posterior part of the t3, and connected with the larger t2 by a crest. The t3 is smaller than t1 and t2. No tlbis is present. The t1 and t3 have no spurs extending to the t5. The t5 is connected with t4 and t6 by distinct crests. The crest between the t4 and t5 is lower than that between t5 and t6. The t4 and t8 are connected by a low crest. No t7 is present. The t9 is well developed and connected with t6 by a distinct crest, and the t12 is crest-like.
The M2 is round-angled triangular in outline, with 3 roots. The t1 is rounded. The t3 is much smaller than the t1, and both connect with the t5. The t9 is slightly larger than t3, but smaller than t6. The other features are similar to M1.
The M3 is more triangular than M2 in outline, also with 3 roots. The t1 is smaller than t4, and connected to t5. The t3 is present but very reduced. The t8 is large and isolated. The t9 is absent.
The m1 is ovoid in outline. The anterolingual cusp (Alc) and anterobuccal cusp (Abc) are subequal in size, and are connected with each other. The Alc extends slightly more anteriorly than the Abc. The small anterocentral cusp (Acc) (= tma, medial anteroconid) is heavily worn and attached to both the Abc and Alc. The Alc is connected with the metaconid. The metaconid and entoconid extend transversely, while the protoconid and hypoconid extend buccoposteriorly. The posterior pair of tubercles (hypoconid and entoconid) is separated from the second pair of tubercles (protoconid and metaconid), without longitudinal crest or longitudinal spur (Ls) connecting the two pairs. The buccal cingulum is developed, with distinct accessory cusps on it. Among them, the c1 is the largest and the c2 is larger than the c3 and attached to the protoconid. The posterior heel (Ph) (= pc, posterior cingulum) is low and crest-like, joining with the entoconid and hypoconid.
On the m2, the Abc is oval-shaped and isolated. The Alc is absent. There is no longitudinal connection between the two posterior pairs of tubercles. The buccal cingulum is broken off. The c2 is smaller than Abc, but larger than the c1, and attached to the protoconid. The posterior heel is low and crest-like, joining with the entoconid and hypoconid.
On m3, the Abc is reduced to a small and isolated cusp. The protoconid is connected with the metaconid to form a transverse crest. The c1 is absent.
The I2 is orthodont and bends strongly. Its anterior part is extending ventrally. The cross-section of the I2 is triangular in outline, longer than wide, with its labial side convex. The enamel layer covers the whole labial side and parts of the mesial and lateral sides. The labial surface is smooth without ridge on it. No gap is seen on the lingual side.
The i2 is slightly curved in longitudinal direction, with its anterior portion turning anterodorsally. The i2 originates in the ascending ramus of the mandible, forming a bulge on the buccal side. The cross-section of the i2 is narrow-ovoid, with convex labial and lateral sides, but a narrow and rounded lingual angle. The enamel covering is similar to that of the I2.
2.4 Cervical vertebrae
The 5 cervical vertebrae (C3?C7) are in quasi-articulated states and well preserved, except C3 and C7 which are more or less damaged. They are similar to each other in general morphology: the body assumes a flattened cylinder in shape; the dorsal part of the vertebral arch is bow-shaped, with the spinous process (sp) low, its pedicle short, the pre- and post-zygapophyses (przy and pzy) originated from the pedicle, and the transverse process (trpr) from the lateral side of the pedicle. In C3?C6, the transverse processes have two roots and a large transverse foramen (trf), while in C7 the transverse process has only one large root and lacks the transverse foramen. The transverse process extends posterolaterally in C3-C5, but transversely with its enlarged lateral part being separated into two laminae [lamina ventral vertebrae cervical VI (lvvc) and lamina dorsal vertebrae cervical VI (ldvc)] in C6 (Fig. 5).Measurements Total length of vertebral bodies of C4?C7: 6 mm; vertebral fossa of C7 is 0.8 mm in height and 1.8 mm in width; width of C7 at transverse process: 5 mm; width of C4 at prezygapophysis: 4.6 mm.
3 Comparison and discussion
3.1 Skull morphology compared with those of the other murines
As described above, the skull of IVPP V 16288 has myomorphous zygomasseteric structure, with the infraorbital foramen specialized into a wide upper portion and a narrow infraorbital fissure. The zygomatic plate is broadened and located ventrolateral to the infraorbital foramen and tilted upward. The frontal crest is extending backwards to the crista supratympanica and the mandible is sciurognathous. The cheek teeth are brachyodont with roots, and the crown cusps arranged in three longitudinal rows. All these characters show clearly that V 16288 belongs to the Murinae.Within the murines which have skulls the V 16288 skull is more similar to Rattus than to any other murines, such as Apodemys, Micromys, Leopoldamys, Maxomys, Niviventer etc., as evidenced in the following characters: rostrum relatively broad and short, interorbital constriction moderate, braincase rather broad, incisive foramen very long, extending to the level of the anterior part of M1, posterior part of hard palate extending slightly behind M3, and bulla large and strongly inflated, etc. (Ellerman, 1941, Huang et al., 1995, Pan et al., 2007). However, V 16288 differs from Rattus as well in having more developed premaxillnary laterodorsal crest, weaker frontal crest and bony alisphenoid canal, but lacking interpterygoid foramen in the pterygoid fossa. In dentition, V 16288 further differs from Rattus in molars being more brachyodont, M1 and M2 having crests linking t4 with t8 and t6 with t9, and having t12, and m1 having anterocentral cusp and subequal Alc and Abc.
Obviously V 16288 represents a distinct genus, more primitive than Rattus.
3.2 Tooth features compared with those of the murines known only by teeth
Some murine genera are known only by teeth. Compared with those genera, IVPP V 16288 is more similar to Progonomys and Huerzelerimys than to others in the following characters: molars being more brachyodont, upper molars with a high connection between t4 and t5, and lacking t7, M1 having more anteriorly located t1 but without t1bis, M1 and M2 having low crest between t4 and t8 and having t12, and m1 and m2 without longitudinal connections between the two posterior pairs of tubercles (protoconid-metaconid and hypoconid-entoconid), and Acc of m1 being reduced etc.Furthermore, in V 16288 M1 and M2 having well developed t9 and distinct connection between t6 and t9, M3 lacking t9, m1 having a distinct Acc, connection between the two anterior pairs of tubercles (anterobuccal-anterolingual cusps, and protoconid-metaconis), and moderately developed cingular margin. All these features of V 16288 are similar to those of Huerzelerimys rather than to Progonomys.
In addition, according to Mein et al. (1993:59), the incisive foramen extends only to the level mesial to the anterior root of M1 in H. minor, but it can extend farther backward beyond the anterior root of M1 in Progonomys. In this feature V 16288 resembles H. minor rather than Progonomys. Meanwhile, the mandible of V 16288 is also similar to that of H. exiguus. Therefore, V 16288 should be referred to the genus Huerzelerimys.
3.3 Comparison of IVPP V 16288 with known species of Huerzelerimys
The genus Huerzelerimys is known to include 5 species: H. vireti (Schaub, 1938), H. turoliensis (Michaux, 1969), H. oreopitheci (Engesser, 1989), H. minor (Mein et al., 1993) and H. exiguus (Qiu and Li, 2008).The cheek teeth of V 16288 are much smaller than those of H. turoliensis and H. oreopitheci, but larger than those of H. exiguus, and subequal to those of H. vireti and H. minor (See: Mein et al., 1993:fig. 4; Qiu and Li, 2008:table 4).
Morpghologically, V 16288 is different from H. minor and H vireti in M1 and M2 having distinct crest connecting t6 with t9 and having a crested t12, m1 having nearly centrally located Acc uniting with both Alc and Abc, m1 and m2 having larger c2 attached to protoconid and having lower and crested Ph, and m3 lacking c1.
V 16288 differs from H. minor in M3 having isolated t8, and Abc of m2 and m3 being isolated. V 16288 further differs from H. vireti in t1 and t3 of M2 and t1 of M3 being connected to t5. Therefore, V 16288 cannot be attributed either to H. minor, nor to H. vireti.
V 16288 differs from H. exiguus in M1 having slightly anteriorly located t1, and t6 and t9 being distinctly connected; m1 having Acc attached to both the Alc and Abc; m1 and m2 having more developed accessory cusps, and lower and weaker Ph.
V 16288 differs from H. turoliensis in M1 and M2 having more developed t12, m1 having distinct Acc, m1 and m2 having more distinct buccal accessory cusps and having lower crested-like Ph.
V 16288 differs from H. oreopitheci in molars being more brachyodont, M1 and M2 having low t12 and smaller t9, M3 having t3, and m1 having more developed Acc and m1 and m2 having larger c2 attached to protoconid, and lower crested-like Ph.
To sum up, V 16288 represents a new species of Huerzelerimys, here named as H. asiaticus.
3.4 Evolutionary consideration regarding Huerzelerimys asiaticus
Judging from the previously known species of Huerzelerimys, the overall evolutionary tendencies of Huerzelerimys can be summed up as follows: increase in size (a), increase in crown height (b), forward shifting of t1 in M1 (c), gradual convergence and union of t6 and t9 (d), reduction of t12 in M1 (e), reduction of Acc in m1 (f), and development of accessory cusps in m1 (g) (Mein et al., 1993; Martín Suárez and Freudenthal, 1993; Freudenthal and Martín Suárez, 1999; Qiu and Li, 2008).As discussed above, H. asiaticus differs from H. exiguus in having such advanced features as a, c, d and g. This shows that H. asiaticus is more advanced than H. exiguus.
Although it is subequal to H. minor and H vireti in size, H. asiaticus has some more advanced features (d and g) than the two latters. It seems that H. asiaticus may represent a slightly more advanced species than H. minor and H vireti.
Although it has more developed accessory cusps in m1, an apomorphic character in Huerzelerimys (g), H. asiaticus is less advanced than H. turoliensis in features a, e and f, and is different from H. oreopitheci in lacking such advanced features as a, b, e and f. Obviously H. asiaticus may represent a more primitive species than the two latters.
To sum up, H. asiaticus is more advanced than H. exiguus, H. minor and H. vireti, but more primitive than H. turoliensis and H. oreopitheci.
Since the age of H. exiguus is early Late Miocene, roughly corresponding to upper MN10 or lower MN11 in European time scale (Qiu and Li, 2008), H. minor is known from upper MN10, H. vireti from MN11, and H. turoliensis from upper MN11 to MN12 (Mein et al., 1993), and H. oreopitheci is known from middle Turolian (middle MN12) (Engesser, 1989), the age of the upper part of the Liushu Formation yielding H. asiaticus may better be estimated as late Bahean, corresponing to upper MN11 or lower MN12.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 6]
[本文引用: 1]
[本文引用: 2]
[本文引用: 4]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}