 ,1, 金昌柱2, Jay KELLEY3,4, Nina G. JABLONSKI5, 吉学平6, Denise F. SU7, 邓涛2,8,9, 李强2,8
,1, 金昌柱2, Jay KELLEY3,4, Nina G. JABLONSKI5, 吉学平6, Denise F. SU7, 邓涛2,8,9, 李强2,8Late Miocene fossil calibration from Yunnan Province for the striped rabbit Nesolagus
Lawrence J. FLYNN,1, JIN Chang-Zhu2, Jay KELLEY3,4, Nina G. JABLONSKI5, JI Xue-Ping6, Denise F. SU7, DENG Tao2,8,9, LI Qiang2,8通讯作者: ljflynn@fas.harvard.edu
收稿日期:2018-12-22网络出版日期:2019-07-20
| 基金资助: |
Corresponding authors: ljflynn@fas.harvard.edu
Received:2018-12-22Online:2019-07-20

摘要
报道了发现于云南昭通水塘坝的晚中新世兔类,它与时代较早的云南禄丰地点的长褶“翼兔”(“Alilepus” longisinuosus)属于同一个种,两者栖息的环境均为潮湿的沼泽地区。前臼齿的形态表明该种是现今生活在湿润地区条纹兔(即苏门答腊兔Nesolagus)的早期代表,因此将该种更名为长褶苏门答腊兔Nesolagus longisinuosus (
关键词:
Abstract
The rabbit from the Late Miocene Shuitangba site in Yunnan Province represents the same species as that from the older site of Lufeng, Yunnan. Both settings record a wet, swampy habitat. Premolar morphology shows that the species is an early representative of the extant striped rabbit genus, which today lives in moist habitat, and should be designated Nesolagus longisinuosus (
Keywords:
PDF (2583KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
Lawrence J. FLYNN, 金昌柱, Jay KELLEY, Nina G. JABLONSKI, 吉学平, Denise F. SU, 邓涛, 李强. 云南晚中新世苏门答腊兔(Nesolagus)分离时间的标定. 古脊椎动物学报[J], 2019, 57(3): 214-224 DOI:10.19615/j.cnki.1000-3118.190326
Lawrence J. FLYNN, JIN Chang-Zhu, Jay KELLEY, Nina G. JABLONSKI, JI Xue-Ping, Denise F. SU, DENG Tao, LI Qiang.
1 Introduction
The developing fossil record of South China, especially Yunnan Province, holds the solutions to several evolutionary and biogeographic puzzles. In contrast to the Late Miocene vertebrate record of North China (Baodean Mammal Stage-Age), which is well-represented by many localities, the South China theater of evolution has yielded few assemblages documenting the different fauna and paleoecology of the region. Late Miocene fossils of South China include distinctive taxa at the species and higher taxonomic levels, some of which may be precursors of Recent Southeast Asian mammalian lineages. The new species Tapirus yunnanensis Ji et al., 2015 and Muntiacus zhaotongensis Dong et al., 2014 are examples. Small mammal fossils, such as tree shrews are no exception. The site of Lufeng with Prodendrogale yunnanica Qiu, 1986, records a northern record of the tree shrew family. Here, the Late Miocene Lufeng and Shuitangba sites elucidate the origin of an enigmatic lagomorph.The living striped rabbits of Sumatra and mainland Southeast Asia (Nesolagus netscheri and N. timminsi) had no known precursor until Qiu and Han (1986) drew the tentative conclusion that the origin of Nesolagus involved an endemic species from Lufeng, Yunnan, then placed in the fossil genus Alilepus. Jin et al. (2010) agreed when they found Pleistocene Nesolagus in neighboring Guangxi Province. The rich Late Miocene mammal locality at Lufeng (7 to 6 Ma, Qi et al., 2006), spans about five meters of lignite with diverse mammalian fossils in a swampy paleoecological environment. Among the small mammal component is a leporine rabbit named on evidence previously available as a species of Alilepus, A. longisinuosus. Morphological variants among the dozens of specimens suggested to Qiu and Han (1986) that A. longisinuosus might represent a transition toward extant Nesolagus.
In recent years, new rabbit fossils have been recovered from Shuitangba, another Late Miocene site in Yunnan (Fig. 1). Near Zhaotong, 300 km northeast of Lufeng, the site is close to the northern limit of the South China paleobiogeographic region and has a fauna very much like that of Lufeng (Jablonski et al., 2014). At ~6.2 Ma (Ji et al., 2013) Shuitangba is younger, includes lignite beds, and samples a wet, water-side paleoenvironment. Its rabbit cannot be distinguished from Lufeng Alilepus longisinuosus but, unlike the sample from Lufeng, the new material uniformly presents Nesolagus morphology that is advanced with respect to typical Alilepus. However, new leporid fossils from Shuitangba are still few. Mandibular specimen ZT 2015-0285 with complete dentition was found appressed to a block of dark clay during the intensive quarrying by IVPP in March 2015. Later in that field season edentulous jaw ZT 2015-0207 was retrieved. Collecting under the direction of Ji Xueping the year before produced palate fragment ZT 2014-0391. Previous field seasons involved trial wet screening and yielded a jaw fragment and an isolated premolar.
Fig. 1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1Location of fossil site Shuitangba in Yunnan Province, South China
Here we document the new leporid fossils from Shuitangba, showing their affinity to the Lufeng rabbit and to extant Nesolagus. As an early representative of Nesolagus, N. longisinuosus confirms a Late Miocene time frame for the differentiation of the modern genus lineage.
2 Systematic paleontology
Order Lagomorpha Brandt, 1885Family Leporidae Gray, 1821
Genus Nesolagus Forsyth Major, 1899
Nesolagus longisinuosus (Qiu & Han, 1986)
Alilepus longisinuosus Qiu & Han, 1986
Alilepus sp. Ji et al., 2013
Alilepus longisinuosus Jablonski et al., 2014
Alilepus longisinuosus Ji et al., 2015
Referred material ZT 2007-2-0063, isolated right p3; ZT 2010-3-0345, left dentary fragment with p3-4, m1; ZT 2014-0391, right maxilla and palatine fragment with all cheek tooth alveoli, P4 and M1-2; ZT 2015-0207, edentulous right mandible with all alveoli; ZT 2015-0285, left mandible with incisor, p3-4, m1-3.
Locality Shuitangba, local spring and lignite quarry in Zhaotong Basin, northeastern Yunnan Province. All specimens from the same “peaty clay” unit that yielded the Lufengpithecus skull reported by Ji et al. (2013), provenance is about 1 m above the skull at the 8-meter level in fig. 2e of Ji et al. (2013).
Fig. 2
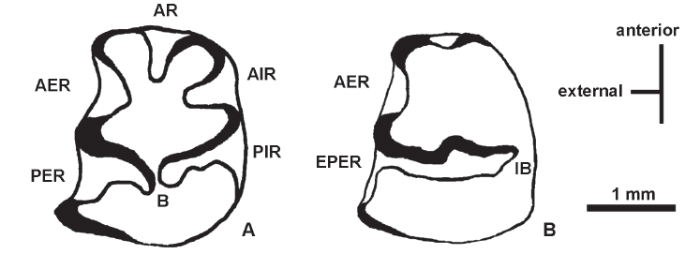 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Simplified leporid p3 dental terminology
A. idealized p3 showing reentrant positions and central trigonid-talonid bridge (B); B. Nesolagus-like p3 showing extended posteroexternal reentrant (EPER) and the bridge in internal position (IB)
AR. anterior reentrant; AER. anteroexternal reentrant; AIR. anterointernal reentrant;PER. posteroexternal reentrant; PIR. posterointernal reentrant
Age Faunal data place the assemblage in the Late Miocene; paleomagnetic correlation to a long Zhaotong Basin section dates the site to ~6.2 Ma (Ji et al., 2013).
Emended diagnosis Close in size to Nesolagus sinensis Jin et al., 2010, premolars toward the lower end of the size range for N. sinensis, but molars slightly longer relative to N. sinensis; short diastema, slightly longer than that of N. sinensis; prominent anterior and posterior external reentrants of p3 contain cement, but the reentrant in the anterior internal position is undeveloped and lacks cement (reentrant terminology of Jin et al., 2010 modified slightly in Fig. 2); trigonid unexpanded anteroposteriorly, unlike Alilepus; posterointernal reentrant of p3 variable, progressively reduced by expansion of the opposed posteroexternal reentrant, with the median bridge displaced to the lingual wall of p3 and the posterointernal reentrant eliminated; other features after Qiu and Han (1986): anterolingual fold shallow and anterobuccal fold distinct on P2, incisive foramen ends at anterior portion of P2.
Measurements See Table 1; cheek tooth lengths and widths include upper dentition for ZT 2014-0391. Lengths of tooth rows at the alveolus level are greater than at the occlusal level of the crowns where teeth converge slightly; diastema length for damaged ZT 2015-0207 is approximate.
Table 1
Table 1Tooth dimensions of Nesolagus longisinuosus from Shuitangba (mm)
| Specimen | p3 L | p3 W | p4 L | p4 W | m1 L | m1 W | m2 L | m2 W | m3 L | m3 W | L: alveoli | L: occlusal | Diastema |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ZT 2015-0285 | 2.95 | 2.62 | 2.44 | 3.29 | 2.78 | 3.18 | 2.81 | 3.01 | 1.9 | 1.65 | 15 | 12.3 | 14 |
| ZT 2015-0207 | 17.2 | 15 | |||||||||||
| ZT 2007-0063 | 2.92 | 2.75 | |||||||||||
| ZT 2010-0345 | 2.99 | 2.95 | 2.48 | 3.05 | 2.7 | 2.96 | |||||||
| P4 L | P4 W | M1 L | M1 W | M2 L | M2 W | ||||||||
| ZT 2014-0391 | 2.04 | 3.65 | 2.17 | 3.68 | 1.86 | 3.55 | 15.1 |
新窗口打开|下载CSV
Description ZT 2015-0285 (Fig. 3) is a complete dentary of a young adult with dentition, except for the posterior extremity of the gonial angle and parts of the ascending ramus. Typical for leporids, p3 is the largest and most complex cheek tooth. The trigonid is short, but broadly rounded anteriorly, and the main buccal reentrant encroaches on the lingual wall of p3, as in young individuals of Nesolagus. Widths of cheek teeth progressively decrease posteriorly. The smallest tooth, m3, has a reduced posterior lobe and its base curves posteriorly so that the alveolus is elongated. The flat face of the incisor, 3.15 mm across, is angled laterally; perpendicular to this surface, the incisor axis in cross section is 2.45 mm. Enamel extends around most of the medial side of the incisor (Fig.3). The mental foramen is above the midlevel of the diastema, 3.7 mm anterior to p3. The diastema is not exceptionally long; the ratio of diastema length to diastema plus alveolar row length is 0.48.
Fig. 3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3Left mandible ZT 2015-0285 in early wear, with incisor and all cheek teeth, p3-4, m1-3 Lingual view above, buccal view below, occlusal view in the middle shows anterior to the left
The edentulous mandible ZT 2015-0207 (Fig. 4) is somewhat larger than ZT 2015-0285. The coronoid rises steeply, but is not vertical, and the mental foramen, 3.5 mm anterior to p3, is high on the diastema; mandibular depth is 12 mm from the p4 alveolar margin to the inferior margin of the dentary.
Fig. 4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 4Buccal (right) and lingual (left) views of edentulous right mandible ZT 2015-0207
ZT 2010-0345 (Fig. 5) is a partial mandible of an apparently older individual. The mental foramen is about 2.5 mm anterior to p3. The thin enamel of the anterior walls of the talonids of p4 and m1 undulates gently. The deep buccal reentrants are filled with cementum; the anteroexternal reentrant of p3 also bears cementum, but there is none on the lingual side of the teeth. The p3 resembles the isolated p3, ZT 2007-0063 (Fig. 6), which preserves 11 mm of crown height. The main buccal reentrant (PER) of both is very deep, extending nearly to the lingual wall of the tooth. A very thin connection of dentine is exposed there. There is no median bridge of dentine, but just buccal to the midline the thick enamel of the posterior wall of the trigonid flexes toward the talonid. Both teeth have a more triangular and anteroposteriorly longer trigonid than the p3 of ZT 2015-0285, a feature more typical of Alilepus than Nesolagus.
Fig. 5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 5Left dentary fragment ZT 2010-0345 with p3-4 and m1 Buccal view above, lingual view below, occlusal view in the middle. Cementum in the buccal folds of p3 is well developed
Fig. 6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 6Occlusal view of isolated right p3, ZT 2007-2-0063
The maxilla and its dentition are represented by only one specimen, ZT 2014-0391 (Fig. 7), a right maxilla with P4 and M1-2 and all cheek tooth alveoli, and preserving a portion of the palatine bone. The latter extends anteriorly to opposite P4, where the palatine foramen is located. The incisive foramen extends posteriorly to a point opposite the middle of P2. Judging by the alveoli of the missing teeth, M3 is smallest, while P2 is oval and larger. P3 is asymmetrical with a buccally expanded metacone. The preserved cheek teeth are uniform in morphology, showing a deep internal reentrant with thin, gently wrinkled enamel, and heavy cementum within the reentrant exposed lingually. The root of the zygomatic process juts laterally opposite P4 and P3, and superior to the cheek teeth, about 3 mm above the alveolar margin.
Fig. 7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 7Right maxilla and palatine fragment ZT 2014-0391 with all cheek tooth alveoli, P4 and M1-2 Lateral view left, ventral view right
3 Discussion
Alilepus is a Miocene age leporine that evolved in North America and spread to Eurasia around 8 Ma (Flynn et al., 2014). Its dentition is distinctive, particularly p3 with a shallow anteroexternal reentrant, and deep posteroexternal and posterointernal reentrants. The two posterior reentrants flank a narrow central bridge between the trigonid and talonid. In describing the large sample of rabbits from Lufeng, Qiu and Han (1986) noted that many specimens of the diagnostic lower premolar (p3) resembled the genotypic species Alilepus annectens (Schlosser, 1924), although the p3 midline bridge between the trigonid and talonid was quite narrow when present.The opposed external and internal posterior reentrants of Alilepus are fully transformed in extant Nesolagus. The posteroexternal reentrant extends lingually to the internal wall of the tooth; the central bridge is eliminated, and there is no posterointernal reentrant. Either the trigonid and talonid are completely isolated, or a thin connection persists at the internal lingual wall. The single specimen of Pleistocene Nesolagus sinensis would show a thin lingual dentine connection in late ontogeny (toward the base of the p3 crown), although at an earlier wear stage (the surface of p3) the trigonid and talonid were totally isolated (Jin et al., 2010).
The Lufeng fossil sample shows a range of p3 morphologies from Alilepus-like premolars to a minority in which the trigonid and talonid are isolated (Fig. 8). Some Lufeng p3 have a very deep posteroexternal reentrant without a central bridge but retain a thin connection at the lingual extremity of the crown. The absence of the bridge with a consequent long external reentrant as a variant in the Lufeng sample led to the name Alilepus longisinuosus Qiu & Han, 1986 and the proposal that the species might lie near the ancestry of Nesolagus.
Fig. 8
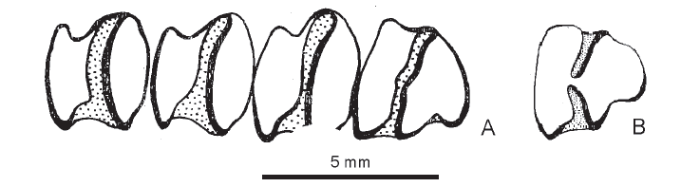 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 8Examples of Lufeng premolar variation from Qiu and Han (1986)
A. holotype IVPP V 8131 dentition of Nesolagus longisinuosus, right p3-4 and m1-2, anterior to right;B. reversed image of p3 V 8132.41 at same
scale. A shows a deep PER (no PIR) on p3 as in Nesolagus;B shows the Alilepus morphotype with retained bridge joining trigonid and talonid
The geographically (and temporally) closest rabbit locality to Shuitangba is Lufeng. Dental sizes for the Shuitangba specimens fall near the means or toward the lower end of the ranges of those from Lufeng (Qiu and Han, 1986). The morphology of the three p3 from Shuitangba is identical to that of the advanced Lufeng morphotype in lacking a dentine bridge between trigonid and talonid. The p3 crown is dominated by a very deep external reentrant reaching the lingual wall of the tooth; the posterointernal reentrant is absent and the complete lingual wall borders a thin fold of dentine connecting trigonid to talonid. As in similar Lufeng specimens, an enamel flexure buccal to the midline of the tooth marks the former position of the bridge.
Remains of the Shuitangba rabbit are compatible with those of the Lufeng rabbit but differ in that all three p3 from Shuitangba show Nesolagus morphology. The geologically younger material from Shuitangba currently available shows that the premolars, variably derived at Lufeng, were more consistently like Nesolagus at Shuitangba. The absence of the trigonid bridge, with consequent long posteroexternal reentrant and lost posterointernal reentrant are derived Nesolagus traits necessitating transfer of A. longisinuosus to that genus. N. longisinuosus illustrates progressive intraspecific transformation toward typical Nesolagus morphology and origin of the genus in the Late Miocene wet habitat of Yunnan.
Comparison In recognizing fossil Nesolagus for the first time, Jin et al. (2010) based N. sinensis on a Pleistocene mandible from Guangxi Province in which a median bridge on p3 is entirely lacking. The talonid and trigonid are joined lingually by enamel and dentine low on the crown toward the root. About the size of N. longisinuosus from Yunnan, N. sinensis is also close to extant Nesolagus in size. Jin et al. (2010) endorsed explicitly the ancestral relationship of “Alilepus” longisinuosus to N. sinensis and later species of Nesolagus.
Nesolagus sinensis shows complete separation of the p3 trigonid from the talonid at an early stage of wear (lingual connection in late wear), unlike most N. longisinuosus from Lufeng, for which the p3 retains a central or lingual connection. Other differences are minor. The lower teeth of N. sinensis are somewhat larger than the Shuitangba Nesolagus specimens (compare Table 1 with Jin et al., 2010); p3 size for Shuitangba N. longisinuosus is near that of N. sinensis, but the molars are relatively longer and narrower in the former. Shuitangba Nesolagus has a diastema length ratio of 0.48, close to that of the Lufeng sample (with ratio of 0.50 based on measurements of Qiu and Han, 1986). Both are close to the 0.46 ratio of Nesolagus sinensis cited by Jin et al. (2010).
Paleoecology Both the Lufeng and Shuitangba sites contain very dark clays, shell beds, fossil wood, and aquatic vertebrate fossils. Sediments accumulated in a reducing swamp environment, and faunal elements from both sites, for example a beaver and bamboo rats, indicate wet conditions. The Shuitangba fauna included a tapir (Ji et al., 2015) and diverse waterfowl. At Shuitangba a large otter (Wang et al., 2018) also attests to water. The Shuitangba ape (Ji et al., 2013) is very similar to Lufengpithecus lufengensis, and these suggest dependable sources of food (likely fruit) at both sites. As for these elements of the fauna, the paleoecological setting for Yunnan Nesolagus is waterside intermountain basins with wet conditions likely for much of the year. Fossil Nesolagus habitat contrasts with that of the cursorial Alilepus, its Holarctic predecessor.
The nocturnal Annamite striped rabbit of Laos and Viet Nam, Nesolagus timminsi, lives in moist habitat without an appreciable dry season (Schai-Braun and Hackl?nder, 2016), and may present a reasonable analogue for Nesolagus longisinuosus. Today other rabbits favor a waterside niche, an example being the North American swamp rabbit, Sylvilagus aquaticus, which swims readily.
4 Conclusion
The diagnostic Nesolagus premolar morphology, occasionally present in the Lufeng specimens that were previously assigned to Alilepus longisinuosus, appears to be more prevalent in the younger Shuitangba sample of that species. Its occurrence and then predominance in the species requires systematic transfer of the species to Nesolagus, as N. longisinuosus. The wet habitat occupied by N. longisinuosus contrasts with the presumed open habitat of Alilepus annectens of North China. Prior to the Late Miocene (~8 Ma, Flynn et al., 2014) modern leporines were absent from the Old World. Late Miocene leporines traversed high latitude steppe and woodland terrain in dispersing from North America to northern Asia. Rabbits later penetrated the mesic habitat of South China in the form of early representatives of Nesolagus. Neither Nesolagus nor Alilepus are known from the older Yuanmou sites of Yunnan Province, dating to ~8 Ma (Qi et al., 2006).The Late Miocene swamp rabbit of Yunnan successfully occupied South China wetlands by about 7 Ma (the greater limit for Lufeng age estimate being about 6.9 Ma, Qi et al., 2006). Surridge et al. (1999) reported their initial mitochondrial analysis of the two living species of Nesolagus and estimated that the considerable difference between them suggested genetic origin about eight million years ago. The current paleontological record of 7 Ma is a minimum age for calibration of the origin of the genus and is reasonably concordant with the Late Miocene 8 Ma molecular split estimate. The fossil evidence calls for distinction of Nesolagus by ~7 Ma, and Nesolagus presumably spread through Southeast Asia to Sumatra about that time. Characteristic in the wet habitat of the Late Miocene of South China, the Yunnan Nesolagus lineage survives to the present as the modern striped rabbits of Southeast Asia.
Acknowledgements
We thank Qiu Zhuding for advice and consultation throughout this study. IVPP organized the 2015 fieldwork and provided preparation and photography of specimens. We appreciate the support of the officials of the Zhaotong Museum, and our discussions with Michèle Morgan and John Barry helped frame some of the ideas herein. This research was supported by grants from the National Natural Science Foundation of China (41430102) and the Chinese Academy of Sciences (XDB26000000), and by grant 1227927 from the US National Science Foundation. We thank George Chaplin for Figure 1.参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
DOIURL [本文引用: 2]
DOIURL [本文引用: 2]
DOIURL [本文引用: 6]
[本文引用: 3]
DOIURL [本文引用: 8]
DOIURL [本文引用: 3]
[本文引用: 1]
[本文引用: 12]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}