 ,1,2,*
,1,2,*Cranial osteology of Beipiaosaurus inexpectus(Theropoda: Therizinosauria)
LIAO Chun-Chi1,2,3, XU Xing,1,2,*收稿日期:2018-06-5网络出版日期:2019-04-20
| 基金资助: |
Corresponding authors: *xu.xing@ivpp.ac.cn
Received:2018-06-5Online:2019-04-20

摘要
意外北票龙(Beipiaosaurus inexpectus)是研究镰刀龙类演化的关键物种,自从徐星等1999年简短报道以来就未被详细描述。对意外北票龙正型标本头部骨骼进行了详细研究。意外北票龙的头部骨骼结构有些独特之处,如额骨的眶后骨突大且明显外侧延伸、顶骨有一细长的前突、顶骨的鳞骨突腹侧支加长以及外下颌孔背腹侧深且位置非常靠后。一系列头部骨骼的祖征(如较大的齿骨和齿骨联合缝处下弯角度较小)揭示意外北票龙是镰刀龙类的一个早期演化分支,符合先行研究所提出的结论。新的数据不只对了解意外北票龙的头部骨骼结构非常重要,更对镰刀龙类演化的研究有显著意义。
关键词:
Abstract
Beipiaosaurus inexpectus, a key taxon for understanding the early evolution of therizinosaurians, has not been fully described since it was briefly reported on by Xu, Tang and Wang in 1999. Here we present a detailed description of the cranial anatomy of the holotype of this theropod dinosaur. B. inexpectus is unique in some of its cranial features such as the postorbital process of the frontal is large and its abrupt transition from the orbital rim, a long and sharp anterior process of the parietal, the elongate ventral ramus of the squamosal process of parietal, and external mandibular fenestra deep dorsoventrally and extremely posteriorly located. A number of plesiomorphic cranial features (such as relatively large dentary and less downturned degree of dentary symphysis) suggest that B. inexpectus is an early-branching Therizinosaurian, as proposed by previous studies. New information derived from our study is not only important for our understanding of the cranial anatomy of B. inexpectus but also significant to the study of the evolution of Therizinosauria.
Keywords:
PDF (4968KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
廖俊棋, 徐星. 意外北票龙(兽脚类:镰刀龙类)头部骨骼学研究. 古脊椎动物学报[J], 2019, 57(2): 117-132 DOI:10.19615/j.cnki.1000-3118.190115
LIAO Chun-Chi, XU Xing.
Therizinosauria is a maniraptoran theropod clade, which is defined here as the most inclusive clade including Therizinosaurus, but not Ornithomimus velox or Oviraptor philoceratops (Zanno, 2010a). The late-branching members of this clade, i.e., the Therizinosauridae (defined as the least inclusive clade including Therizinosaurus and Erlianosaurus), have a distinctive combination of skeletal features, such as rostrally edentulous snout, dentary with a lateral shelf, small tightly packed lanceolate teeth, elongate neck, extremely broad and opisthopubic pelvis, shortened tibia, and tetradactyl pes (Clark et al., 1994, 2004; Zanno, 2006, 2010b). To date, this clade is known from the Cretaceous period of Asia and North America (Barsbold, 1976; Perle, 1979; Russell and Dong, 1993; Xu et al., 1999; Kirkland et al., 2005), although Eshanosaurus deguchiianus from the Lufeng Formation, Yunnan, China might represent an Early Jurassic taxon (Xu et al., 2001; Barrett, 2010).
Beipiaosaurus inexpectus from the Lower Cretaceous Yixian Formation in Liaoning, China is a key taxon for understanding therizinosaurian evolution because its discovery provides direct evidence for the presence of filamentous feathers in this clade and demonstrates the independent evolution of sauropodomorphan-like features such as the tetradactyl pes in late-branching members of the clade (Xu et al., 1999). Despite the importance of Beipiaosaurus, only three brief papers have been published on the skeletal and integumentary features of this taxon (Xu et al., 1999, 2003, 2009). A detailed osteological description of the holotype specimen (IVPP V 11559) is needed in order to fully understand this important taxon. Here we present such a description of the cranial osteology, which will certainly shed new light on the cranial evolution within Therizinosauria.
Institutional abbreviations IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China; UMNH VP, Utah Museum of Natural History, Salt Lake City, Utah.
Anatomical abbreviations a.f, frontal articulation; amp, anteromedial process; an, angular; ap.fp, accessory process of frontal process; a.sup, supraoccipital articulation; dt, denticles; emf, external mandibular fenestra; en, external naris; ff, floccular fossa; fp, frontal process; f.pp, postorbital process of frontal; jp, jugal process; lf, lateral foramina; l.p, left parietal; ls, lateral shelf; mc, mandibular condyle; mg, Mecklian groove; ob, orbital; pld, posterolateral depression; po.sqp, squamosal process of postorbital; p.pp, postorbital process of parietal; pqf, paraquadrate foramen; p.sqp, squamosal process of parietal; ptf, pterygoid flange; r.f, right frontal; r.ls, right laterosphenoid; rp, replacement pits; sa, surangular; stf, supratemporal fenestra; tc, tooth crown; tr, tooth root; vo, vomer; V2, foramen for maxillary branch of the trigeminal nerve; VII, foramen for the facial nerve; VIII, foramen for the vestibulocochlear nerve.
1 Systematic paleontology
Dinosauria Owen, 1842Theropoda Marsh, 1881
Coelurosauria sensu Gauthier, 1986
Therizinosauria Russell & Dong, 1993
Therizinosauroidea Maleev, 1954
Beipiaosaurus inexpectusXu et al., 1999
(Figs. 1-3)
Holotype1)(1) After Beipiaosaurus inexpectus was named in 1999, additional material of the holotype (IVPP V 11559) was collected from the type locality (Xu et al., 2003). The holotype thus comprises more skeletal elements than listed in the original naming paper.) IVPP V 11559, a partial, semi-articulated skeleton including some cranial elements and most of the postcranial elements.
Type locality and horizon Sihetun locality near Beipiao, Liaoning, China. Lower Cretaceous (Aptian) Yixian Formation (Swisher et al., 2001).
Revised diagnosis (from cranium only) A small therizinosaurian distinguishable from other therizinosaurians in having the following cranial autapomorphies: postorbital process of frontal large and abruptly transits from orbital rim, parietal with a long and sharp anterior process, ventral ramus of parietal squamosal process extremely long, and external mandibular fenestra deep dorsoventrally and extremely posteriorly located.
2 Description and comparisons
The orbit is large and probably more anteriorly than laterally positioned as indicated by the triangular frontal with a strongly laterally extended postorbital process (Fig. 1A). The supratemporal fenestra is relatively small as indicated by transversely wide parietals, and it is sub-trapezoidal in outline, with the medial border being much shorter than the lateral border (Fig. 1B). The external mandibular fenestra is large, dorsoventrally deep, and located close to the posterior end of the mandible (Fig. 1C, D). This is mostly probably apomorphic for Beipiaosaurus inexpectus.2.1 Skull
Nasal One thin bone is tentatively identified as a nasal, but it is unknown which side of the snout it comes from (Fig. 2A). It is a flat bone with a slender anterior portion, which represents the premaxillary process, and a wide posterior portion, which represents the main body. The posteriormost portion of the bone is missing. The bone lacks a transverse arching that is typical of the theropod nasal, but this might be due to a postmortem distortion. As in Erlikosaurus (Clark et al., 1994; Lautenschlager et al., 2014) and Jianchangosaurus (Pu et al., 2013), the nasal is an elongate bone to join the formation of a long rostrum, unlike the robust and subtriangular outline of Falcarius (Zanno, 2010b). The premaxillary process is extremely slender, about one-third of the main body in width, and this ratio is smaller than in Erlikosaurus where it nearly reaches 50% (Clark et al., 1994). It tapers anteriorly with a straight ventral margin to form the posterodorsal border of the external naris as in Erlikosaurus (Clark et al., 1994) and Jianchangosaurus (Pu et al., 2013). The elongated premaxillary process indicates the presence of a long external naris as in other therizinosaurians such as Jianchangosaurus and Erlikosaurus. Also similar to other therizinosaurians, no distinct maxillary process (subnarial process) is present. The main body is flat and few informative features are available.Frontal The posterior half of the right frontal is preserved in dorsal view (Fig. 1A). The frontal is inferred to be sub-triangular in outline as in other therizinosaurians, including Falcarius (Zanno, 2010b), Jianchangosaurus (Pu et al., 2013), and Erlikosaurus (Clark et al., 1994), but its posterior portion is transversely broader relative to the anterior portion (three times as wide as the latter), which is in contrast to that in most other therizinosaurians (typically twice as wide). The frontal is flattened dorsally as in Falcarius (Zanno, 2010b) and Jianchangosaurus (Pu et al., 2013), different from the domed frontal seen in Erlikosaurus and most other coelurosaurians (Zanno, 2010b; Lautenschlager et al., 2014). The lateral edge of the frontal appears to be smooth as in early-branching therizinosaurians, including Falcarius (Zanno, 2010b) and Jianchangosaurus (Pu et al., 2013), but it is rugose in Erlikosaurus (Lautenschlager et al., 2014). The postorbital process is prominent and has an abrupt transition from the orbital rim (Fig. 1A), while most therizinosaurians seem to have a small postorbital process which smoothly transits from the orbital rim in dorsal view. The posterior margin of the frontal transversely contacts the anterior margin of the parietal in an interdigitate suture and its posterolateral corner overlies the anterodorsal end of the laterosphenoid in lateral view.
Parietal The partial left parietal and nearly complete right parietal are preserved in dorsal view (Fig. 1A, B). The parietals are sutured along the dorsal midline, unlike the fusion of the two bones in other therizinosaurians such as Erlikosaurus (Clark et al., 1994; Lautenschlager et al., 2014). Clark et al. (1994) had considered a tiny crack near the midline of the two parietals to be a suture, but CT data rejected this interpretation (Lautenschlager et al., 2014). The parietal fusion has been reported in most theropods, including early-branching theropods (Coria and Currie, 2003; Brusatte and Sereno, 2007), tyrannosauroids (Brochu, 2003; Xu et al., 2004), oviraptorosaurians (Makovicky and Sues, 1998; Clark et al., 2002; Balanoff et al., 2009), and troodontids (Makovicky and Norell, 2004; Xu and Norell, 2004). The lack of parietal fusion is likely to be an indicator that IVPP V 11559 is still in an early ontogenetic stage, though fusions are seen in various vertebrae of the holotype.
The parietal is a large flat bone and slightly arched transversely (Fig. 1A, B). As in Erlikosaurus (Clark et al., 1994), the main body of parietal is subrectangular in outline, but it seems to be comparatively wider transversely than in Erlikosaurus (Fig. 1B). The minimum transverse width of the parietal is more than twice that of the frontal, while in Erlikosaurus the two bones are about the same in transvers width. The proportionally wider parietal might represent an ontogeny-related feature. The dorsal surface is flat, continuous with that of the frontal, unlike in many other maniraptorans in which the surface is flexing along the frontal-parietal contact and faces posterodorsally (Clark et al., 1994). The posterior skull roof lacks a sagittal crest between the two parietals but contains a wide interfenestral area (Fig. 1A, B) as in Erlikosaurus (Clark et al., 1994), many early-branching theropods (Rauhut, 2003), ornithomimosaurs (Makovicky et al., 2004), and some alvarezsaurians such as Shuvuuia (Choiniere et al., 2014). The supratemporal fossa is delimited medially by a distinctive longitudinal ridge that locates about the mid-width of the parietal and anteriorly by a curved ridge that is about 5 mm away from the frontal-parietal suture. This ridge is obscured laterally. A supratemporal fossa that is located entirely within the parietal seems to be a therizinosaurian feature. Another possible therizinosaurian feature is the sub-trapezoidal outline of the supratemporal fossa, with the medial border being much shorter than the lateral border. The anteromedial corner of the main body of the parietal bears a long and sharp process. This process together with its counterpart from the other parietal, projects anteriorly and deeply inserts into the two frontals along the dorsal midline (Fig. 1A, B). In Erlikosaurus, the anteromedial process is the minimum size. Anterolaterally, the postorbital process contacts the posterior margin of the postorbital process of the frontal and its lateral margin overlies the laterosphenoid (Fig. 1B). It is triangular and relatively robust. The squamosal process is large and as in Erlikosaurus (Lautenschlager et al., 2014), there is a posterolaterally oriented lambdoidal crest on the squamosal process dorsally. Distally the squamosal process is bifurcated, and the ventral process is sharp and much longer than the dorsal process (i.e, the lambdoidal crest), unlike the one in Erlikosaurus which is subequal in length to the latter process (Lautenschlager et al., 2014). As in Erlikosaurus (Lautenschlager et al., 2014), the occipital portion of the parietal faces dorsally rather than posteriorly, but it differs from Erlikosaurus in having a much more pronounced supraoccipital process.
Postorbital The right postorbital is preserved in V 11559. As typical of theropods, the postorbital is a tri-radiate bone with an anterior frontal process, a posterior squamosal process, and a ventral jugal process (Fig. 1E). The anterior frontal process is the longest and most robust one of the three processes, and is similar in general morphology to that of Erlikosaurus (Lautenschlager et al., 2014). The frontal process curves anterodorsally from the main body, as in other maniraptorans (Currie, 1985; Norell et al., 2006; Norell et al., 2009; Lautenschlager et al., 2012) including Erlikosaurus (Lautenschlager et al., 2014). In addition, the frontal process is uniquely comparable to that of Erlikosaurus (Lautenschlager et al., 2014) in several features including that the process is robust and with little anterior extension, has an accessory process extending medially, and a rugose area formed by transverse ridges is located at the base of the accessory medial process. However, V 11559 differs from Erlikosaurus in that the main body of the frontal process is proportionally shorter, its accessory medial process is proportionally longer, and the rugose area is more dorsally located. There is a shallow depression on the posterolateral surface of the accessory process.
The squamosal process is transversely thin and has a subtriangular outline in lateral view, with a slightly concave lateral surface. There is a small convexity along the dorsal margin of the squamosal process near its base. As in Erlikosaurus, the squamosal process diminishes sharply in width posteriorly and sends out a tapered process posteriorly.
The jugal process is slender and bears a broad anterior surface within the orbit, as in other therizinosaurians (Clark et al., 1994; Lautenschlager et al., 2014; Zanno, 2010b). In general, the jugal process is more similar to that of Erlikosaurus than of Falcarius: the process is relatively short; it is relatively straight in lateral view; and it displays apparent anteroposterior narrowing distally, in contrast to the jugal process of Falcarius which is proportionally long, distinctively curved in lateral view, and is nearly consistent in width for most of its length.
Quadrate The quadrate is represented by a partial left one missing its dorsal portion and a small portion close to the ventral end, exposing only the medial half of the posterior surface (Fig. 2B). As in Erlikosaurus (Lautenschlager et al., 2014), the main body can be divided into a posterodorsally arched dorsal portion and a more flattened and straighter ventral portion. Dorsally there appears to be a large process along the lateral margin of the quadrate shaft as in Erlikosaurus, representing the squamosal process (which also articulates the exoccipital). The ventral end of the quadrate is mediolaterally wide and blunt in posterior view as in Falcarius (Zanno, 2010b), and clearly bears a central condyle on the posterior surface, suggesting the presence of the tricondylar condition as in Erlikosaurus, probably Jianchangosaurus (Pu et al., 2013; Lautenschlager et al., 2014), and the ornithomimosaurian Sinornithomimus (Kobayashi and Lü, 2003), but a central condyle is absent in most other theropods including Falcarius (Zanno, 2010b).
Vomer A long slender bone is tentatively identified as the vomer, missing both the anterior and posterior ends (Fig. 1D). It is probably a compound bone formed by a fusion of the left and right vomer, as in Erlikosaurus (Clark et al., 1994; Lautenschlager et al., 2014), which is the only other therizinosaurian with this element known. In Erlikosaurus, though the vomer is a single element, CT scans show that the vomer consists of two co-ossified counterparts indistinguishable externally, a typical feature of theropods (Lautenschlager et al., 2014). The preserved anterior end is dorsoventrally flattened, with the transverse width considerably greater than the dorsoventral depth. Posteriorly the bone becomes thinner transversely than dorsoventrally, with a sub-triangular cross section. The preserved length of the vomer suggests that it is long anteroposteriorly and probably extends posteriorly to contact the cultriform process as in Erilikosaurus (Clark et al., 1994; Lautenschlager et al., 2014).
2.2 Braincase
Laterosphenoid The right laterosphenoid is partially preserved and exposes its lateral surface (Fig. 1A). The laterosphenoid contacts the postorbital process of the frontal anteriorly and is overlain by the frontal process of the postorbital anterodorsally, and by the parietal dorsally. It is composed of a main body which contributes to the anterolateral wall of the braincase and a laterally projected horizontal process. The main body is slightly arching along the medial margin of the supratemporal fenestra. The lateral process of the laterosphenoid contacts the posterior margin of the postorbital process of the parietal, and both elements contact the ventral surface of the frontal process of the postorbital as in Erlikosaurus (Clark et al., 1994; Lautenschlager et al., 2014).Prootic The prootic is represented by the left one, exposing the medial surface (Fig. 2C). It is irregular in shape, contacting the exoccipital/opisthotic posteriorly, the laterosphenoid and orbitosphenoid anteriorly, the parietal dorsally, and the basisphenoid ventrally. In Erlikosaurus, the medial surface of the prootic is swollen ventrally, accommodating the vestibular eminence (Lautenschlager et al., 2014). The vestibular eminence of Beipiaosaurus inexpectus is mostly broken, but displays a slight swelling ventrally. Dorsal to the vestibular eminence, there is a dorsally pointed process, which contributes to the anterior portion of the floccular fossa as in Erlikosaurus (Lautenschlager et al., 2014). Internally, the prootic contains the anterior and lateral semicircular canals, as well as the cochlear duct. Anterior to the vestibular eminence, there are two foramina for vestibulocochlear (VIII) and facial (VII) nerves. The vestibulocochlear foramen and facial foramen are similar in size and both exit laterally, unlike Erlikosaurus in which the vestibulocochlear foramen is smaller and penetrates posteriorly and the facial foramen is enlarged (Lautenschlager et al., 2012, 2014). Anteriorly, there is a large notch representing the foramen for the maxillary branch of trigeminal nerve (V2).
2.3 Mandible
Dentary The dentary is represented by a nearly complete right dentary missing its anterior end and posterodorsal end, and it exposes its medial side (Fig. 1D). The preserved portion is about 65% the length of the femur, suggesting that the skull is relatively large (Xu et al., 1999) as in other early-branching therizinosaurians (e.g., this ratio is 62% in Falcarius (Zanno, 2010b) and 55% in Jianchangosaurus (Pu et al., 2013)). In late-branching therizinosaurians, the skull is relatively small (e.g., this ratio is about 35% in Alxasaurus (Russell and Dong, 1993) and Segnosaurus (Perle, 1979)).The dentary is a long and thin bone, sub-triangular in lateral view. In medial view, dentary symphysial region seems to be downturned, though the region is mostly broken. The anterior one-third of the dentary exhibits sub-parallel dorsal and ventral margins in lateral view and the posterior two-thirds of the dentary significantly deepens posteriorly. In most other therizinosaurians (Zanno et al., 2016), possibly except for Neimongosaurus (Zhang et al., 2001), the sub-parallel portion is close to the half of the dentary length or even longer. The minimum dorsoventral height of the dentary is approximately 5 mm, which is about one-fourth of the maximum height (approximately 20 mm). This ratio is much greater in most other therizinosaurians, including Jianchangosaurus (Pu et al., 2013), Alxasaurus (Dong, 2002), Neimongosaurus (Zhang et al., 2001), Erlikosaurus (Clark et al., 1994; Lautenschlager et al., 2014), Segnosaurus (Perle, 1979), and Falcarius (Kirkland et al., 2005; Zanno, 2010b). The ventral margin of the dentary descends strongly posteriorly, which is a feature seen in most therizinosaurians including Falcarius (Zanno, 2010b; Zanno et al., 2016). The dorsal margin of the dentary is transversely thickened, particularly along the posterior two-thirds, but otherwise the dentary is extremely thin transversely.
The dentary tooth row is located relatively close to the lateral surface of the dentary along the anterior one-third of the dentary, but more posteriorly it is inset from the lateral margin, forming a lateral shelf as in most other therizinosaurians except Falcarius (Zanno, 2010b). The Mecklian groove is anteriorly deep and narrow, and is located close to the ventral margin of the dentary as in Segnosaurus (Zanno et al., 2016), unlike the more dorsally located Mecklian groove in Erlikosaurus (Lautenschlager et al., 2014). More posteriorly, the Mecklian groove becomes shallower and wider, transiting into a wedge-shaped fossa, as in Falcarius (Zanno, 2010b) and Erlikosaurus (Lautenschlager et al., 2014).
Surangular Left surangular is partially preserved, missing the anterior portion, and exposes its lateral surface (Fig. 1C). The surangular forms the majority of the lateral surface of the posterior half of the mandible. It contacts the angular ventrally, and together with the latter, it forms a large external mandibular fenestra, which is elliptic in outline, and much deeper dorsoventrally than that of other therizinosaurians such as Segnosaurus (Zanno et al., 2016), Erlikosaurus, and Jianchangosaurus (Pu et al., 2013). Two surangular foramina are present on the lateral surface, the anterior one of which is smaller and positioned dorsal to the external mandibular fenestra, and the posterior one of which is larger in size and is located posterodorsal to the external mandibular fenestra.
Angular Posterior half of the left angular is preserved and exposes its lateral surface (Fig. 1C). It is a transversely thin element, and forms the posteroventral and ventral borders of the external mandibular fenestra.
Fig. 1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1Close-up photographs of cranial elements of Beipiaosaurus inexpectus (IVPP V 11559)
A. right frontal, left parietal, and right laterosphenoid in dorsal view; B. right parietal in dorsal view; C. left surangular and left angular in lateral view; D. vomer and right dentary in medial view; E. right postorbital in lateral view Scale bars for A-C and E equal 1 cm; for D equal 2 cm
2.4 Dentition
There are 37 alveoli preserved in the right dentary, which is missing its anterior portion, suggesting that Beipiaosaurus inexpectus has more than 37 dentary teeth. This number is similar to that in Alxasaurus (Russell and Dong, 1993) and Eshanosaurus (Xu et al., 2001), and is much greater than in most other therizinosaurians (e.g., 25-28 in Jianchangosaurus (Pu et al., 2013) and Falcarius (Zanno, 2010b), 31 in Erlikosaurus (Lautenschlager et al., 2014), and 24 in Segnosaurus (Zanno et al., 2016)). The dentary teeth are restricted in their distribution to the anterior two-thirds of the dentary (Fig. 1D), as in Segnosaurus and Jianchangosaurus, but unlike the more posteriorly extended tooth row in Erlikosaurus (Zanno et al., 2016). In medial view, there are dorsally pointed, sub-triangular interdental plates between neighboring teeth (Xu et al., 1999), as in Alxasaurus (Russell and Dong, 1993), Segnosaurus (Perle, 1979), and Falcarius (Zanno, 2010b; Zanno et al., 2016).Fig. 2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Close-up photographs of cranial elements of Beipiaosaurus inexpectus (IVPP V 11559)
A. partial nasal; B. right quadrate in posterolateral view; C. left prootic in medial view. Scale bars equal 1 cm
The preserved dentary teeth are relatively homodont, and they are folidont (Hendrickx et al., 2015), bearing a pronounced constriction cervix and displaying a lanceolate outline in lateral view (Fig. 3B, C). It is unknown whether the anteriormost dentary teeth are enlarged and conidont as in most other therzinosaurians (Russell and Dong, 1993; Zanno, 2010b; Pu et al., 2013; Zanno et al., 2016). There is no wear facet on the dentary teeth, as in Falcarius (Zanno, 2010b), Erlikosaurus (Clark et al., 2004), and Alxasaurus (Russell and Dong, 1993), but Segnosaurus has been reported to display wear facet on the dentary teeth.
The tooth crowns are short, posteriorly recurved, and basally bulbous (Xu et al., 1999) (Fig. 3B, C, E), as in Falcarius (Zanno, 2010b) and Jianchangosaurus (Pu et al., 2013), in contrast to the more symmetrical ones in Erlikosaurus as well as the relatively tall, labiolingually compressed crowns in Segnosaurus (Zanno et al., 2016). Coarse denticles are present in both mesial and distal carinae (Fig. 3E) as in most other therizinosaurians (Clark et al., 1994; Xu et al., 1999) except for Falcarius in which denticles are proportionally small as in most other theropods. The denticles are similar in size (three denticles per millimeter along the crown carina) to those in Jianchangosaurus (Pu et al., 2013), larger than in Erlikosaurus (Clark et al., 1994), but smaller than in Segnosaurus (Zanno et al., 2016). As in Alxasaurus (Russell and Dong, 1993), the denticles are pointed toward the crown tip (Fig. 3B, C, E), and apically oriented denticles are also present in other therizinosaurians such as Erlikosaurus (Clark et al., 1994) and Segnosaurus (Zanno et al., 2016), although to a less degree.
All the preserved teeth are constricted at the crown base, and their roots are slightly compressed mediolaterally (Fig. 3D), unlike the ones with a more sub-circular cross section in Erlikosaurus (Clark et al., 1994) and Segnosaurus (Zanno et al., 2016). Large oval resorption pits, which contain replacement teeth, are present on the medial surfaces of some of the roots of functional teeth (Fig. 3A) as in Erlikosaurus (Clark et al., 1994).
Fig. 3
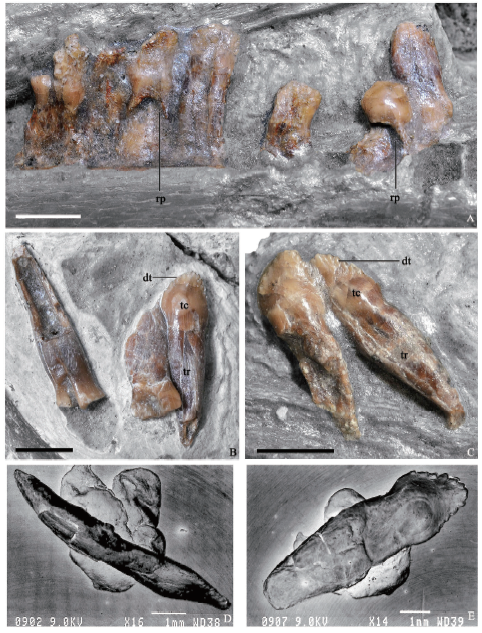 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3Close-up photographs of dentition of Beipiaosaurus inexpectus (IVPP V 11559)
A. in situ dentition in medial view; B, C. isolated teeth in lateral view; D. an isolated tooth in distal view; E. an isolated tooth in lateral view. Scale bars for A-C equal 3 mm; for D-E equal 1 mm
3 Discussion
The holotype of Beipiaosaurus inexpectus preserves only a few cranial elements, but they display many morphological features informative for the systematic position of this taxon. Here we discuss some cranial features that probably represent the autapomorphies for B. inexpectus as well as some others that may help to verify the existing phylogenetic relationships of the taxon among maniraptoran theropods. They are discussed below.The accessory process of the frontal process of the postorbital In both Beipiaosaurus inexpectus and Erlikosaurus (Lautenschlager et al., 2014), the frontal process of the postorbital is bifurcated, with an accessory process extending medially to form the anterior border of the supratemporal fossa. It is not known whether this accessory process is present in Falcarius (Zanno, 2010b), and a large depression near the posterior margin of the frontal in Falcarius might represent the contact facet for this accessory process. Nevertheless, this process is absent in non-therizinosaurian theropods (such as Haplocheirus (IVPP V 14988), Dilong (V 14243), Incisivosaurus (V 13326)) and thus it might be either a synapomorphy for the Therizinosauria or a slightly less inclusive clade.
The squamosal process of the postorbital sharply tapering off distally In Beipiaosaurus inexpectus, the squamosal process decreases its width abruptly, which is also seen in Erlikosaurus (Clark et al., 1994). This feature might be a synapomorphy for the Therizinosauroidea or even the more inclusive Therizinosauria.
Rugose area on the accessory process of the frontal process of the postorbital in lateral view There is a rugose area formed by transverse ridges on the lateral surface of the accessory process of the frontal process close to the main body of frontal process in Beipiaosaurus inexpectus. This feature is also present in Erlikosaurus but absent in most other theropods (such as Haplocheirus (V 14988), Dilong (V 14243), Incisivosaurus (V 13326)), suggesting that it might be a synapomorphy for the Therizinosauroidea or even a more inclusive clade.
Tricondylar condition of the quadrate ventral end A central condyle is present on the posterior surface of the ventral end of the quadrate in Beipiaosaurus inexpectus. A similar feature is also present in Erlikosaurus, but in most other theropods (Lautenschlager et al., 2014) the ventral end of quadrate bears only two condyles: i.e., a medial and a lateral one. A tricondylar condition of the quadrate ventral end might be a synapomorphy for the Therizinosauroidea or even a more inclusive clade.
The occipital region of the parietal facing dorsally In Beipiaosaurus inexpectus, the occipital surface of the parietal faces dorsally rather than posteriorly as in most other theropods. A dorsally facing occipital surface of the parietal is also seen in Erlikosaurus, and it is possibly a synapomorphy for the Therizinosauroidea or even a more inclusive therizinosaurian clade.
The external mandibular fenestra large and posteriorly located In Beipiaosaurus inexpectus, the external mandibular fenestra is large and extremely posteriorly located, the posterior border of which is only about 10 mm away from the glenoid. Most of other therizinosaurians also have a large and posteriorly located external mandibular fenestra, though not as posteriorly located as in Beipiaosaurus. This feature is probably a synapomorphy for the Therizinosauroidea or Therizinosauria, depending on the condition in Falcarius. In most other theropods, such as Haplocheirus (V 14988), Citipati, Oviraptor (Clark et al., 2002), the external mandibular fenestra is not as posteriorly located as in therizinosaurians.
The large posteroventral process of the dentary In Beipiaosaurus inexpectus, the dentary bears a large posteroventral process, which is about 40% of the dentary length. A similarly-sized process is also present in many other therizinosaurians, but it appears to be absent in Falcarius (Zanno, 2010b). In most theropods, except oviraptorosaurians, the posteroventral process is small (such as Haplocheirus (V 14988), Dilong (V 14243)). A large dentary posteroventral process might represent a therizinosauroid synapomorphy, depending on the relationship between the Therizinosauria and Oviraptorosauria.
Teeth with large denticles In Beipiaosaurus inexpectus, the denticles are large in size in all the preserved teeth, and this feature is also present in most other therizinosaurians including Jianchangosaurus (Pu et al., 2013), Erlikosaurus (Clark et al., 1994), and Segnosaurus (Zanno et al., 2016). However, in Eshanosaurus and Falcarius the denticles are small in size, suggesting that this feature is possibly a synapomorphy for a clade that is slightly more inclusive than the Therizinosauroidea. Large denticles are also seen in the teeth of some troodontids (Farlow, 1991) as well as some sauropodomorphs such as Lufengosaurus, but it is probably independently evolved.
Downturned anterior end of the dentary symphysis Although the symphyseal region of the dentary is missing in the holotype specimen of Beipiaosaurus inexpectus, the preserved anterior portion of the dentary indicates that the mandibular symphyseal region is downturned as in most therizinosaurians, except Falcarius (Kirkland et al., 2005). This feature is possibly a synapomorphy for a clade that is slightly more inclusive than the Therizinosauroidea. This feature is also present in some ornithomimosaurians and late-branching oviraptorsaurians, but it might be possible to have evolved independently in these three clades.
The lateral shelf of the dentary In Beipiaosaurus inexpectus, there is a shelf lateral to the dentary tooth row as in most other therizinosaurians, except Falcarius. This feature is possibly a synapomorphy for a clade that is slightly more inclusive than the Therizinosauroidea. A dentary lateral shelf is also known in most Ornithischia (Paul, 1984) and it is probably independently evolved.
Vomer extremely long The vomer is not completely preserved, but the preserved portion of the vomer suggests that it is an extremely long element as in several therizinosaurians such as Erlikosaurus and Nothronychus (Kirkland and Wolfe, 2001). This feature is likely to be a synapomorphy for the Therizinosauroidea or even a more inclusive clade.
Beipiaosaurus inexpectus has several cranial features which are different from those of other therizinosaurians in which the cranium region is known. These features are considered to be diagnostic features for B. inexpectus, until they are later demonstrated to be present in other therizinosaurians.
The postorbital process of the frontal large and its abrupt transition from the orbital rim In Beipiaosaurus inexpectus, the postorbital process of the frontal is large and transits from the orbital rim abruptly, unlike the small and smoothly transited one in most other therizinosaurians.
A long and sharp anterior process of the parietal In Beipiaosaurus inexpectus, the anteromedial corner of the parietal sends out a long and sharp anterior process (Fig. 1A, B). A similar process is absent in Erlikosaurus but unknown in other therizinosaurians owing to poor preservation.
The elongate ventral ramus of the squamosal process of parietal In Beipiaosaurus inexpectus, the distally bifurcated squamosal process of the parietal has a ventral ramus much longer than the dorsal one, while in most theropods including Erlikosaurus, they are subequal in length.
External mandibular fenestra deep dorsoventrally and extremely posteriorly located In Beipiaosaurus inexpectus, the external mandibular fenestra is not only long anteroposteriorly as in other therizinosaurians, but also deep dorsoventrally, unlike the shallow condition seen in other known therizinosaurians. It is also more posteriorly located than in other therizinosaurians.
As discussed above, the cranial features of Beipiaosaurus inexpectus provide strong support for the therizinosaurian affinity of the taxon, and they further demonstrate that B. inexpectus has already acquired many of the cranial modifications seen in late-branching therizinosaurians. New diagnostic features from this cranial study further support the validity of the taxon and some of the previously unknown cranial features derived from this study indicate that early therizinosaurians differ in cranial morphology, which is possibly related to adaptations to different ecological niches.
Acknowledgments
We thank Ding Xiaoqing for preparing the specimen as well as You Hailu and Wu Xiaochun for their constructive comments. This work is supported by the National Natural Science Foundation of China (Grant No. 41688103 and 91514302) and the Strategic Priority Research Program of the Chinese Academy of Sciences (Grant No. XDB18030504).参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 21]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 2]
[本文引用: 27]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 4]
[本文引用: 14]
[本文引用: 1]
[本文引用: 7]
[本文引用: 1]
[本文引用: 1]
[本文引用: 7]
[本文引用: 2]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 21]
[本文引用: 13]
[本文引用: 2]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}