0 引言
【研究意义】日周期是玉米生育周期中的基本代谢单元。光合碳代谢是玉米进行干物质积累的根本途径,探究不同杂交种在干旱胁迫下单位时间的代谢规律和响应差异,对于加深对品种特性的认识及筛选光合育种指标都具有重要的理论和实践意义。【前人研究进展】光合作用是对环境最敏感的生理过程之一,在规律性的光温变化环境下的光合碳固定、分配决定了植株的代谢特征和环境适应能力。光合速率是植物光合作用研究中应用最广泛的参数,也是直接体现作物光合能力水平的重要指标,大量的研究将强光高温下的最高光合速率作为评判玉米杂交种光合生产能力强弱的依据,但叶源器官的光合作用是阶段性持续进行的,因此全天的光合速率总体情况以及逆境下的表现决定了最终的光合碳积累水平。国内外****以玉米[1-4]、高粱[5]、水稻[2]等作物为对象,对光合日变化进行了广泛深入的研究,发现C3植物多呈现双峰曲线,高温强光下往往出现“光合午休”,而以玉米为代表的C4植物则往往不会有这种现象的出现[1, 6-7],晴天玉米叶片光合日变化趋势与光强的变化基本一致,12:00—14:00光强最高时光合速率达到一天中的最高值[8-9],干旱胁迫会造成其相应参数的降低[10-11]。光合碳在玉米叶片中的固定形式主要是蔗糖和淀粉,二者浓度会随着光合作用的进行而逐步提高,且二者的合成存在一定竞争关系,环境胁迫导致光合碳积累总量降低和淀粉浓度下降的同时,蔗糖含量稳定或有一定程度提高[12-14];非光照条件下叶源器官的淀粉会规律性地降解用以维持蔗糖的输出,这已在不同作物种类和品种间的比较研究中得到证实,并且双糖的输出量和输出速率存在差别 [15-16]。因此,光合碳总量的变幅和再分配特性等可能可以用于不同品种的类型划分。【本研究切入点】本研究是基于这一现状,针对干旱条件下的玉米光合碳固定、分配的日变化及其品种偏向性特征,通过设置不同程度干旱逆境,在玉米关键生育时期——吐丝期进行胁迫处理,从单位时间内玉米穗位叶生理生化动态变化的角度进行研究,对其品种差异进行分析。【拟解决的关键问题】进一步明确玉米叶源器官的光合速率、光合初始产物浓度的变化特征,以及对梯度干旱胁迫的响应差异,为挖掘玉米品种差异性评价的新指标提供一定的参考基础和理论依据。1 材料与方法
1.1 试验设计
试验于2012、2013年在中国农业大学吴桥实验站进行。选择玉米杂交种郑单958、先玉335、浚单20、农大108和ND66为试验材料(种子来源为纯系父母本,经人工授粉获得)。采用人工旱棚与控水池结合的方法控制土壤水分含量,从土壤饱和含水量至凋萎含水量范围内设置4级水分梯度,即土壤相对含水量分别达到85%、65%—70%、55%—60%、45%—50%,分别代表土壤水分供应充足对照(control)、轻度干旱胁迫(mild stress)、中度干旱胁迫(moderate stress)、重度干旱胁迫(severe stress)。5月1日播种,人工点播,种植密度为67 500株/hm2。水分处理时期为玉米吐丝期,通过提前控水使植株生长至吐丝期时达到相应土壤水分要求。分别在0:00(2012年未设置)、4:00、10:00、15:00、20:00五个时间点对穗位叶进行取样,同日使用LI-COR 6400便携式光合测定系统对穗位叶光合速率等参数及相应的生态环境要素进行测定。光合速率测定时,叶室CO2浓度、光强、温度模拟实时环境条件进行设定,于6:00—18:00进行测定,时间间隔为3 h,每个处理重复4次。采用高效液相色谱法测定蔗糖含量,采用蒽酮硫酸法测定淀粉含量。
1.2 计算方法
光合生产的最初阶段蔗糖、淀粉进行大量快速积累,光合碳转化相对较少,此时浓度的提高是真正意义上的含量升高。因此,针对光合初始期,参照张荣铣的方法[17],计算出6:00—9:00的光合速率积累量;利用10:00与4:00的蔗糖、淀粉浓度差值,以及二者的差值比值,对应相应的光合速率积累量,分别计算单位光合速率对蔗糖浓度提高的贡献率,单位光合速率对淀粉浓度提高的贡献率和单位光合速率对蔗糖/淀粉比的贡献率,具体计算公式如下:单位光合速率对蔗糖浓度提高贡献率=

单位光合速率对淀粉浓度提高贡献率=

单位光合速率对蔗糖/淀粉比贡献率=

1.3 数据统计
利用Microsoft Excel和SAS 9.1软件进行试验数据标准化处理和统计分析,采用GraphPad Prism进行作图。2 结果
2.1 梯度干旱胁迫下玉米穗位叶光合速率日变化特征
从一天中不同时间点的定时测定结果看(图1),充足水分供应情况下光合速率呈现单峰曲线变化趋势,一天当中的光合高峰出现在12时前后。充足的水分供应使得高温强光对叶片的伤害在一定程度上降至最低,品种间没有出现光合速率较低的品种,均表现高产玉米杂交种所具有的充分利用适宜光温快速生产的高光合特征;轻度干旱胁迫导致全天光合水平略有下降,峰值出现了18.5%—37.5%的降低,且在12:00之后快速下降,降幅均达到60%以上;中度干旱胁迫初期表现与前两级水分处理相近,9:00过后光照和温度快速爬升,还未达到适宜于玉米生长的光温条件,其光合速率便出现下降趋势,12:00左右光合速率均处于14 μmol·m-2·s-1以下;重度干旱胁迫对穗位叶的光合造成了极大的抑制,光合速率在9:00达到峰值20 μmol·m-2·s-1,之后随光照的增强和温度的上升而快速下降,维持在5 μmol·m-2·s-1上下直至光照完全消失。从干旱胁迫处理的品种间比较看,农大108的综合表现相对较好,降幅偏小。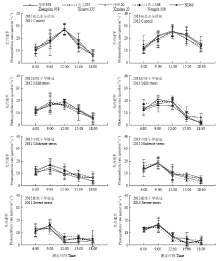 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1梯度干旱胁迫下吐丝期不同玉米杂交种穗位叶光合速率的日变化
-->Fig. 1Diurnal variation of photosynthetic rate of ear leaf of different maize hybrids under gradient drought stresses at silking stage
-->
2.2 梯度干旱胁迫下玉米穗位叶蔗糖浓度日变化特征
作物光合生产的实质是碳水化合物的积累,即绿色组织光合同化后的初级产物经多种形式转化转运的最终结果。对玉米而言,蔗糖和淀粉是最主要的光合碳固定形式,二者的浓度在很大程度上直接反映了光合碳的积累和分配情况。由图2可知,蔗糖的日周期代谢特征呈单峰曲线变化,日出前为全天最低值,尔后随环境光温资源的补给,即光合作用的启动以及光合速率的升高逐渐升高,在12:00—15:00达到峰值30—40 mg·g-1,然后平稳下降;土壤干旱胁迫并未对蔗糖的全天浓度极值造成很大影响,但从不同品种的比较看,品种间存在一定的差异。综合来看,农大108和ND66的穗位叶蔗糖浓度略低于其余3个杂交种。
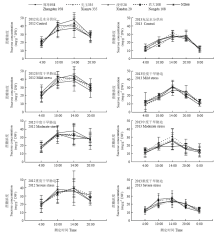 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2梯度干旱胁迫下吐丝期不同玉米杂交种穗位叶蔗糖浓度的日变化
-->Fig. 2Diurnal variation of sucrose concentration in ear leaf of different maize hybrids under gradient drought stresses at silking stage
-->
2.3 梯度干旱胁迫下玉米穗位叶淀粉浓度日变化特征
与蔗糖浓度的变化特征相似,玉米穗位叶淀粉浓度也呈现单峰变化曲线(图3),其浓度高峰出现时间在15:00前后,且峰值高于蔗糖。2012年的测定结果表明,穗位叶淀粉浓度在中度和重度干旱胁迫下的变幅都在30 mg·g-1之内,2013年的测定结果的降幅则相对较小,峰值浓度降幅在17.7%—35.6%。品种间比较发现,先玉335的淀粉浓度在多个处理、多个时间点都相对偏低;整体来看,虽然各个杂交种间正常水分供给下峰值没有太大差异,但从光合初始阶段(10时前)的淀粉浓度看,农大108和ND66两个杂交种在光合初始期就迅速积累淀粉,说明在良好的环境条件下二者在淀粉积累上有较大的优势。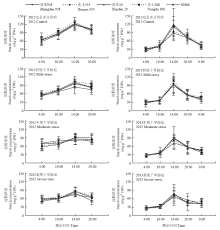 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3梯度干旱胁迫下吐丝期不同玉米杂交种穗位叶淀粉浓度的日变化
-->Fig. 3Diurnal variation of starch concentration in ear leaf of different maize hybrids under gradient drought stresses at silking stage
-->
2.4 干旱胁迫下光合碳流向偏向性的品种差异
为了分析土壤干旱胁迫对玉米光合碳流向造成的影响及其品种间的差异,计算得到了单位光合速率对蔗糖浓度提高的贡献率,单位光合速率对淀粉浓度提高的贡献率和单位光合速率对蔗糖/淀粉比的贡献率(表1)。Table 1
表1
表1玉米杂交种吐丝期单位光合速率对叶片光合碳固定相关指标的贡献率
Table 1Contribution to photosynthetic carbon from unit photosynthetic rate of different maize hybrids at silking stage
| 年份 Year | 杂交种 Hybrid | 充足水分供应 Control | 轻度干旱胁迫 Mild stress | 中度干旱胁迫 Moderate stress | 重度干旱胁迫 Severe stress | 平均 Mean | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 单位光合速率对蔗糖浓度提高贡献率 CISuC | 单位光合速率对淀粉浓度提高贡献率 CIStC | 单位光合速率对蔗糖/淀粉比贡献率 CRSu/St | 单位光合速率对蔗糖浓度提高贡献率 CISuC | 单位光合速率对淀粉浓度提高贡献率 CIStC | 单位光合速率对蔗糖/淀粉比贡献率 CRSu/St | 单位光合速率对蔗糖浓度提高贡献率 CISuC | 单位光合速率对淀粉浓度提高贡献率 CIStC | 单位光合速率对蔗糖/淀粉比贡献率 CRSu/St | 单位光合速率对蔗糖浓度提高贡献率 CISuC | 单位光合速率对淀粉浓度提高贡献率 CIStC | 单位光合速率对蔗糖/淀粉比贡献率 CRSu/St | 单位光合速率对蔗糖浓度提高贡献率 CISuC | 单位光合速率对淀粉浓度提高贡献率 CIStC | 单位光合速率对蔗糖/淀粉比贡献率 CRSu/St | ||||||
| 2012 | 郑单958 Zhengdan 958 | 0.534 | 0.523 | 0.027 | 0.465 | 0.472 | 0.018 | 0.394 | 0.193 | 0.035 | 0.327 | 0.111 | 0.024 | 0.430 | 0.325 | 0.026 | ||||
| 先玉335 Xianyu 335 | 0.405 | 0.462 | 0.026 | 0.457 | 0.291 | 0.038 | 0.408 | 0.193 | 0.024 | 0.338 | 0.075 | 0.027 | 0.402 | 0.255 | 0.029 | |||||
| 浚单20 Xundan 20 | 0.283 | 0.465 | 0.018 | 0.486 | 0.285 | 0.030 | 0.459 | 0.235 | 0.033 | 0.406 | 0.160 | 0.033 | 0.409 | 0.286 | 0.029 | |||||
| 农大108 Nongda 108 | 0.354 | 0.552 | 0.037 | 0.461 | 0.405 | 0.019 | 0.328 | 0.265 | 0.024 | 0.364 | 0.190 | 0.012 | 0.377 | 0.353 | 0.023 | |||||
| ND66 | 0.527 | 0.562 | 0.023 | 0.319 | 0.351 | 0.021 | 0.438 | 0.041 | 0.019 | 0.532 | 0.095 | 0.026 | 0.454 | 0.262 | 0.022 | |||||
| 2013 | 郑单958 Zhengdan 958 | 0.105 | 0.507 | 0.004 | 0.041 | 0.391 | 0.004 | 0.018 | 0.341 | 0.012 | 0.051 | 0.214 | 0.007 | 0.054 | 0.363 | 0.007 | ||||
| 先玉335 Xianyu 335 | 0.112 | 0.417 | 0.008 | 0.086 | 0.297 | 0.007 | 0.110 | 0.386 | 0.011 | 0.053 | 0.307 | 0.009 | 0.090 | 0.352 | 0.009 | |||||
| 浚单20 Xundan 20 | 0.079 | 0.390 | 0.004 | 0.078 | 0.446 | 0.004 | 0.143 | 0.320 | 0.006 | 0.073 | 0.206 | 0.008 | 0.093 | 0.341 | 0.006 | |||||
| 农大108 Nongda 108 | 0.036 | 0.558 | 0.002 | 0.091 | 0.378 | 0.004 | 0.170 | 0.413 | 0.004 | 0.128 | 0.583 | 0.006 | 0.106 | 0.483 | 0.004 | |||||
| ND66 | 0.055 | 0.586 | 0.002 | 0.117 | 0.452 | 0.005 | 0.122 | 0.465 | 0.006 | 0.154 | 0.312 | 0.009 | 0.112 | 0.454 | 0.006 | |||||
| 平均 Mean | 郑单958 Zhengdan 958 | 0.320 | 0.515 | 0.016 | 0.253 | 0.432 | 0.011 | 0.206 | 0.267 | 0.024 | 0.189 | 0.163 | 0.016 | 0.242 | 0.344 | 0.016 | ||||
| 先玉335 Xianyu 335 | 0.259 | 0.440 | 0.017 | 0.272 | 0.294 | 0.023 | 0.259 | 0.290 | 0.018 | 0.196 | 0.191 | 0.018 | 0.246 | 0.304 | 0.019 | |||||
| 浚单20 Xundan 20 | 0.181 | 0.428 | 0.011 | 0.282 | 0.366 | 0.017 | 0.301 | 0.278 | 0.020 | 0.240 | 0.183 | 0.021 | 0.251 | 0.313 | 0.017 | |||||
| 农大108 Nongda 108 | 0.195 | 0.555 | 0.020 | 0.276 | 0.392 | 0.012 | 0.249 | 0.339 | 0.014 | 0.246 | 0.387 | 0.009 | 0.242 | 0.418 | 0.014 | |||||
| ND66 | 0.291 | 0.574 | 0.013 | 0.218 | 0.402 | 0.013 | 0.280 | 0.253 | 0.013 | 0.343 | 0.204 | 0.018 | 0.283 | 0.358 | 0.014 | |||||
新窗口打开
总体来看,光合碳流向属于较为稳定的代谢特征。单位光合速率对蔗糖浓度提高贡献率由于各水分处理间蔗糖浓度未有较大变幅而表现的相对稳定,不同品种间比较以ND66略高。单位光合速率对淀粉浓度提高贡献率参数随胁迫出现而下降,虽然这种降低并没有随胁迫的加重而逐级降低,但所造成的影响已经在光合产物的分配上得到了体现。品种间比较发现,农大108的单位光合速率对淀粉浓度提高贡献率均处于较高水平,这在两年的多个处理水平上都很稳定,2年4个水分处理的平均值为0.418,体现了偏淀粉积累的代谢特点;其次为ND66和郑单958,单位光合速率对淀粉浓度提高贡献率分别为0.358和0.344,浚单20和先玉335的值较低。单位光合速率对蔗糖/淀粉比的贡献率参数能够反映代谢偏向性,系数值高说明向蔗糖合成多,反之则说明同等的光合碳固定水平下会优先偏向于淀粉积累。在本试验条件下,先玉335为偏蔗糖代谢型,特别是在2013年的测定结果中,充足水分供应、轻度、中度和重度干旱胁迫的单位光合速率对蔗糖/淀粉贡献率分别为0.008、0.007、0.011和0.009,均为同处理品种中的最高值,农大108同时期计算值为0.002、0.004、0.004和0.006,均为同处理品种中的最低值。本试验年际间测定数值有一定的差异,这可能与两个年份玉米生长所处光温条件等差异有关,也可能与年度批次测定中的系统差异有关,但对品种间的横向比较影响有限。不同品种计算结果比较发现,先玉335的值最高,为0.019,表现为偏蔗糖代谢特点;农大108和ND66的值最低,均为0.014,表现为偏淀粉代谢特点。
3 讨论
高光合效率一直是玉米育种家新品种选育的重要目标,并基本形成了较为统一的认识,以抽雄—吐丝期的光合速率作为评价的关键时期,光照充足且相对稳定时段(10时—14时左右)的光合速率测定值能够较好地体现某一杂交种或自交系是否属于高光效品种[18-19]。本研究从光合速率日周期变化的角度,结合梯度干旱胁迫,对多个玉米杂交种的光合速率进行了分析,发现所有处理均呈现单峰变化特征,充足水分供应下光合速率在正午前后达到峰值,轻度土壤干旱使得峰值降低并提前出现;随胁迫的进一步加重,叶片光合速率全天未能超过20 μmol·m-2·s-1,且峰值出现在9时前后。相较于充足水分供应条件,光合速率在轻度干旱下有25%左右的下降;中度胁迫变幅更大,平均降幅在50%以上;重度干旱胁迫下光合速率基本处于停滞状态,降幅都在70%以上,部分处理达到90%。通过品种间比较发现,在充足水分供应条件下,特别是光合高峰时段,品种间并未出现显著差异,说明所选品种的光合生产能力较为接近,因此,在一定程度上说明,单纯依靠光合速率来进行品种划分或育种选择实现的可能性较低。作物高产不仅要求功能叶片有较强的光合生产能力,而且要求光合器官中形成的光合产物能够合理地分配运输[20]。因此,在相近的光合水平下,如何进行光合产物分配转运是光合碳代谢差异的关键。昼夜的碳流动对植株的正常生理代谢起到了重要作用,85%以上的光合产物是在形成后24小时以内运转出去的[12],夏叔芳等[15, 20]分别在大豆和玉米中证明了光合同化物在光周期中的单峰变化规律,蔗糖和淀粉的合成在一定程度上存在竞争关系,特别是在经历较长时间黑暗或胁迫结束时[16, 21-23]。本研究的结果验证了淀粉和蔗糖的代谢呈现周期性的单峰变化趋势,在光照开始前浓度最低,同等条件下蔗糖的合成启动时间点往往优先于淀粉,最适光温时间点及之后数小时维持相对较高的浓度。在本研究中,日周期内淀粉和蔗糖浓度都是呈现出较为明显的单峰变化趋势,水分供应状况会影响玉米叶片光合产物的常规代谢特性,干旱胁迫下碳同化总量下降。很多植物如豆类通过降低淀粉合成速率来维持蔗糖浓度,使可溶性碳水化合物的含量增加,获得更高的细胞渗透势,抵御环境胁迫的同时保证光合同化物的输出[24-26],本研究同样出现了干旱胁迫下蔗糖浓度峰值变化不大、淀粉含量大幅降低的情况。REDDY[12]通过14C标记的研究发现,通常水分亏缺能够降低叶片14C同化物的输出速率,也降低输导组织14C同化物的输入速率,对分配格局的改变趋势使输出比例随着水分胁迫的增强而降低。本研究结果中干旱胁迫下叶片淀粉浓度有所降低,一方面是光合固定量降低造成的,另一方面也可能与部分淀粉的分解,通过形成小分子六碳糖用于参与细胞的渗透调节,以提高抵御环境胁迫能力有关[27-32]。
不同的品种之间存在光合初产物分配比例的差别,如何将有限的光合碳进行合理的分配运输,在很大程度上决定品种的代谢类型,特别是在自然生长环境下,阶段性地遭遇土壤干旱、低温、寡照等逆境,这些逆境胁迫会使得品种代谢特性差异表现得更加突出。日出前蔗糖和淀粉浓度都处于日周期中的最低值,光合初始阶段结束时其浓度提高值可以代表该时段内的实际增长量,对该增长量与同时段内光合速率积累值的比值分析,就可以得知在光合碳分配上蔗糖和淀粉两个途径的偏向程度,说明是优先合成蔗糖还是优先合成淀粉,即单位光合碳流向蔗糖途径的比例高,还是流向淀粉途径的比例高。
因此,笔者提出了利用日变化中最初进行光合碳同化固定时间段内,单位光合速率与蔗糖及淀粉浓度的提高值及二者比例的关系,即单位光合速率对蔗糖浓度提高的贡献率,单位光合速率对淀粉浓度提高的贡献率和单位光合速率对蔗糖/淀粉比值的贡献率来进行品种类型的评估划分。单位光合速率对淀粉浓度提高贡献率参数随胁迫出现下降,虽然这种降低并没有随胁迫的加重而逐级降低,但胁迫所带来的影响已经在光合产物的分配上得到了体现,品种间比较发现农大108最高且稳定;单位光合速率对蔗糖浓度提高贡献率参数没有呈现出明显地受到干旱胁迫的影响而变化的趋势,品种间表现也相对稳定,但以ND66表现最好;单位光合速率对蔗糖/淀粉比贡献率参数是该系列参数中最能够直接反映代谢偏向型的,以先玉335为代表的蔗糖积累偏向型和农大108为代表的淀粉积累偏向型构成了两大基本类型,前者的数值稳定偏高体现在多个处理中,后者淀粉积累稳定的特点不仅明显表现在单位光合速率对淀粉浓度提高贡献率参数上,也在此参数上得到了体现。相较而言,ND66的单位光合速率对蔗糖/淀粉比贡献率参数低,单位光合速率对淀粉浓度提高贡献率参数低于农大108但处于较高的水平,具有偏淀粉代谢型特点;郑单958和浚单20在3个参数的横向比较中虽在部分处理中偶有突出表现,但规律性不明显,数值处于5个品种的中间,但其在代谢类型划分上能否归类为均衡型品种还有待进一步的研究。
4 结论
本试验中5个玉米杂交种对梯度干旱胁迫的总体响应较为一致,叶片光合速率在日周期中呈现单峰变化趋势,干旱胁迫导致峰值降低且提前出现,随胁迫程度的提高,光合速率在9:00尚未达到适宜的光温条件时就出现了大幅降低且未能恢复;玉米叶片的蔗糖、淀粉浓度日周期变化特征都为单峰曲线,前者高峰出现时间早于后者,峰值低于后者,干旱胁迫导致淀粉浓度随胁迫加重而降低,蔗糖浓度较为稳定;日周期中光合初始阶段玉米叶片蔗糖及淀粉浓度的提高值及二者的比值分别与光合速率积累量建立关系,得到单位光合速率对其的贡献率,结果表明在单位光合速率对淀粉浓度提高的贡献率方面,农大108和ND66较高且稳定,有淀粉积累优势;在单位光合速率对蔗糖/淀粉比贡献率方面,先玉335处于较高水平,有偏蔗糖积累代谢的特点。本研究认为光合速率对蔗糖/淀粉比贡献率可以在一定程度上作为玉米光合碳代谢类型划分指标。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}